3.2. Identification of Strain YHM-G
After growing for 48 h at 30 °C on WL plates, the colonies of strain YHM-G were white, raised, with dry and rough surfaces and smooth edges and not easy to pick up (
Supplementary Figure S1a). The WL medium had changed from blue to yellow where the yeast had grown (
Supplementary Figure S1a). The cells of YHM-G observed under a microscope were rodlike with asexual budding reproduction occurring at one end of the cells without an endospore and mycelium (
Supplementary Figure S1b). Thus, strain YHM-G exhibited the morphological and cellular structure of yeast.
The physiological characteristics of strain YHM-G are shown (
Supplementary Table S2). The sugar fermentations showed that strain YHM-G could use most types of sugar. Acid production and aerogenesis were observed when strain YHM-G was fermented with glucose, maltose, D-galactose and sucrose. Strain YHM-G only produced acid from D-maltose and aerogenesis from D-xylose and was unable to ferment L-rhamnose monohydrate, D-arabinose and lactose, as indicated by the lack of acid or gas production. Strain YHM-G was able to utilize glucose, maltose, D-galactose, sucrose, D-maltose, D-xylose, ethanol, glycerol, inulin, L-rhamnose monohydrate, D-arabinose and lactose but not D-raffinose, D-trehalose, mannose, D-ribose and D-sorbose. All nitrogen sources tested, including urea, ammonium sulfate, sodium nitrate, potassium nitrite, L-phenylalanine and L-lysine, were able to be used as the sole nitrogen source for growth.
The YHM-G strain was positive for the indole test, methyl red test, starch hydrolysis test, citrate test and urea test, indicating that it was able to produce indole from tryptophan, acid from glucose and amylase to decompose starch and make use of sodium citrate or urea as a carbon source or nitrogen source for growth (
Supplementary Table S2). Additionally, the Voges–Proskauer test, hydrogen sulfide test and gelatin liquification test were negative. From these physiological and biochemical characteristics, the YHM-G strain was preliminarily confirmed as
Hyphopichia sp. These results were helpful in understanding its metabolic activities, so that its metabolic pathway could play its corresponding functions.
After searching the NCBI database, the ribosomal large subunit 26S rRNA gene D1/D2 domain fragment amplificated from YHM-G showed a 100% sequence similarity with a partial 26S rRNA gene sequence from
H. burtonii (CP024760.1, MH867400.1, KY107885.1, KY107883.1, KY107882.1, KY107877.1, HF952839.2, KC192660.1 and MK373312.1). According to branch clustering, strain YHM-G was closest to
H. burtonii CBS:2352 (KY107882.1) and
H. burtonii CBS:232.37 (MH867400.1) in the phylogenetic tree constructed using MEGA 7.0.14 software (
https://www.megasoftware.net, accessed on 17 July 2022) based on the neighbor-joining statistical method (
Supplementary Figure S1c). Overall, strain YHM-G was identified as
H. burtonii, according to the analysis of morphological, physiological and biochemical characteristics, and phylogenetic analysis. Although there is relatively little research on this strain, it has been reported to exist in the production process of many fermented foods, such as
Baijiu [
28],
Huangjiu [
28], ham [
29],
Douchi [
30], Douban sauce [
31], Kombucha [
32], wine [
33], traditional sourdough starters [
34] and cocoa beans [
35] through the use of traditional screening and high-throughput sequencing methods. Its presence has most often been reported in
Baijiu, and there have been few studies on the role of this strain in other fermented foods [
28,
36]. To enhance research on the role of yeasts other than
S. cerevisiae in food, the functions of strain YHM-G will be continuously analyzed.
3.3. Tolerance Features of YHM-G
3-Met not only has outstanding flavor characteristics, which endow specific flavor characteristics to traditionally brewed food products, such as
Baijiu, but also offers the healthy function of antioxidant activity [
7,
37]. Therefore, increasing the content of 3-Met in traditionally brewed foods, such as
Baijiu, will help to improve their quality. Strain YHM-G has been identified as a yeast with a high yield of 3-Met preliminarily screened from the
Baijiu production environment. Whether it can be applied to the
Baijiu production to improve its 3-Met content depends on its biological characteristics because
Baijiu production takes place in a constantly changing microenvironment. Only the strain YHM-G was found to offer excellent environmental tolerance and survivability for a long time in the
Baijiu production process, allowing it to produce 3-Met and improve
Baijiu quality. Therefore, we analyzed the adaptability of strain YHM-G to the environment regarding its range of growth temperatures and pH and tolerance to sugar, NaCl, ethanol and 3-Met (
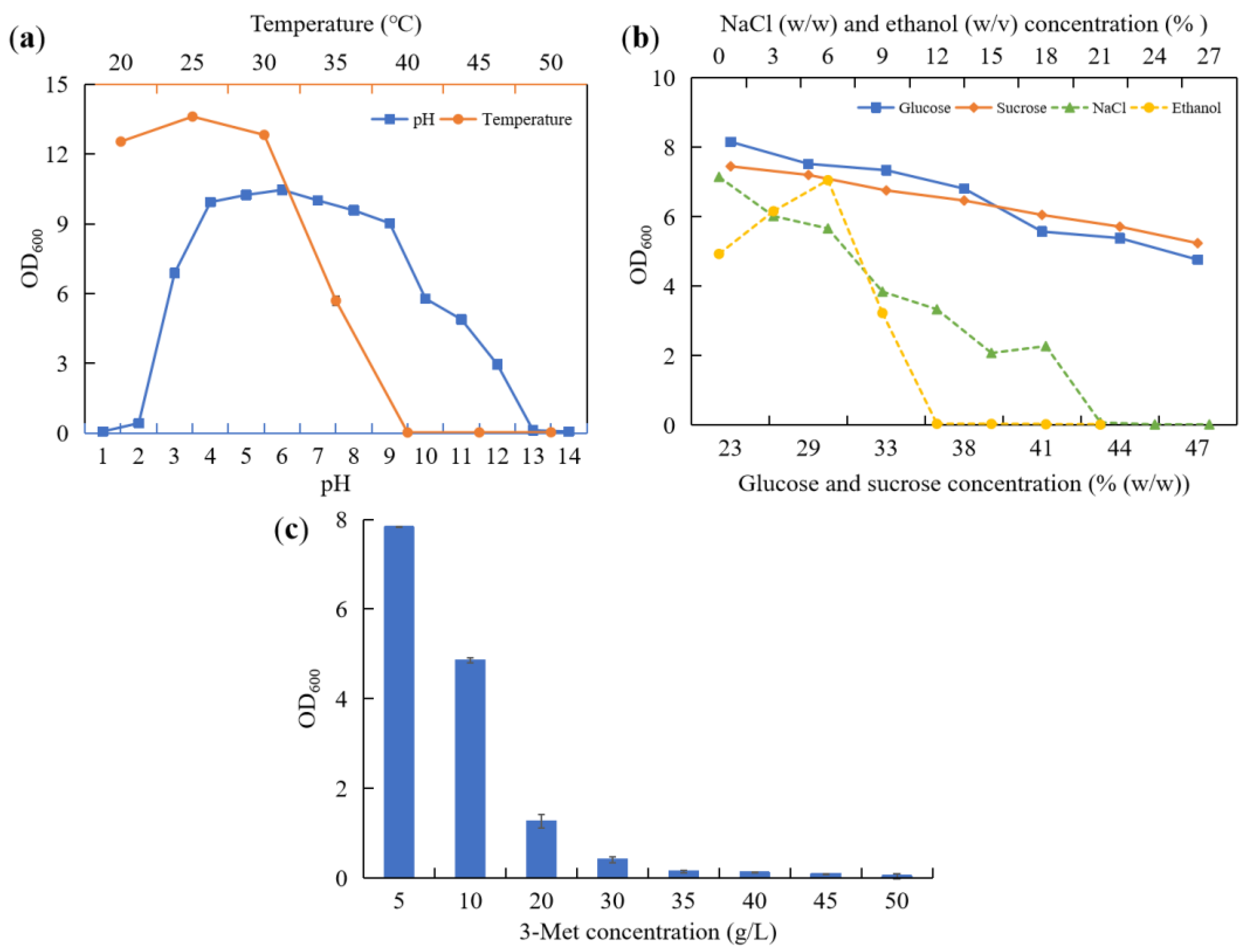
Figure 1). The optimum growth temperature of strain YHM-G was 25 °C, similar to most yeasts found in previous reports, with a maximum growth temperature of 40 °C (
Figure 1a), which was higher than most temperatures (20–40 °C) during the
Baijiu production process [
38,
39,
40,
41,
42]. Therefore, strain YHM-G can adapt well to the different temperatures used in
Baijiu production. Strain YHM-G was able to grow between pH 2 and pH 12 (
Figure 1a), a range wider than previously reported [
42]. Its optimum pH for growth was weak acidic, at pH 4–7 (
Figure 1a). This range is within the range of pH used in the
Baijiu-production process [
38,
39,
40]. As the glucose concentration, sucrose concentration and NaCl concentration increased the growth rate of strain YHM-G declined. When the concentration of glucose, sucrose and NaCl exceeded 47%, 47% and 17%, respectively, strain YHM-G could not grow (
Figure 1b). In general, yeast strains have a high tolerance to sugar with strains, such as
Pichia kudriavzevii YF1702 and
Clavispora lusitaniae YX3307 [
25,
43,
44,
45,
46]. Additionally, like other microorganisms, yeasts have a limited tolerance to NaCl, a concentration of 15% preventing the growth of most microorganisms [
42,
47]. Compared with most other reports on yeast, strain YHM-G had a high tolerance to NaCl, so it could better adapt to different environments [
47,
48]. However, there are a few yeasts with a high tolerance of up to 30% NaCl screened from particular environments, possibly related to the activation of antioxidant enzymes in the antagonistic yeast, including catalase, thioredoxin reductase, glutathione peroxidase and glutathione reductase [
49,
50]. As the ethanol concentration increased, the biomass concentration of strain YHM-G increased first and then decreased. At a concentration of ≤6%, ethanol could be used as a carbon source for the growth of strain YHM-G, consistent with the previous physiological and biochemical results (
Figure 1b and
Supplementary Table S2). At an ethanol concentration of ≥6%, the growth of strain YHM-G was inhibited because of the influence of ethanol on the cell structure and enzyme activity of the strain (
Figure 1b) [
25,
51]. The ethanol concentration was 12% at maximum growth (
Figure 1b), a value much higher than that produced during
Baijiu production processes (2–4%), thus strain YHM-G can be considered a yeast with high ethanol tolerance [
51,
52]. The ability of a strain to tolerate 3-Met determines its level of 3-Met production. The results showed that strain YHM-G could grow at a 3-Met level of 40 g/L (
Figure 1c), similar to that of
S. cerevisiae CEN.PK113-7D, indicating that strain YHM-G has the potential to produce more 3-Met [
53]. Overall, strain YHM-G can adapt well to different environments for the traditional brewing foods, such as
Baijiu. Therefore, strain YHM-G could be used for brewing these foods and increasing their contents of 3-Met.
3.4. Optimization of Culture Conditions by Single Factor Design
Although producing 3-Met by yeasts from sugars via de novo biosynthesis has been rare, previous reports have emphasized that its biosynthesis by yeasts requires sugars [
2,
3]. Therefore, the effect of glucose concentration on 3-Met production was evaluated: it increased initially as the glucose concentration increased and then decreased at higher concentrations. The yield of 3-Met was highest when the concentration range was 30–60 g/L (
Supplementary Figure S2a), which agreed with previous reports [
19]. When glucose is used as a carbon source, it provides energy for yeast growth and its concentration influences the rate of growth and the metabolic pathway and intensity. At a low glucose concentration, the biosynthesis of 3-Met was affected by a slow growth rate and fewer cells [
54]. Although previous reports have shown that a higher glucose concentration was conducive to accumulating 3-Met, an excessive concentration can change the metabolic pathway because the oxygen concentration was insufficient due to rapid cell growth [
11,
55]. Some studies have also shown that more ethanol and other higher alcohols may be produced at an excessive glucose concentration, which would be toxic to cells in combination with 3-Met [
43].
Yeast extracts contain many vitamins and microelements, which can be used for yeast growth and affect the accumulation of yeast metabolites [
8,
43,
56]. In the present study, 3-Met production was lower when yeast extract was absent because strain YHM-G was not able to grow well (
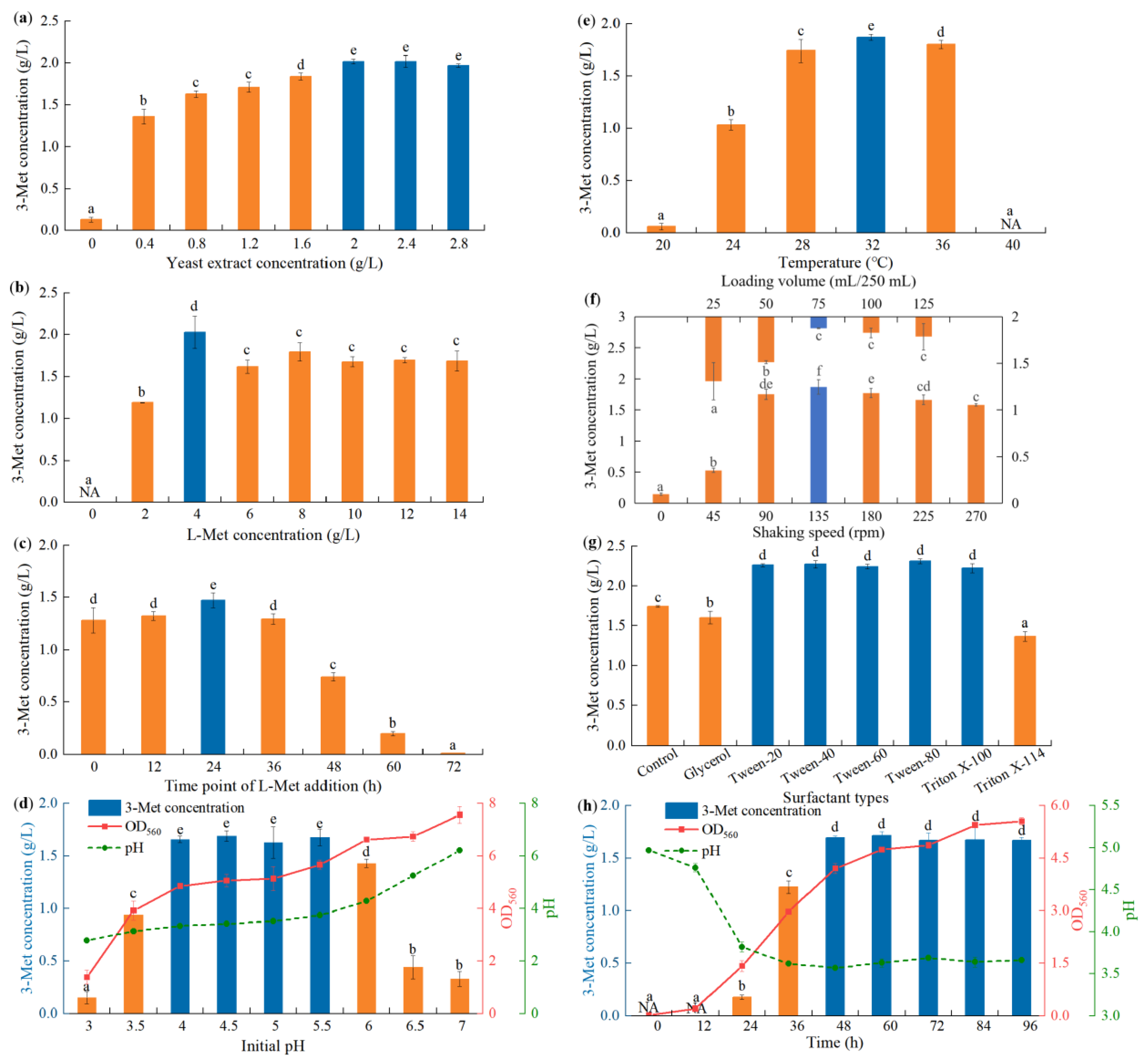
Figure 2a). Supplementation with yeast extract increased 3-Met production compared with no addition, with the yield increasing as its concentration increased. Strain YHM-G was able to produce higher levels of 3-Met when 2–2.8 g/L of yeast extract was present with the highest production of 2.02 g/L when 2.0 g/L of yeast extract was used (
Figure 2a). At a concentration of less than 2.0–2.4 g/L, yeast extract was able to be introduced as a nutrient to promote the growth of strain YHM-G [
8]. While as the concentration of yeast extract increased, the bioproduction of 3-Met decreased. Previous reports have shown that almost all 3-Met is biosynthesized from L-Met as the precursor via the Ehrlich pathway, which can be affected by other nitrogen sources during this process [
2,
8]. Thus, under an excessive yeast extract concentration, partial yeast extraction was applied as the yeast assimilable nitrogen to cause nitrogen catabolite repression to the Ehrlich pathway, leading to a decrease in production [
5,
8,
57,
58]. An excessive addition would lead to rapid cell growth, resulting in excessive substrate consumption for cell reproduction but reducing the accumulation of metabolites. A similar observation has also been reported in previous studies where the effect of yeast extract on 3-Met bioproduction was higher at low concentrations of yeast extract [
2,
3,
8]. It should be noted that the optimum concentration of yeast extract for 3-Met production by different yeasts varies, possibly because of their different metabolic capabilities [
2,
3,
5].
As L-Met is the precursor for 3-Met synthesis in the Ehrlich pathway, its concentration significantly influences 3-Met synthesis by yeasts [
5,
59]. Like previous reports, the present study found that no 3-Met was produced when L-Met was not present, thus confirming the importance of L-Met for the biosynthesis of 3-Met and the unlikeness of successful 3-Met biosynthesis from de novo sugars by strain YHM-G (
Figure 2b) [
2]. More 3-Met was synthesized as L-Met increased in the fermentation medium as it is an essential precursor for 3-Met production. It has been reported that increasing the L-Met concentration can enhance the formation of 3-Met, but in the present study, the yield reached a peak of 4 g/L, and when L-Met concentrations were >4 g/L, the production of 3-Met decreased slightly (
Figure 2b) [
3,
5,
8]. The reason was that an excessive L-Met concentration would strengthen demethiolation, thereby reducing the effective L-Met concentration and intermediate metabolites in the Ehrlich pathway [
3,
5]. Some studies have also shown that the nitrogen content is positively correlated with the fusel acid content, so more 3-methylthiopropionic acid was produced via the Ehrlich pathway at high L-Met concentrations, which was more toxic to the cells [
3,
5,
60,
61]. Regarding the economics of production, the optimum L-Met concentration in most studies was between 1 and 5 g/L. The results of the present study were also in this range, and the differing optimal L-Met concentrations for 3-Met production with different yeasts may have been caused mainly by differences in the characteristics of the strains and also by the slight differences between the initial fermentation mediums [
3,
5,
6,
8].
Although our results have confirmed the importance of L-Met for 3-Met synthesis by strain YHM-G, the question arises as to when adding L-Met could promote 3-Met synthesis. At present, no information detailing the optimal time for L-Met addition has been reported. The optimal time at which L-Met should be added for 3-Met production by strain YHM-G was determined by adding it at different culture times. 3-Met production was significantly higher when L-Met was added after 24 h of culture compared with adding at the start of culture, with no significant increase in production at 12 or 36 h but lower production at other times (
Figure 2c). The results also showed that 3-Met production decreased gradually as the time of addition was greater than 24 h (
Figure 2c). This was due to the overall effect of several factors: (1) L-Met is not only required for 3-Met production but is also an amino acid required for optimum growth of yeast; (2) adding L-Met later would shorten the time available for the transformation and utilization of L-Met to 3-Met; (3) the conversion of L-Met to 3-Met via the Ehrlich pathway relies on either ATP or the electrochemical gradient so that adding L-Met at the beginning of fermentation would inhibit cell growth and adding too late would lead to insufficient ATP because of the high consumption of glucose and dissolved oxygen [
5]; and (4) yeast extract contained in the fermentation medium can take over the role of L-Met as an amino acid for growth when added later. For strain YHM-G, when L-Met was added at 24 h of culture, L-Met can be fully utilized as a precursor and amino acid, so that the state of the cells and energy storage in the system was optimal.
Studies have shown that fluctuations in the environmental pH can stress microorganisms, thus changing the permeability of cell membranes and the activity of enzymes. The pH can change the state of nutrients in the system and affect the utilization of nutrients by microorganisms, resulting in different growth states and products for the microorganisms [
8,
25,
62,
63]. The optimum pH for 3-Met production was measured over a range from pH 3 to 7. The yield of 3-Met from strain YHM-G increased rapidly at a pH value between 3 to 4 and was optimal at a pH value between 4.0 and 5.5 (
Figure 2d). The yield decreased with increasing pH when the pH was ≥6 (
Figure 2d), which was consistent with the study of Seow et al. [
2] but not with that of Matthew et al. [
5]. The result showed that an initial of pH 4.0–5.5 would result in a final pH between 3.34–3.74, while when the initial pH was set at 3.0–3.5 and 6.0–7.0 and the final pH was 2.78–3.15 and 4.2–6.2, respectively. In addition, the result also illustrated that the final fermentation biomass was positively correlated with the initial pH (
Figure 2d). Apparently, the optimal pH for 3-Met production was slightly lower than the optimal pH for the growth of strain YHM-G. Thus, the reason for these results was that enzyme activity in the Ehrlich pathway for 3-Met production was probably optimal under slightly acidic conditions due to its long-term survival in the slightly acidic environment of
Baijiu making [
2,
5].
Temperature is an important factor affecting the growth of microorganisms and the production of metabolites. If the temperature is too low, microbial growth will be greatly compromised; an excessive temperature may lead to cell death or enzyme inactivation, discouraging the accumulation of the end-products [
8,
64,
65]. Because of the growth temperature range of strain YHM-G, the optimum temperature for 3-Met production was measured from 20 to 40 °C. The 3-Met production increased as the temperature of incubation increased from 20 to 32 °C (
Figure 2e), which was consistent with the study by Seow et al. [
2]. As previously reported, the relationship between temperature and 3-Met production was not linear [
2]. The increases in 3-Met formation were large, at 300% and 150%, for an increase in temperature of 4 °C, from 20 to 24 °C, and from 24 to 28 °C (
Figure 2e), respectively, which was not consistent with a previous study [
2], mainly because of the different strains. The highest yield of 3-Met occurred at 32 °C (
Figure 2e), similar to
K. lactis KL71 [
2], which was consistent with their optimum growth temperature [
2,
5]. At this temperature, growth was faster and the enzymes for 3-Met biosynthesis may have exhibited a higher activity [
2]. A reduction in the growth rate and activity of enzymes related to 3-Met synthesis led to a decrease in 3-Met production at high temperatures [
8,
66].
Although yeasts are facultative anaerobic microorganisms, their growth and most biological metabolic activities benefit from aerobic conditions [
5,
43]. The metabolic pathways for different metabolites differ and the oxygen concentration required also varies [
25,
43]. In shaking flask fermentation, the shaking speed and loading volume are the two parameters that most directly affect oxygen concentration during the fermentation process. They both regulate the production of metabolites by affecting oxygen concentration [
43]. Therefore, the effect of shaking speed and loading volume on 3-Met synthesis were investigated. The results showed that 3-Met produced by strain YHM-G first increased then decreased with increasing shaking speed or loading volume (
Figure 2f). With increasing shaking speed or decreasing loading volume, the oxygen concentration in the fermentation system rapidly increased growth and invigorated the metabolism, significantly increasing the yield of 3-Met [
5]. When the shaking speed was 135 rpm or the loading volume was 75 mL/250 mL, the yield of 3-Met reached its highest value, at almost 10 times that achieved in an unaerated fermentation (shaking speed = 0 rpm), which was consistent with a previous study (
Figure 2f) [
2]. With a continuing increase in shaking speed or decrease in loading volume, the 3-Met production decreased (
Figure 2f). The possible reasons for this were that the increase in shaking speed or the decrease in loading volume increased the shear force and damaged the cells, thus affecting the metabolism of the yeast, and the consequent excessive oxygen concentration caused more methional produced in the Ehrlich pathway to be oxidized to 3-methylthio-1-propanoic acid with less available for producing 3-Met [
3]. Thus, it is very important to maintain an appropriate oxygen concentration to ensure the cell biomass and reaction direction in the Ehrlich pathway by regulating the shaking speed and loading volume to ensure a high yield of 3-Met.
Inoculum size is also an important parameter often considered when investigating the biosynthetic metabolites during microbial fermentation. It affects not only the yield of the target products but also the fermentation cycle [
67]. An appropriate inoculum size can shorten the fermentation cycle while maintaining the yield. A low inoculum volume, such as 0.1% (the cell density was adjusted to a concentration of 1 × 10
6 cells/mL by using a blood cell counting plate), can lead to low 3-Met production owing to the fact that longer delay period may lead to lower biomass, whereas a high inoculum volume, such as 6.4%, would also decrease the yield because excessive cell growth may lead to insufficient oxygen supply (
Supplementary Figure S2b) [
67]. There was no significant difference in 3-Met production when the inoculum size was between 0.2% and 3.2% and the yield was higher because of the superior growth rate and biomass and sufficient nutrients (
Supplementary Figure S2b) [
43]. Many previous studies have shown that the optimal inoculum size was also in this range, for example,
P. kudriavzevii YF1702 for producing 2-phenylethanol [
43] and
Bacillus licheniformis NCIMB 8059 for producing 2,3-butanediol [
68].
The cell membrane is an important component of microorganisms as it maintains cell structure and activity. It is also an important channel for material and energy transfer between cells and the surrounding environment. Changes in the permeability of the cell membrane would cause the absorption of nutrients and the secretion and extravasation of intracellular substances, thus affecting cell metabolism. The cell membrane consists of a phospholipid bilayer, so some amphiphilic surfactants, such as Tween and Triton, would affect its permeability and microbial metabolites [
69,
70]. Previous studies have shown that different surfactants have different effects on different metabolites of the same microorganism or on the same metabolites of different microorganisms [
43]. Therefore, it was necessary to study the effect of surfactant type and concentration on 3-Met production by strain YHM-G, as no information is available at present. The results showed that both Triton X-100 and the Tween series were able to promote 3-Met production from strain YHM-G in a similar way, while glycerol and Triton X-114 inhibited 3-Met production (
Figure 2g). This was because different types of surfactants had different effects on the cell membrane of strain YHM-G, which also confirmed previous reports on the effects of surfactants on microbial metabolism [
43]. Tween-80 led to the maximum yield of 3-Met at 20.3% higher than the control (
Figure 2g). Varying the Tween-80 concentration had little effect on 3-Met production (
Supplementary Figure S2c), possibly because Tween-80 was compatible with the cells of strain YHM-G; even at high concentrations it still increased cell membrane permeability and promoted the dissolved oxygen level in the cells but did not permeabilize the cell membrane [
70]. From considering economic factors, a lower Tween 80 concentration was chosen.
As the fermentation cycle extends, metabolites continuously accumulate, but during this process, with the consumption of nutrients, certain harmful substances also accumulate and environmental growth factors, such as pH, can change and microbial metabolic stress reactions will occur, utilizing some metabolites and converting them into other substances, resulting in a reduction in metabolites [
71]. Therefore, a suitable fermentation cycle is critical for successful fermentation. The results presented in
Figure 2h show the effect of culture time on 3-Met production by strain YHM-G. No 3-Met was detected during the first 12 h, possibly because the strain was multiplying during this period (
Figure 2h). This may also be a reason for optimizing the time when L-Met is added or because adding L-Met at the initial stage of fermentation may impose a certain pressure on cell reproduction and growth. Previous reports have shown that the production of 3-Met may start during the initial growth phase of
K. lactis, which was different from strain YHM-G [
72]. The 3-Met concentration increased rapidly between 12 and 48 h then the production level remained essentially unchanged between 48 and 96 h (
Figure 2h), which was consistent with
S. cerevisiae EC-1118 [
8]. It was observed that 3-Met was produced mainly during the early logarithmic stage of yeast growth by analyzing the growth curve and the 3-Met production curve. Production did not necessarily start if or when the growth of strain YHM-G was limited, which was consistent with a previous study [
72]. 3-Met production also mainly occurred at a pH of 3.6, which may be conducive to its synthesis and was similar to results on the initial pH of the medium (
Figure 2d and h).
3.7. Optimization of the Production Conditions by RSM Design
According to the results of PB and the steepest ascent path design, a three-factor (time point of L-Met addition, glucose concentration and yeast extract concentration) and three-level BBD were used to optimize the best fermentation conditions for 3-Met production by YHM-G. Various maximum and minimum levels of three factors were used for the 15 experimental runs in
Table 4. The results showed considerable variation in 3-Met production with a maximum of 3.04 g/L in test number 10 and a minimum of 1.04 g/L in test number 4.
A second order polynomial equation E2 was obtained to explain 3-Met production by applying multiple regression analysis on the experimental data:
where Y is the predicted response and A, B and C are the time point of L-Met addition, glucose concentration and yeast concentration, respectively.
The ANOVA (
F-test) shows that the model was statistically significant. The
R2 was 0.9709, indicating that the sample variation of 97.09% for 3-Met production was attributed to the independent variables and only about 2.93% of the total variation was unexplained by the model (
Table 7). Furthermore, the model terms of A (the time point of L-Met addition); B (glucose concentration); C (yeast extract concentration); and A
2, B
2 and C
2 were significant at the 5% level, and the lack of fit was not significant with a
p value of 0.2658 (
Table 7). Thus, the model was adequate for prediction with the range of factors used.
The
p values and coefficients suggested the independent B (glucose concentration), and C (yeast extract concentration) had a significant positive effect on 3-Met production (
Table 7), indicating that they could cause an increase linearly in 3-Met production. Additionally, factor A (time point of L-Met addition) significantly negatively affected 3-Met production, indicating that factor A caused 3-Met production to decrease (
Table 7). The quadric term of A, B and C also had a significant effect, whereas there was no evidence of interaction among them (
Table 7).
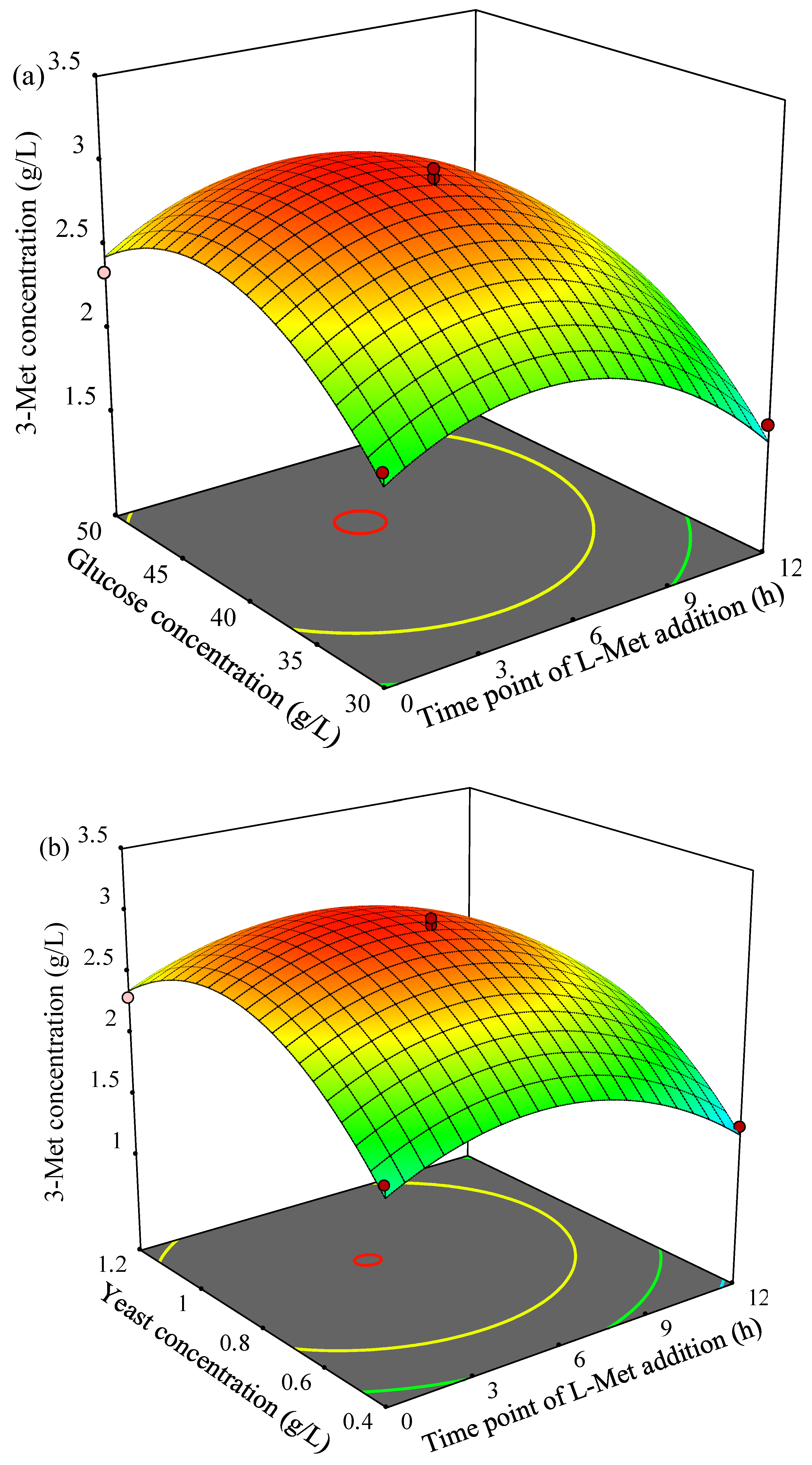
The three-dimensional (3D) response surface plots for 3-Met production described by the above model were drawn to illustrate the effects of the independent variables and to depict the interactions between two variables by keeping the third variables at their zero levels (
Figure 3). As shown in
Figure 3a, 3-Met production increased first then decreased gradually as the time point of L-Met addition or glucose concentration increased, and the higher yield of 3-Met was obtained when time point of L-Met addition was at 4.2–5.6 h and glucose concentration was in the range of 41.3–43.8 g/L. The similar trend of 3-Met production was observed in
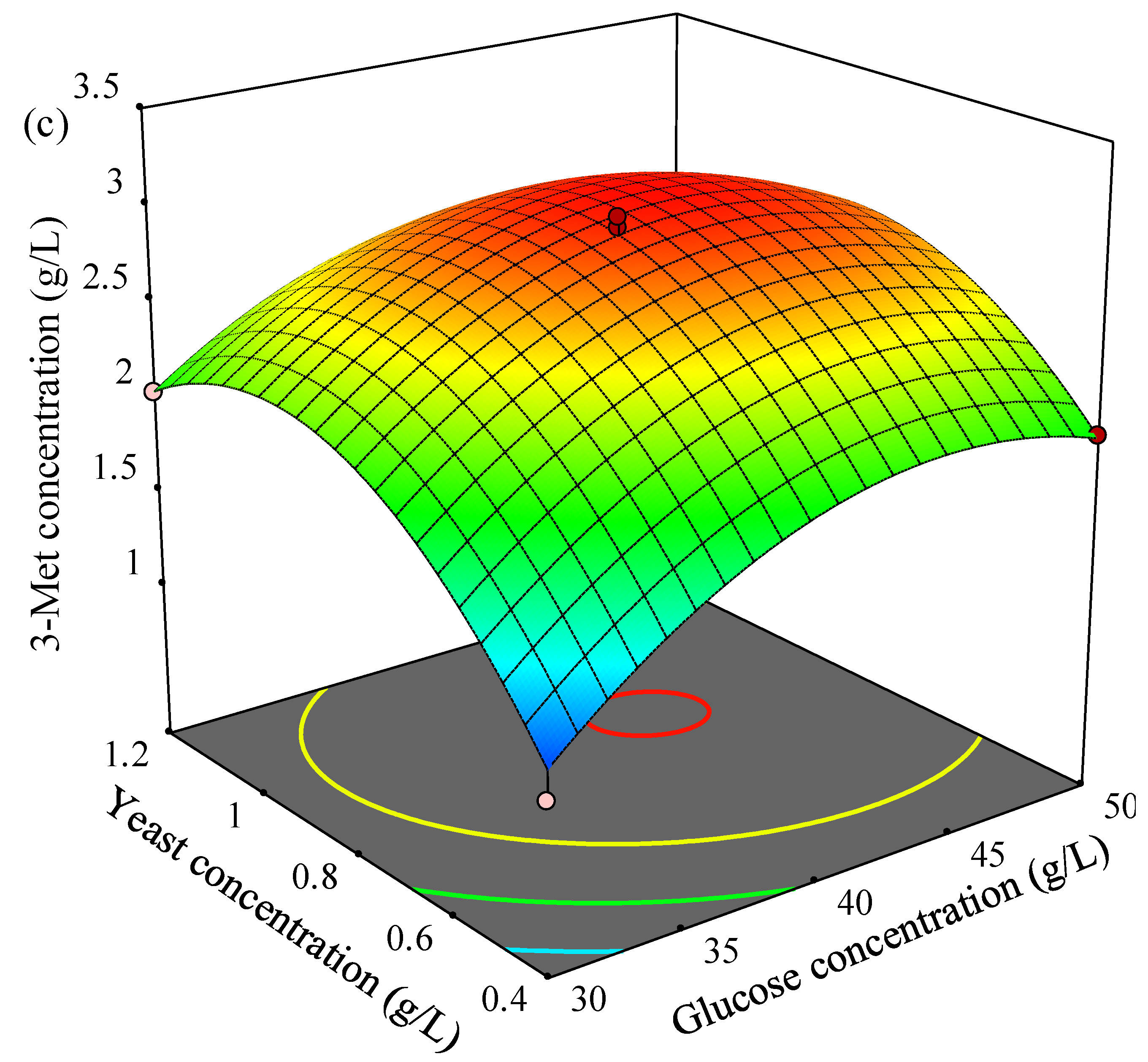
Figure 3b as time point of L-Met addition or yeast concentration increased, and the yield of 3-Met was higher 3.0 g/L when time point of L-Met addition was in the range of 4.4–5.2 h and yeast concentration was in the range of 0.88–0.92 g/L. In addition, 3-Met production increased then decreased as glucose concentration or yeast concentration increased, and more than 3.0 g/L of 3-Met was obtained by YHM-G when glucose concentration was in the range of 40.7–44.6 g/L and yeast concentration was in the rage of 0.82–0.95 g/L (
Figure 3c).
The maximum predicted value of 3-Met production was 3.03 g/L derived by the Design expert 11 software with the following critical values: A (time point of L-Met addition) = 4.9 h, B (glucose concentration) = 42.7 g/L and C (yeast extract concentration) = 0.9 g/L. According to the results of the statistically designed experiments, the optimized process parameters were: 42.7 g/L glucose, pH 6, 0.9 g/L yeast extract, 6 g/L L-Met, 28 °C, shaking speed 210 rpm, loading volume 50 mL/250 mL, inoculum size 0.5% (
v/v), time point of L-Met addition 4.9 h, culturing period 48 h and 2.5 g/L Tween-80. Validation experiments were performed at the optimal level, and the yield of 3-Met was 3.16 g/L, which was 88.1% higher than the value before optimization (1.68 g/L,
Table 5). Additionally, YHM-G has obvious advantages in terms of yield compared with other reported 3-Met-producing natural yeast strains (
Table 1).
3.8. Aroma Production
Flavor substances affect food quality to a certain extent, with the unique flavor characteristics of fermented foods being mainly transformed from the ingredients in the substrate by microorganisms [
73]. Thus, microorganisms with excellent flavor-producing characteristics have certain advantages for producing fermented foods. Strain YHM-G is a flavor-producing yeast screened from the
Baijiu-making environment. The analysis of its flavor-producing characteristics would encourage its application in fermented foods. The results showed that a total of 23 flavor substances, including seven alcohols, five phenols, three esters, two alkanes, one ketone, one aldehyde, one olefin, one furan, one sulfur-containing compound and one nitrogen-containing compound, were detected in SHM after culturing strain YHM-G for 48 h (
Table 8). Compared with the volatile components in SHM, there were fewer common substances in the fermentation broth and SHM, except for phenylethanol, dibutyl phthalate and benzothiazole. The content changes in two of these three flavor substances, dibutyl phthalate and benzothiazole in the fermentation broth would have been derived from SHM, but for phenylethanol, unlike SHM, most arose mainly from de novo synthesis by strain YHM-G through the metabolism of sugars [
43]. Ethyl palmitate, ethyl trans oleate, ethyl linoleic acid, aniline, 2,4-dimethylbenzaldehyde and 2-methylpyrazine found in SHM was not detected in the fermentation broth, so they may have been converted into other flavor substances after fermentation by strain YHM-G, such as 2,4-dimethylbenzaldehyde being converted into 2,4-di-t-butylphenol. Similarly, flavor substances, which was not found in SHM were present in the fermentation broth. These substances include isopentanol, citronellol, p-vinyllignocerol and 2,3-dihydrobenzofuran, which were produced by strain YHM-G metabolizing substances in SHM. Notably, no 3-Met was detected in the fermentation broth since strain YHM-G could not produce it without the L-Met precursor, which was consistent with the results on optimizing the amount of L-Met optimization (
Figure 3b). This observation further demonstrated that 3-Met was produced by strain YHM-G via the Ehrlich pathway not via a de novo pathway from sugars, similar to most previous reports [
74]. Although strain YHM-G could not synthesize 3-Met without the L-Met precursor, it could be produced by the strain by constructing a workable synthetic microbial community, as successfully reported [
7]. Among the flavor substances produced by strain YHM-G, higher alcohols, such as isobutanol, isoamyl alcohol and 2-propyl-1-pentanol, have pine, alcoholic and mushroom aromas, which can be important contributions to the flavor characteristics of fermented food [
75]. Terpene alcohols, such as citronellol, trans-nerolidol and farnesol, can impart floral and fruit aromas to fermented food, and provide certain physiological functional characteristics [
76,
77]. Phenylethanol and its corresponding ester (phenylethyl acetate) can impart rose and honey aromas to fermented food [
78]; C-nonanolactone a coconut aroma [
79]; and phenolic substances produced by strain YHM-G, such as guaiacol and 4-ethyl-2-methoxyphenol, fruit, flower and sweet aromas, with antioxidant properties for food [
78,
80]; and 2,3-dihydrobenzofuran and N,N-dibutyl formamide can give foods a roasted flavor [
81,
82]. In conclusion, strain YHM-G has good potential for application in fermented foods.







{kind=link}
{kind=link}
{kind=link}
{kind=link}