
An Exploration of Pepino (Solanum muricatum) Flavor Compounds Using Machine Learning Combined with Metabolomics and Sensory Evaluation

{kind=link}
{kind=link}
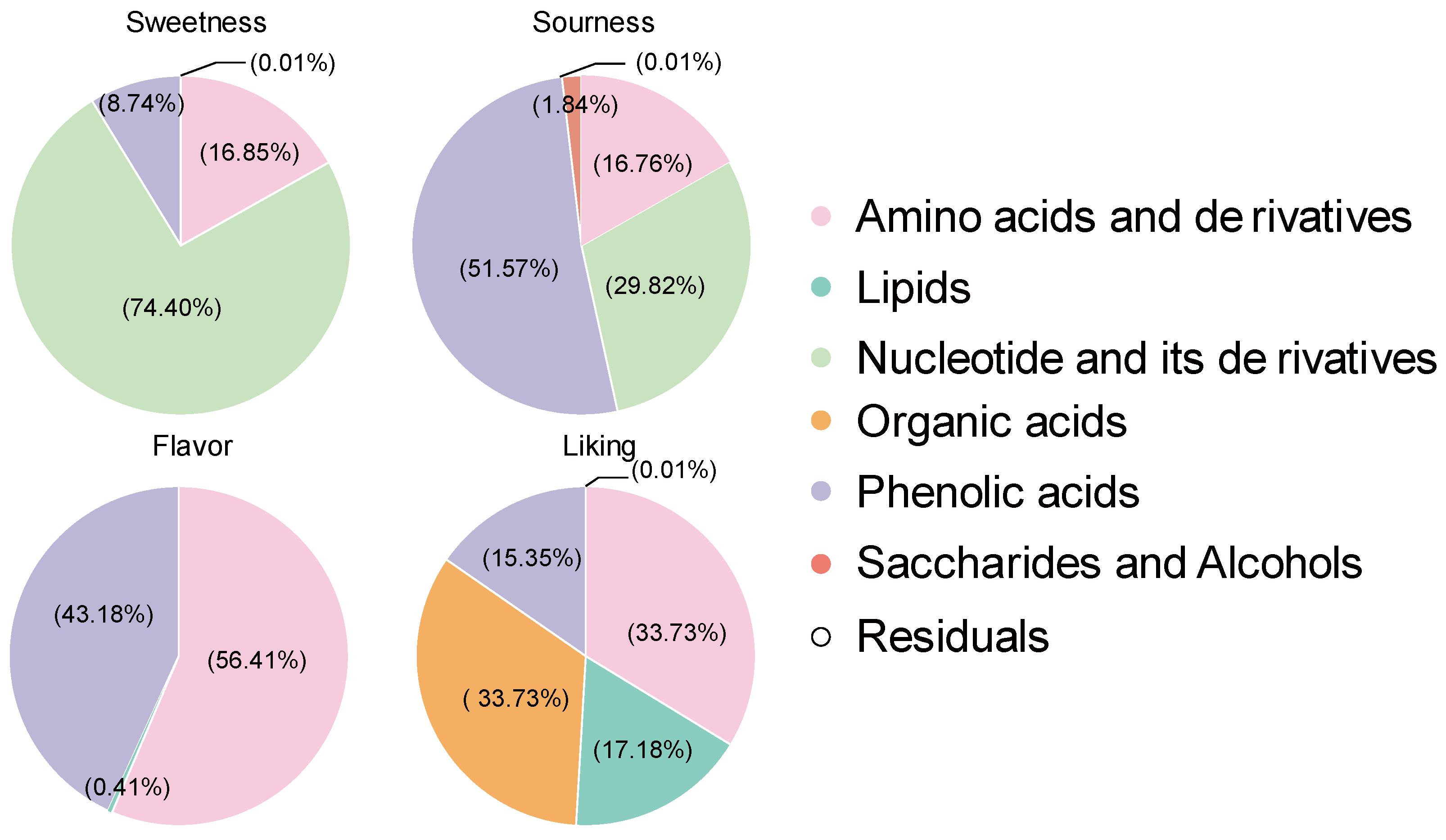{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Methods
2.2.1. Sensory Evaluation of Pepino
2.2.2. Data Analysis
3. Results
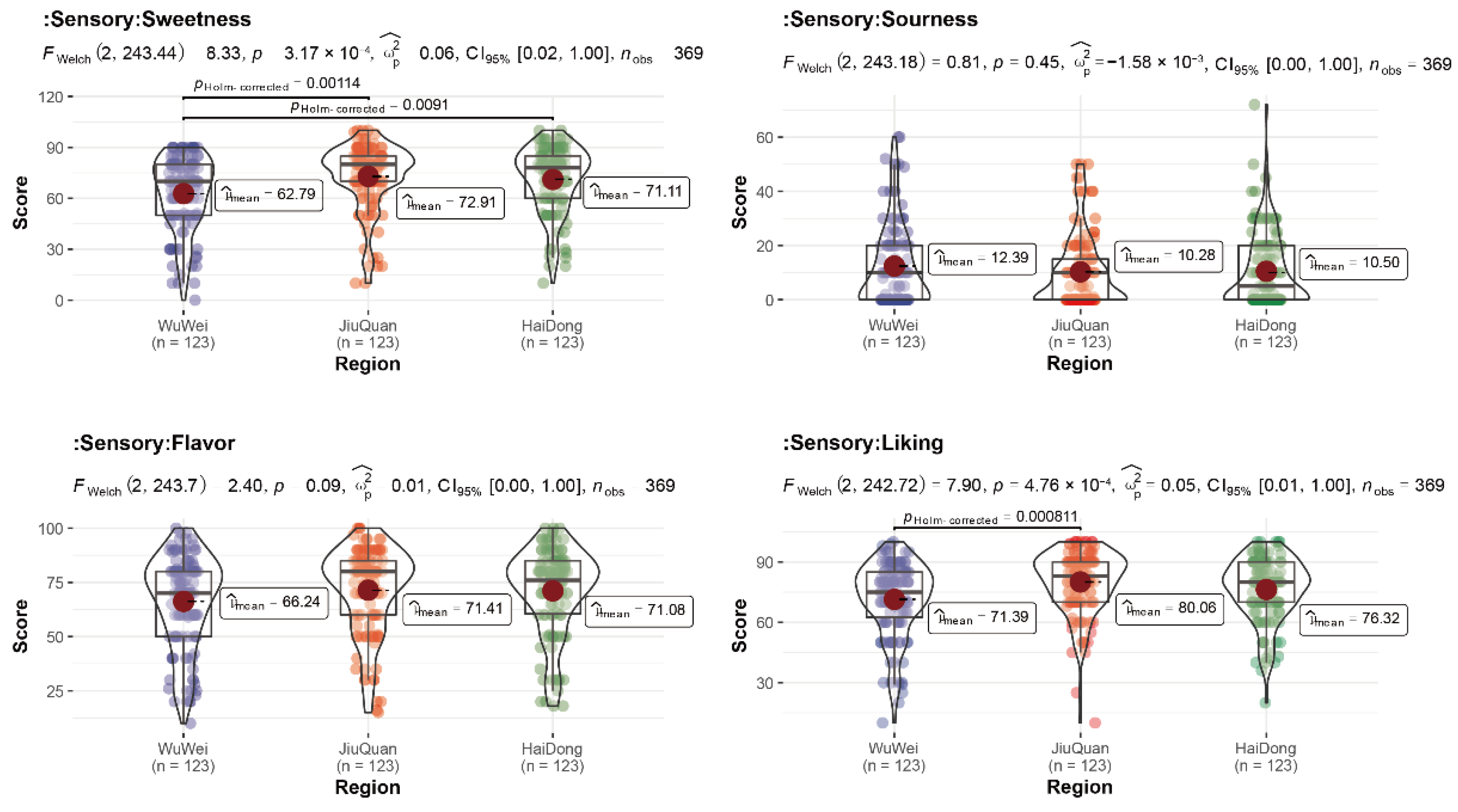
3.1. Statistics of Sensory Evaluation
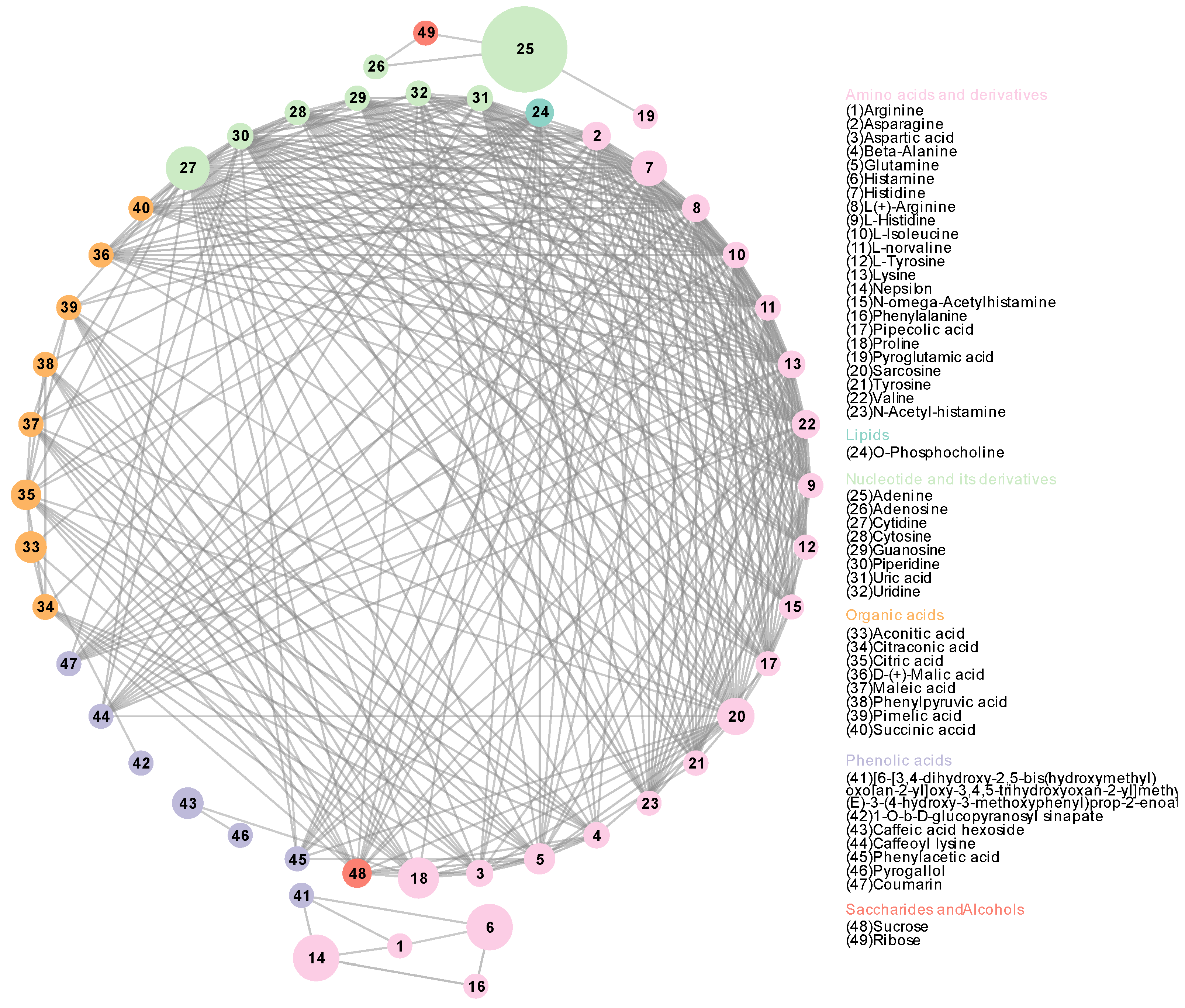
3.2. Metabolic Network
3.3. Contribution of Various Metabolites to Flavor Perception
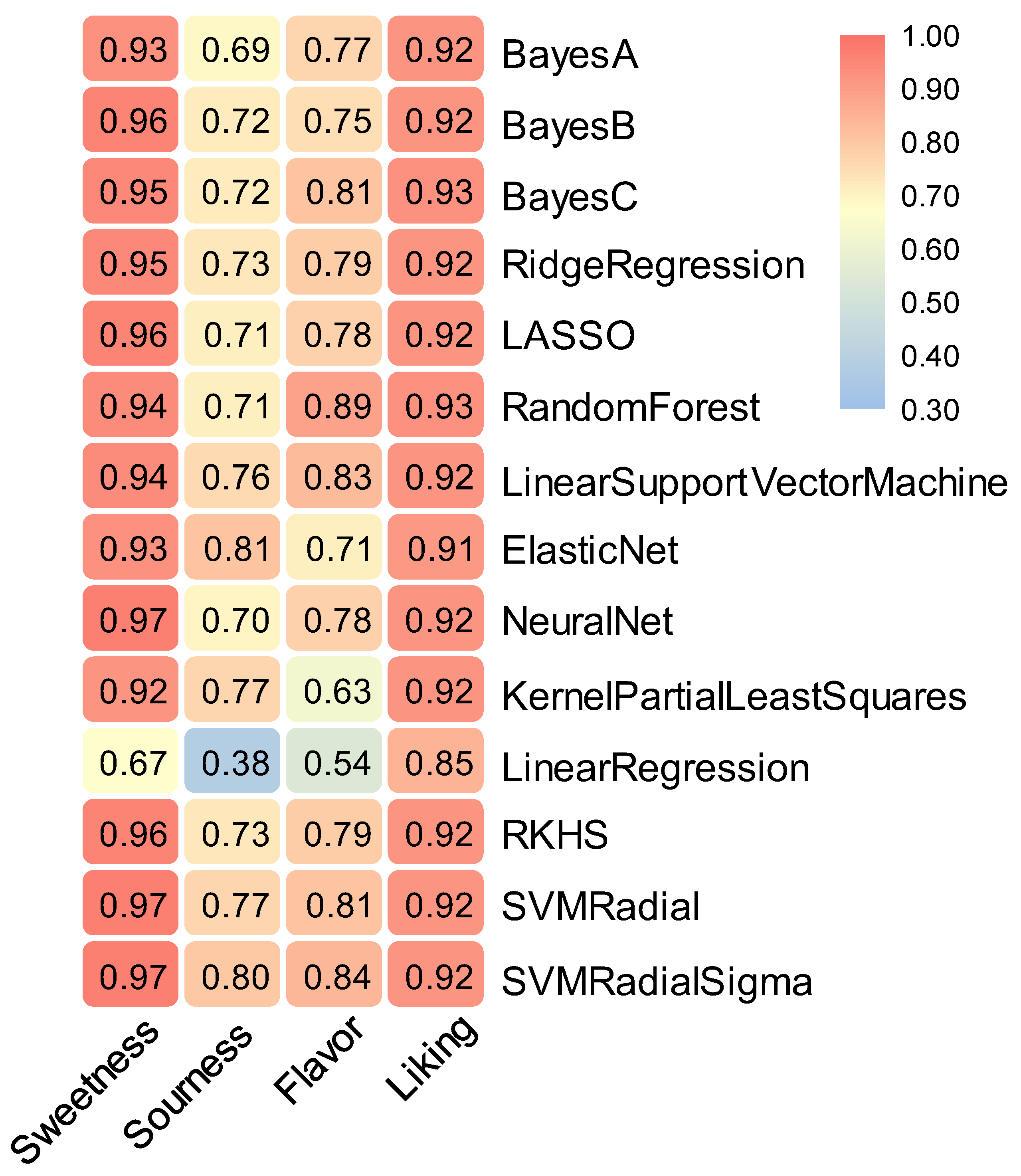
3.4. Consumer Flavor Preference Prediction
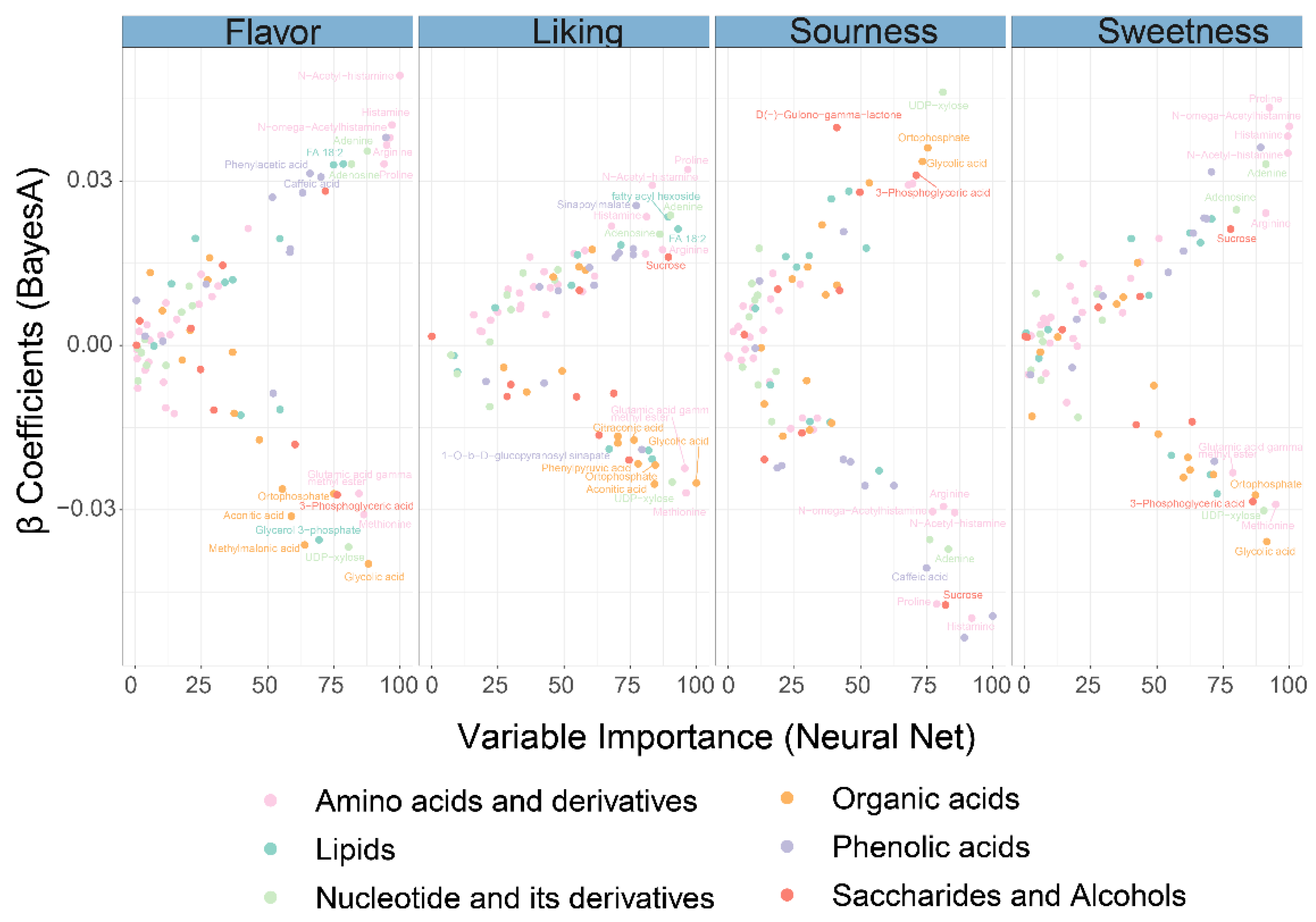
3.5. Important Metabolites Associated with Sensory Characteristics
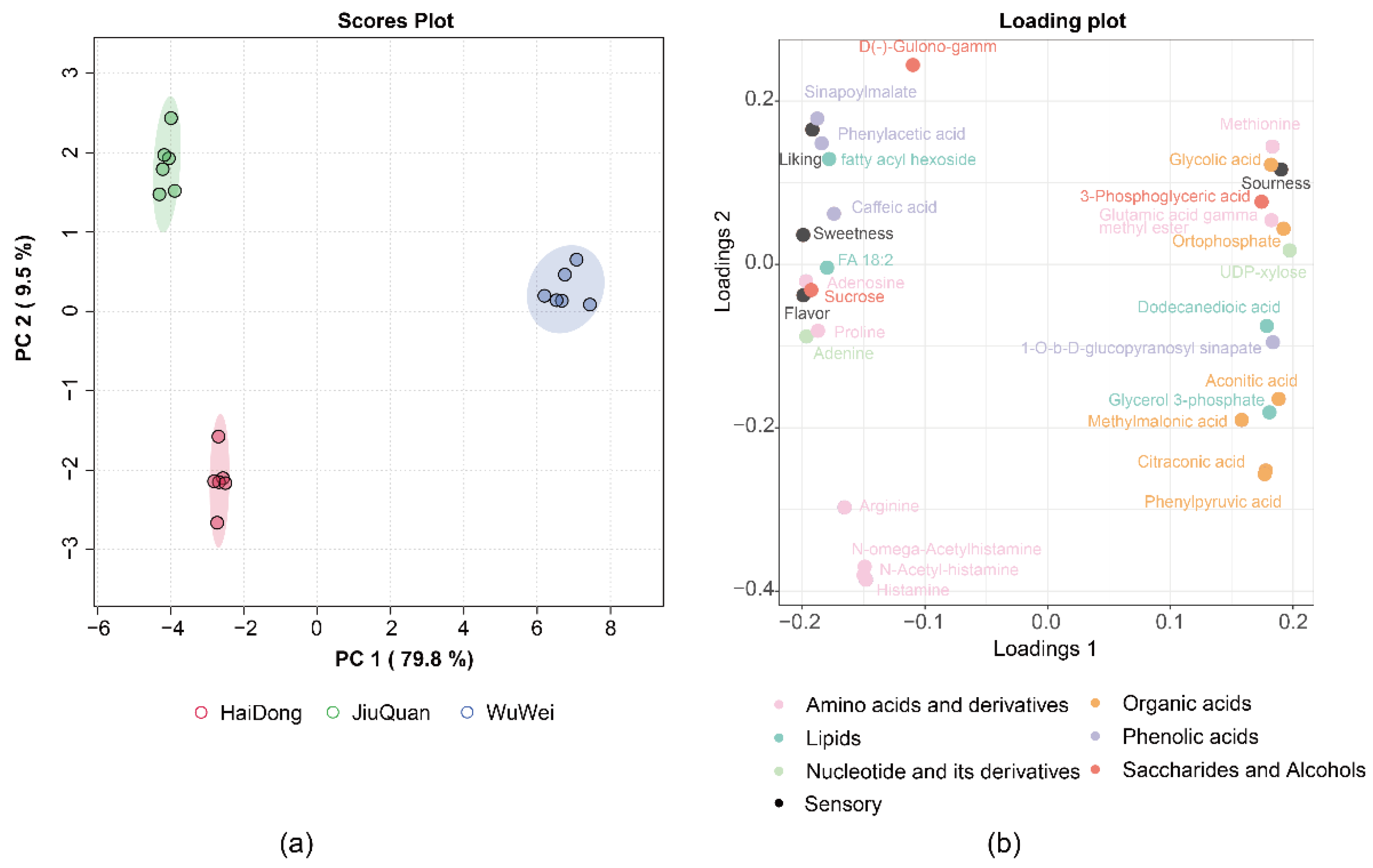
3.6. Metabolites Distinguish Pepino Regional Origin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prohens, J.; Ruiz, J.J.; Nuez, F. The Pepino (Solanum Muricatum, Solanaceae): A “New” Crop with a History. Econ. Bot. 1996, 50, 355–368. [Google Scholar] [CrossRef]
- Virani, D.; Chaerunnisa, N.N.; Suarsi, I.; Dachlan, D.M.; Thahir, A.I.A. Pepino Extract (Solanum Muricatum Ait.) on HDL and LDL in Type 2 Diabetic Rats. Enferm. Clin. 2020, 30, 163–166. [Google Scholar] [CrossRef]
- Shathish, K.; Guruvayoorappan, C. Solanum Muricatum Ait. Inhibits Inflammation and Cancer by Modulating the Immune System. J. Cancer Res. Ther. 2014, 10, 623. [Google Scholar]
- Pizarro, P.; Villena, G.L.; Duviols, P. Relación Del Descubrimiento y Conquista de Los Reinos Del Perú; Pontificia Universidad Católica del Perú Lima: San Miguel, Peru, 1986; ISBN 84-89292-71-X. [Google Scholar]
- Nuez, F.; Morales, R.; Ruiz, J.J.; de Córdova, P.F.; Valdivieso, E.; González, F.I. Recolección de Especies Hortícolas En Ecuador. Plant Genet. Resour. Newsl. 1993, 96, 29–33. [Google Scholar]
- Yang, S.; Zhu, H.; Huang, L.; Zhang, G.; Wang, L.; Jiang, X.; Zhong, Q. Transcriptome-Wide and Expression Analysis of the NAC Gene Family in Pepino (Solanum Muricatum) during Drought Stress. PeerJ 2021, 9, e10966. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Wang, L.; Zhang, G.; Yang, S.; Zhong, Q. Pepino (Solanum Muricatum) Metabolic Profiles and Soil Nutrient Association Analysis in Three Growing Sites on the Loess Plateau of Northwestern China. Metabolites 2022, 12, 885. [Google Scholar] [CrossRef]
- Dafny-Yalin, M.; Glazer, I.; Bar-Ilan, I.; Kerem, Z.; Holland, D.; Amir, R. Color, Sugars and Organic Acids Composition in Aril Juices and Peel Homogenates Prepared from Different Pomegranate Accessions. J. Agric. Food Chem. 2010, 58, 4342–4352. [Google Scholar] [CrossRef]
- Colaric, M.; Veberic, R.; Stampar, F.; Hudina, M. Evaluation of Peach and Nectarine Fruit Quality and Correlations between Sensory and Chemical Attributes. J. Sci. Food Agric. 2005, 85, 2611–2616. [Google Scholar] [CrossRef]
- Cohen, S.; Itkin, M.; Yeselson, Y.; Tzuri, G.; Portnoy, V.; Harel-Baja, R.; Lev, S.; Sa‘Ar, U.; Davidovitz-Rikanati, R.; Baranes, N. The PH Gene Determines Fruit Acidity and Contributes to the Evolution of Sweet Melons. Nat. Commun. 2014, 5, 4026. [Google Scholar] [CrossRef] [Green Version]
- Aboud, O.A.; Weiss, R.H. New Opportunities from the Cancer Metabolome. Clin. Chem. 2013, 59, 138–146. [Google Scholar] [CrossRef] [Green Version]
- McGarrah, R.W.; Crown, S.B.; Zhang, G.-F.; Shah, S.H.; Newgard, C.B. Cardiovascular Metabolomics. Circ. Res. 2018, 122, 1238–1258. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martínez-González, M.A.; Salas-Salvadó, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-Analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef]
- Bhowmick, S.; Mazumdar, A.; Moulick, A.; Adam, V. Algal Metabolites: An Inevitable Substitute for Antibiotics. Biotechnol. Adv. 2020, 43, 107571. [Google Scholar] [CrossRef]
- Yu, Y.; Guan, J.; Xu, Y.; Ren, F.; Zhang, Z.; Yan, J.; Fu, J.; Guo, J.; Shen, Z.; Zhao, J. Population-Scale Peach Genome Analyses Unravel Selection Patterns and Biochemical Basis Underlying Fruit Flavor. Nat. Commun. 2021, 12, 3604. [Google Scholar] [CrossRef]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C. Rewiring of the Fruit Metabolome in Tomato Breeding. Cell 2018, 172, 249–261.e12. [Google Scholar] [CrossRef] [Green Version]
- Cebolla-Cornejo, J.; Roselló, S.; Valcárcel, M.; Serrano, E.; Beltrán, J.; Nuez, F. Evaluation of Genotype and Environment Effects on Taste and Aroma Flavor Components of Spanish Fresh Tomato Varieties. J. Agric. Food Chem. 2011, 59, 2440–2450. [Google Scholar] [CrossRef]
- Karppinen, K.; Zoratti, L.; Nguyenquynh, N.; Häggman, H.; Jaakola, L. On the Developmental and Environmental Regulation of Secondary Metabolism in Vaccinium Spp. Berries. Front. Plant Sci. 2016, 7, 655. [Google Scholar] [CrossRef] [Green Version]
- Ferrão, L.F.V.; Johnson, T.S.; Benevenuto, J.; Edger, P.P.; Colquhoun, T.A.; Munoz, P.R. Genome-wide Association of Volatiles Reveals Candidate Loci for Blueberry Flavor. New Phytol. 2020, 226, 1725–1737. [Google Scholar] [CrossRef]
- Cappai, F.; Benevenuto, J.; Ferrão, L.F.V.; Munoz, P. Molecular and Genetic Bases of Fruit Firmness Variation in Blueberry—A Review. Agronomy 2018, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Gonda, I.; Sun, H.; Ma, Q.; Bao, K.; Tieman, D.M.; Burzynski-Chang, E.A.; Fish, T.L.; Stromberg, K.A.; Sacks, G.L. The Tomato Pan-Genome Uncovers New Genes and a Rare Allele Regulating Fruit Flavor. Nat. Genet. 2019, 51, 1044–1051. [Google Scholar] [CrossRef]
- Alonge, M.; Wang, X.; Benoit, M.; Soyk, S.; Pereira, L.; Zhang, L.; Suresh, H.; Ramakrishnan, S.; Maumus, F.; Ciren, D. Major Impacts of Widespread Structural Variation on Gene Expression and Crop Improvement in Tomato. Cell 2020, 182, 145–161.e23. [Google Scholar] [CrossRef]
- Klee, H.J. Improving the Flavor of Fresh Fruits: Genomics, Biochemistry, and Biotechnology. New Phytol. 2010, 187, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Ferrao, L.F.V.; Sater, H.; Lyrene, P.; Amadeu, R.R.; Sims, C.A.; Tieman, D.M.; Munoz, P.R. Terpene Volatiles Mediates the Chemical Basis of Blueberry Aroma and Consumer Acceptability. Food Res. Int. 2022, 158, 111468. [Google Scholar] [CrossRef]
- Colantonio, V.; Ferrão, L.F.V.; Tieman, D.M.; Bliznyuk, N.; Sims, C.; Klee, H.J.; Munoz, P.; Resende Jr, M.F. Metabolomic Selection for Enhanced Fruit Flavor. Proc. Natl. Acad. Sci. 2022, 119, e2115865119. [Google Scholar] [CrossRef]
- Niazian, M.; Niedbała, G. Machine Learning for Plant Breeding and Biotechnology. Agriculture 2020, 10, 436. [Google Scholar] [CrossRef]
- van Dijk, A.D.J.; Kootstra, G.; Kruijer, W.; de Ridder, D. Machine Learning in Plant Science and Plant Breeding. Iscience 2021, 24, 101890. [Google Scholar] [CrossRef]
- Sun, S.; Wang, C.; Ding, H.; Zou, Q. Machine Learning and Its Applications in Plant Molecular Studies. Brief. Funct. Genom. 2020, 19, 40–48. [Google Scholar] [CrossRef]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D. Accurate Prediction of Protein Structures and Interactions Using a Three-Track Neural Network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef]
- Sperschneider, J. Machine Learning in Plant–Pathogen Interactions: Empowering Biological Predictions from Field Scale to Genome Scale. New Phytol. 2020, 228, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Koh, J.C.; Spangenberg, G.; Kant, S. Automated Machine Learning for High-Throughput Image-Based Plant Phenotyping. Remote Sens. 2021, 13, 858. [Google Scholar] [CrossRef]
- Tieman, D.; Bliss, P.; McIntyre, L.M.; Blandon-Ubeda, A.; Bies, D.; Odabasi, A.Z.; Rodríguez, G.R.; van der Knaap, E.; Taylor, M.G.; Goulet, C. The Chemical Interactions Underlying Tomato Flavor Preferences. Curr. Biol. 2012, 22, 1035–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinform. 2008, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Legeay, M.; Doncheva, N.T.; Morris, J.H.; Jensen, L.J. Visualize Omics Data on Networks with Omics Visualizer, a Cytoscape App. F1000Research 2020, 9, 157. [Google Scholar] [CrossRef] [Green Version]
- Butler, D.G.; Cullis, B.R.; Gilmour, A.R.; Gogel, B.J.; Thompson, R. ASReml-R Reference Manual Version 4; Vsn International Limited: Hemel Hempstead, UK, 2017. [Google Scholar]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Villanueva, R.A.M.; Chen, Z.J. Ggplot2: Elegant Graphics for Data Analysis; Taylor & Francis: Oxfordshire, UK, 2019; ISBN 1536-6367. [Google Scholar]
- Cullen, K.A.; Gentzke, A.S.; Sawdey, M.D.; Chang, J.T.; Anic, G.M.; Wang, T.W.; Creamer, M.R.; Jamal, A.; Ambrose, B.K.; King, B.A. E-Cigarette Use among Youth in the United States, 2019. Jama 2019, 322, 2095–2103. [Google Scholar] [CrossRef] [PubMed]
- Duan, N.; Bai, Y.; Sun, H.; Wang, N.; Ma, Y.; Li, M.; Wang, X.; Jiao, C.; Legall, N.; Mao, L. Genome Re-Sequencing Reveals the History of Apple and Supports a Two-Stage Model for Fruit Enlargement. Nat. Commun. 2017, 8, 249. [Google Scholar] [CrossRef]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational Research on Trichoderma: From’omics to the Field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef] [Green Version]
- Talhout, R.; van de Nobelen, S.; Kienhuis, A.S. An Inventory of Methods Suitable to Assess Additive-Induced Characterising Flavours of Tobacco Products. Drug Alcohol Depend. 2016, 161, 9–14. [Google Scholar] [CrossRef]
- Zhao, Y.; Zuo, J.; Yuan, S.; Shi, W.; Shi, J.; Feng, B.; Wang, Q. UV-C Treatment Maintains the Sensory Quality, Antioxidant Activity and Flavor of Pepino Fruit during Postharvest Storage. Foods 2021, 10, 2964. [Google Scholar] [CrossRef]
- Contreras, C.; Schwab, W.; Mayershofer, M.; González-Agüero, M.; Defilippi, B.G. Volatile Compound and Gene Expression Analyses Reveal Temporal and Spatial Production of LOX-Derived Volatiles in Pepino (Solanum Muricatum Aiton) Fruit and LOX Specificity. J. Agric. Food Chem. 2017, 65, 6049–6057. [Google Scholar] [CrossRef]
- Yu, X.; Xiao, J.; Chen, S.; Yu, Y.; Ma, J.; Lin, Y.; Li, R.; Lin, J.; Fu, Z.; Zhou, Q. Metabolite Signatures of Diverse Camellia Sinensis Tea Populations. Nat. Commun. 2020, 11, 5586. [Google Scholar] [CrossRef]
- Mofokeng, M.M.; Prinsloo, G.; Araya, H.T.; Du Plooy, C.P.; Sathekge, N.R.; Amoo, S.O.; Steyn, J.M. Yield and Metabolite Production of Pelargonium Sidoides DC. in Response to Irrigation and Nitrogen Management. Metabolites 2020, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Fortuny, A.P.; Bueno, R.A.; Pereira da Costa, J.H.; Zanor, M.I.; Rodríguez, G.R. Tomato Fruit Quality Traits and Metabolite Content Are Affected by Reciprocal Crosses and Heterosis. J. Exp. Bot. 2021, 72, 5407–5425. [Google Scholar] [CrossRef]
- Durán-Soria, S.; Pott, D.M.; Will, F.; Mesa-Marín, J.; Lewandowski, M.; Celejewska, K.; Masny, A.; Żurawicz, E.; Jennings, N.; Sønsteby, A. Exploring Genotype-by-Environment Interactions of Chemical Composition of Raspberry by Using a Metabolomics Approach. Metabolites 2021, 11, 490. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Yang, Z. Biosyntheses of Volatile Phenylpropanoids/Benzenoids in Tea (Camellia Sinensis) and Their Formations in Response to Stresses. J. Trop. Subtrop. Bot. 2019, 27, 591–600. [Google Scholar]
- Fuke, S.; Ueda, Y. Interactions between Umami and Other Flavor Characteristics. Trends Food Sci. Technol. 1996, 7, 407–411. [Google Scholar] [CrossRef]
- Tsakiris, A.; Kallithraka, S.; Kourkoutas, Y. Grape Brandy Production, Composition and Sensory Evaluation. J. Sci. Food Agric. 2014, 94, 404–414. [Google Scholar] [CrossRef]
- Tressl, R.; Grünewald, K.G.; Helak, B. Formation of Flavour Components from Proline and Hydroxyproline with Glucose and Maltose and Their Importance to Food Flavour. In Flavour’81; De Gruyter: Berlin, Germany, 2019; pp. 397–416. ISBN 3-11-084406-0. [Google Scholar]
- Nepi, M.; Soligo, C.; Nocentini, D.; Abate, M.; Guarnieri, M.; Cai, G.; Bini, L.; Puglia, M.; Bianchi, L.; Pacini, E. Amino Acids and Protein Profile in Floral Nectar: Much More than a Simple Reward. Flora-Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 475–481. [Google Scholar] [CrossRef]
- Kang, H.J.; Yang, H.J.; Kim, M.J.; Han, E.-S.; Kim, H.-J.; Kwon, D.Y. Metabolomic Analysis of Meju during Fermentation by Ultra Performance Liquid Chromatography-Quadrupole-Time of Flight Mass Spectrometry (UPLC-Q-TOF MS). Food Chem. 2011, 127, 1056–1064. [Google Scholar] [CrossRef]
- Xiang, Y.; Huang, C.-H.; Hu, Y.; Wen, J.; Li, S.; Yi, T.; Chen, H.; Xiang, J.; Ma, H. Evolution of Rosaceae Fruit Types Based on Nuclear Phylogeny in the Context of Geological Times and Genome Duplication. Mol. Biol. Evol. 2017, 34, 262–281. [Google Scholar] [CrossRef] [Green Version]
- Contreras, C.; González-Agüero, M.; Defilippi, B.G. A Review of Pepino (Solanum Muricatum Aiton) Fruit: A Quality Perspective. HortScience 2016, 51, 1127–1133. [Google Scholar] [CrossRef] [Green Version]
- Piperata, B.A.; Salazar, M.; Schmeer, K.K.; Herrera Rodríguez, A. Tranquility Is a Child with a Full Belly: Pathways Linking Food Insecurity and Maternal Mental Distress in Nicaragua. Ecol. Food Nutr. 2020, 59, 79–103. [Google Scholar] [CrossRef]
- Gill, J.; Park, Y.; McGinnis, J.P.; Perez-Sanchez, C.; Blanchette, M.; Si, K. Regulated Intron Removal Integrates Motivational State and Experience. Cell 2017, 169, 836–848.e15. [Google Scholar] [CrossRef]
- Sun, X.; Jiao, C.; Schwaninger, H.; Chao, C.T.; Ma, Y.; Duan, N.; Khan, A.; Ban, S.; Xu, K.; Cheng, L. Phased Diploid Genome Assemblies and Pan-Genomes Provide Insights into the Genetic History of Apple Domestication. Nat. Genet. 2020, 52, 1423–1432. [Google Scholar] [CrossRef]
- Xiao, Y.; Jiang, S.; Cheng, Q.; Wang, X.; Yan, J.; Zhang, R.; Qiao, F.; Ma, C.; Luo, J.; Li, W. The Genetic Mechanism of Heterosis Utilization in Maize Improvement. Genome Biol. 2021, 22, 148. [Google Scholar] [CrossRef]
- Konermann, S.; Brigham, M.D.; Trevino, A.E.; Joung, J.; Abudayyeh, O.O.; Barcena, C.; Hsu, P.D.; Habib, N.; Gootenberg, J.S.; Nishimasu, H. Genome-Scale Transcriptional Activation by an Engineered CRISPR-Cas9 Complex. Nature 2015, 517, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.; Liu, Y.; Guo, Y.; Duan, W.; Fan, P.; Li, S.; Liang, Z. Optimizing the CRISPR/Cas9 System for Genome Editing in Grape by Using Grape Promoters. Hortic. Res. 2021, 8, 52. [Google Scholar] [CrossRef]
- Xing, S.; Chen, K.; Zhu, H.; Zhang, R.; Zhang, H.; Li, B.; Gao, C. Fine-Tuning Sugar Content in Strawberry. Genome Biol. 2020, 21, 230. [Google Scholar] [CrossRef]
- Dong, L.; Qi, X.; Zhu, J.; Liu, C.; Zhang, X.; Cheng, B.; Mao, L.; Xie, C. Supersweet and Waxy: Meeting the Diverse Demands for Specialty Maize by Genome Editing. Plant Biotechnol. J. 2019, 17, 1853. [Google Scholar] [CrossRef]
- Forner, J.; Kleinschmidt, D.; Meyer, E.H.; Fischer, A.; Morbitzer, R.; Lahaye, T.; Schöttler, M.A.; Bock, R. Targeted Introduction of Heritable Point Mutations into the Plant Mitochondrial Genome. Nat. Plants 2022, 8, 245–256. [Google Scholar] [CrossRef]
- Maher, M.F.; Nasti, R.A.; Vollbrecht, M.; Starker, C.G.; Clark, M.D.; Voytas, D.F. Plant Gene Editing through de Novo Induction of Meristems. Nat. Biotechnol. 2020, 38, 84–89. [Google Scholar] [CrossRef]
- Lian, Z.; Nguyen, C.D.; Liu, L.; Wang, G.; Chen, J.; Wang, S.; Yi, G.; Wilson, S.; Ozias-Akins, P.; Gong, H. Application of Developmental Regulators to Improve in Planta or in Vitro Transformation in Plants. Plant Biotechnol. J. 2022, 20, 1622. [Google Scholar] [CrossRef]
- de Araújo, A.C.A.; Santos, E.G.D.R.; De Sà, K.S.G.; Furtado, V.K.T.; Santos, F.A.; De Lima, R.C.; Krejcová, L.V.; Santos-Lobato, B.L.; Pinto, G.H.L.; Cabral, A.d.S. Hand Resting Tremor Assessment of Healthy and Patients with Parkinson’s Disease: An Exploratory Machine Learning Study. Front. Bioeng. Biotechnol. 2020, 8, 778. [Google Scholar] [CrossRef]
- Munblit, D.; Abrol, P.; Sheth, S.; Chow, L.Y.; Khaleva, E.; Asmanov, A.; Lauriola, S.; Padovani, E.M.; Comberiati, P.; Boner, A.L. Levels of Growth Factors and Iga in the Colostrum of Women from Burundi and Italy. Nutrients 2018, 10, 1216. [Google Scholar] [CrossRef] [Green Version]
- Shukla, P.; Verma, A.; Verma, S.; Kumar, M. Interpreting SVM for Medical Images Using Quadtree. Multimed. Tools Appl. 2020, 79, 29353–29373. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, S.; Ai, Z.; Shen, T.; Zhang, H.; Duan, S.; Chen, Y.-C.; Yin, X.; Sun, J. Development and External Validation of a Stability Machine Learning Model to Identify Wake-up Stroke Onset Time from MRI. Eur. Radiol. 2022, 32, 3661–3669. [Google Scholar] [CrossRef]
- Morse, J.G.; Hoddle, M.S. Invasion Biology of Thrips. Annu. Rev. Entomol. 2006, 51, 67–89. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Wishart, D.S. MSEA: A Web-Based Tool to Identify Biologically Meaningful Patterns in Quantitative Metabolomic Data. Nucleic Acids Res. 2010, 38, W71–W77. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Zhao, W.; Li, Y.; Si, C.; Sun, X.; Zhong, Q.; Yang, S. An Exploration of Pepino (Solanum muricatum) Flavor Compounds Using Machine Learning Combined with Metabolomics and Sensory Evaluation. Foods 2022, 11, 3248. https://doi.org/10.3390/foods11203248
Sun Z, Zhao W, Li Y, Si C, Sun X, Zhong Q, Yang S. An Exploration of Pepino (Solanum muricatum) Flavor Compounds Using Machine Learning Combined with Metabolomics and Sensory Evaluation. Foods. 2022; 11(20):3248. https://doi.org/10.3390/foods11203248
Chicago/Turabian StyleSun, Zhu, Wenwen Zhao, Yaping Li, Cheng Si, Xuemei Sun, Qiwen Zhong, and Shipeng Yang. 2022. "An Exploration of Pepino (Solanum muricatum) Flavor Compounds Using Machine Learning Combined with Metabolomics and Sensory Evaluation" Foods 11, no. 20: 3248. https://doi.org/10.3390/foods11203248