

Screening of a Novel Lactiplantibacillus plantarum MMB-05 and Lacticaseibacillus casei Fermented Sandwich Seaweed Scraps: Chemical Composition, In Vitro Antioxidant, and Volatile Compounds Analysis by GC-IMS
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation, Purification, and Identification of L. Plantarum MMB-05
2.1.1. Determination of Growth Value and pH Value of Strains
2.1.2. Salt Tolerance Test and Antibacterial Activity of L. plantarum MMB-05
2.2. Preparation of P. yezoensis Sauces and Fermentation
2.2.1. Extraction of Supernatant
2.2.2. Sensory Evaluation
2.2.3. Determination of Basic Physicochemical Indexes
2.2.4. Total Phenolic and Total Flavonoids Contents
2.2.5. Determination of Organic acid Content and Antioxidant Activities
2.2.6. Determination of Free Amino Acid Content
2.2.7. Determination of Volatile Substances by GC-IMS
2.3. Statistical Analysis
3. Results and Discussion
3.1. Identification of L. plantarum MMB-05
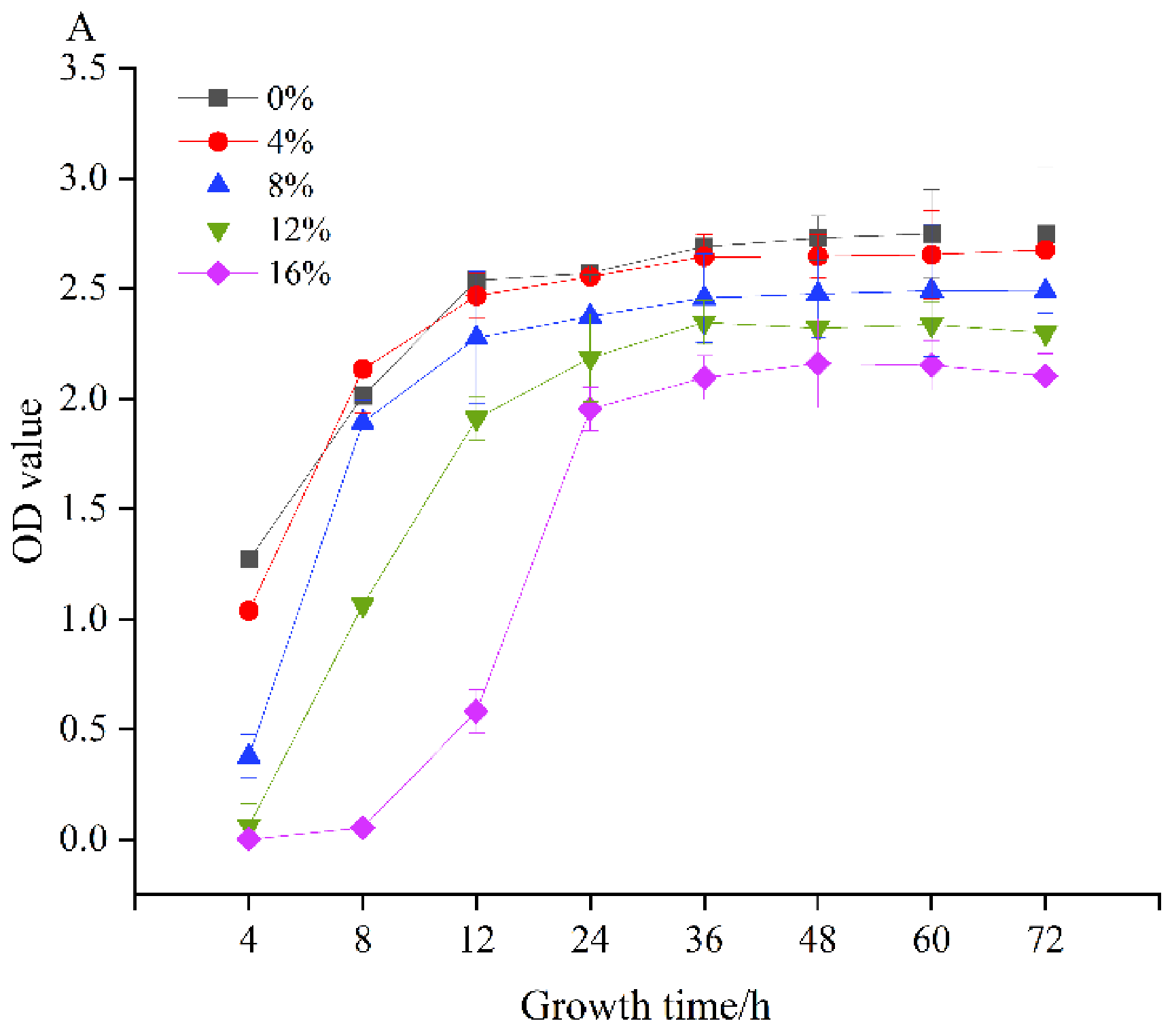
3.2. Growth Characteristics
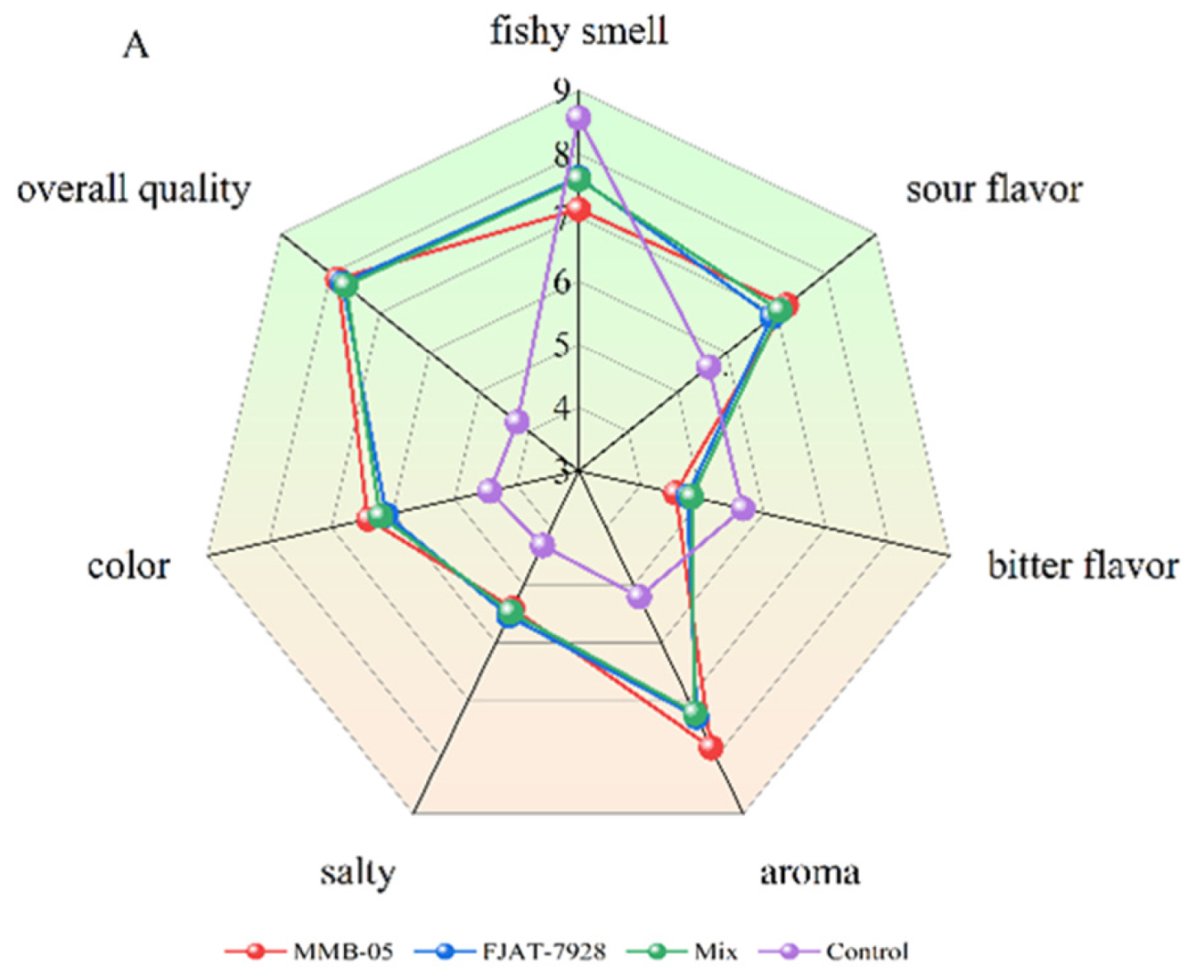
3.3. Sensory Evaluation
3.4. Physicochemical Indexes
3.5. Viable Cell Counts, pH, and Total Sugar
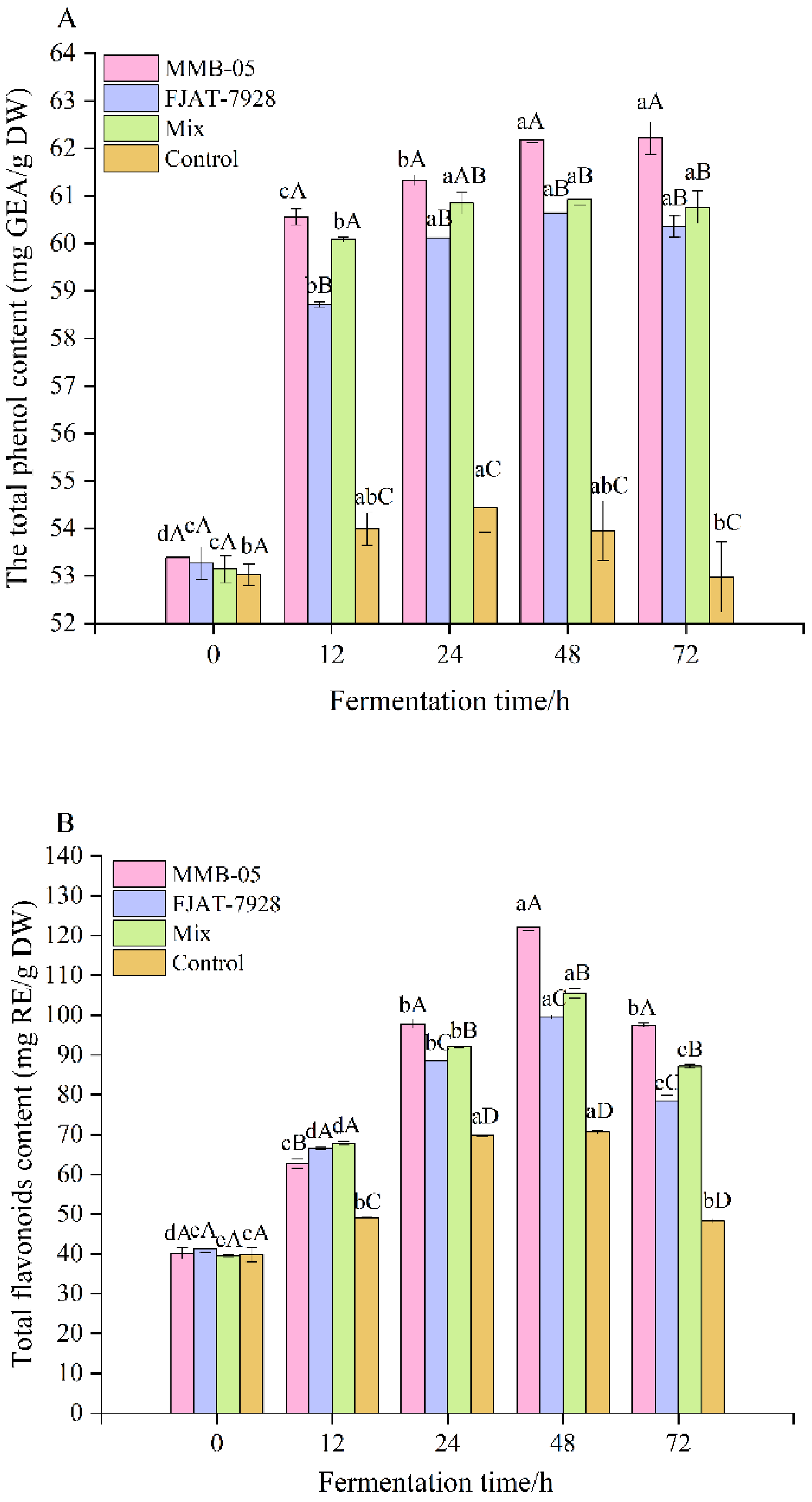
3.6. Total Phenolic, Flavonoids Contents and Organic Acids
3.7. In Vitro Antioxidant Capacity
3.8. Free Amino Acids
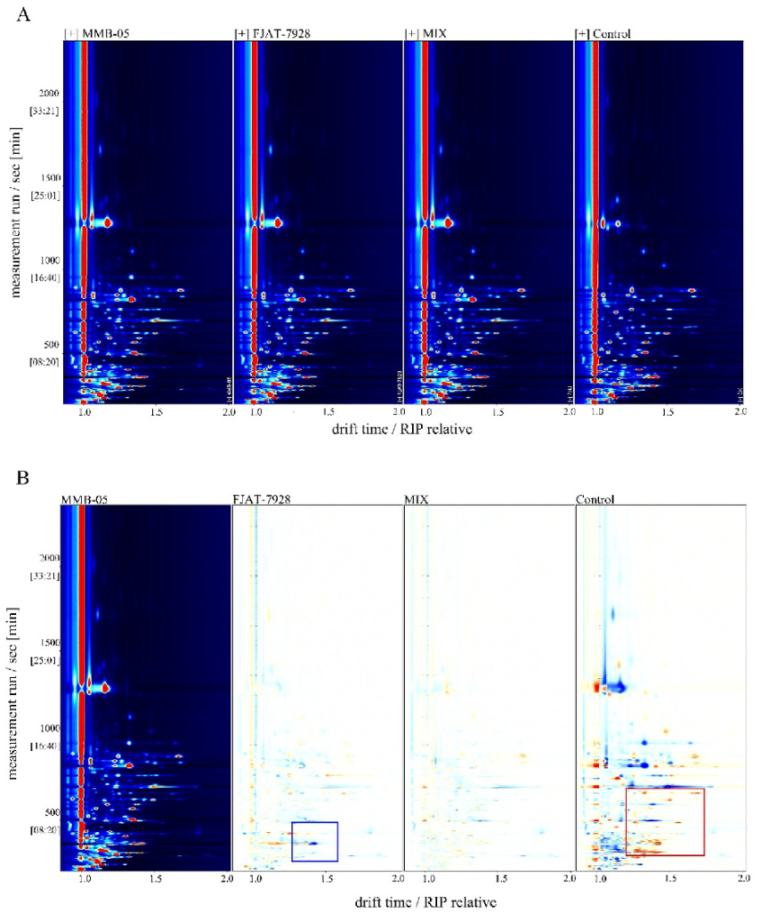
3.9. Volatiles Compounds Analysis
3.9.1. Principal Component Analysis of Volatiles Compounds
3.9.2. Aroma Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aung, T.; Eun, J. Production and characterization of a novel beverage from laver (Porphyra dentata) through fermentation with kombucha consortium. Food Chem. 2021, 350, 129274. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Y.P. The growth performance and nonspecific immunity of juvenile grass carp ( Ctenopharyngodon idella ) affected by dietary Porphyra yezoensis polysaccharide supplementation. Fish. Shellfish Immunol. 2019, 87, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.D.P.; De Lima, P.F.; Santiago-Aguiar, R.S.; Rocha, M.V.P. Evaluation of protic ionic liquids as potential solvents for the heating extraction of phycobiliproteins from Spirulina (Arthrospira) platensis. Algal Res. 2019, 38, 101391. [Google Scholar] [CrossRef]
- Sutherland, J.E.; Lindstrom, S.C.; Nelson, W.A.; Brodie, J.; Lynch, M.D.; Hwang, M.S.; Choi, H.G.; Miyata, M.; Kikuchi, N.; Oliveira, M.C. A new look at an ancient order: Generic revision of the Bangiales (Rhodophyta). J. Phycol. 2011, 47, 1131–1151. [Google Scholar] [CrossRef]
- Zhou, X.H.; Wang, P.; Yan, B.L.; Li, X.S.; Yi, L.F. Characterization and expression patterns of two 70-kDa heat shock protein genes in the intertidal red alga Porphyra yezoensis. Bot. Mar. 2011, 54, 447–455. [Google Scholar] [CrossRef]
- Hadj, S.J.; Levante, A.; Ferrillo, A.; Trapani, F.; Bernini, V.; Galaverna, G.; Neviani, E.; Lazzi, C. Exploring the Potential of Lactic Acid Fermentation for the Recovery of Exhausted Vanilla Beans. Front. Nutr. 2022, 9, 858716. [Google Scholar] [CrossRef]
- Lee, K.H.; Jang, Y.W.; Lee, J.; Kim, S.; Park, C.; Yoo, H.Y. Statistical Optimization of Alkali Pretreatment to Improve Sugars Recovery from Spent Coffee Grounds and Utilization in Lactic Acid Fermentation. Processes 2021, 9, 494. [Google Scholar] [CrossRef]
- Luan, X.X.; Feng, M.Q.; Sun, J. Effect of Lactobacillus plantarum on antioxidant activity in fermented sausage. Food Rev. Int. 2021, 144, 110351. [Google Scholar] [CrossRef]
- Chaudhary, A.; Verma, K.; Saharan, S.B. Probiotic Potential of Blueberry Jam Fermented with Lactic Acid Bacteria. Curr. Res. Nutr. Food Sci. J. 2020, 8, 65–78. [Google Scholar] [CrossRef]
- Tang, V.C.Y.; Sun, J.P.A.; Goh, R.M.V.; Huang, Y.; Ee, K.H.; Lassabliere, B. Biovalorisation of spent Konacha tea leaves via single-culture fermentation involving wine yeasts and lactic acid bacteria. J. Appl. Microbiol. 2022, 133, 1461–1478. [Google Scholar] [CrossRef]
- Wu, H.; Shum, T.F.; Chiou, J. Characterization of the Probiotic Potential of Lactic Acid Bacteria Isolated from Kimchi, Yogurt, and Baby Feces in Hong Kong and Their Performance in Soymilk Fermentation. Microorganisms 2021, 9, 2544. [Google Scholar] [CrossRef]
- Rahmat, F.Z.; Micheal, O.O.; Kamoldeen, A.A.; Kudirat, B.S.; Risikat, N.A.; Tariq, O.A.; Alhassan, S. Optimisation of Lactic Acid Fermentation from Cassava Peel by Lactobacillus casei (ATCC334). Microbiol. Res. J. Int. 2021, 31, 29–42. [Google Scholar]
- Obioha, P.I.; Ouoba, L.I.I.; Anyogu, A.; Awamaria, B.; Atchia, S.; Ojimelikwe, P.C.; Sutherland, J.P.; Ghoddusi, H.B. Identification and characterisation of the lactic acid bacteria associated with the traditional fermentation of dairy fermented product. Braz. J. Microbiol. 2021, 52, 1–13. [Google Scholar] [CrossRef]
- Yang, G.; Cui, X.L.; Liu, S.; Lu, J.; Hou, X.Y.; Meng, W.R.; Zhu, R.J.; Zhang, W.; Zhang, H.Y.; Zheng, W.; et al. Effects of dietary Lactobacillus helveticus on the growth rate, disease resistance and intestinal health of pond loach (Misgurnus anguillicaudatus). Aquaculture 2021, 544, 737038. [Google Scholar] [CrossRef]
- Yang, J.; Lu, J.; Zhu, Q.Z.; Tao, Y.; Guo, C.; Fang, Y.W.; Wang, S.J.; Chen, L.; Koyande, A.; Krishna, S.P. Isolation and characterization of a novel Lactobacillus plantarum MMB-07 from traditional Suanyu for Acanthogobius hasta fermentation. J. Biosci. Bioeng. 2021, 132, 161–166. [Google Scholar] [CrossRef]
- Uchida, M.; Miyoshi, T.; Yoshida, G.; Niwa, K.; Mori, M.; Wakabayashi, H. Isolation and characterization of halophilic lactic acid bacteria acting as a starter culture for sauce fermentation of the red alga Nori (Porphyra yezoensis). J. Appl. Microbiol. 2014, 116, 1506–1520. [Google Scholar] [CrossRef]
- Uchida, M.; Kurushima, H.; Ishihara, K.; Murata, Y.; Touhata, K.; Ishida, N.; Niwa, K.; Araki, T. Characterization of fermented seaweed sauce prepared from nori (Pyropia yezoensis). J. Biosci. Bioeng. 2017, 123, 327–332. [Google Scholar] [CrossRef]
- Puton, J.; Nanienik, J. Ion mobility spectrometry: Current status and application for chemical warfareagents detection-sciencedirect. Trac-Trend. Anal. Chem. 2016, 85, 10–20. [Google Scholar] [CrossRef]
- Gallegos, J.; Arce, C.; Jordano, R.; Arce, L.; Medina, L.M. Target identification of volatile metabolites to allow the differentiation of lactic acid bacteria by gas chromatography-ion mobility spectrometry. Food Chem. 2017, 220, 362–370. [Google Scholar] [CrossRef]
- Gao, C.; Wang, R.; Zhang, F.; Sun, Z.C.; Meng, X.H. The process monitors of probiotic fermented sour cherry juice based on the HS-GC-IMS. Microchem. J. 2022, 180, 107537. [Google Scholar] [CrossRef]
- Chi, C.H.; Lin, Y.; Miao, L.H.; Liu, B.; Ge, X.P. Effects of dietary supplementation of a mixture of ferulic acid and probiotics on the fillet quality of Megalobrama amblycephala fed with oxidized oil. Aquaculture 2022, 549, 737786. [Google Scholar] [CrossRef]
- Yang, J.; Gao, T.Q.; Ge, F.; Sun, H.; Cui, Z.H.; Wei, Z.; Wang, W. Porphyra yezoensis Sauces Fermented with Lactic Acid Bacteria: Fermentation Properties, Flavor Profile, and Evaluation of Antioxidant Capacity in vitro. Front. Nutr. 2022, 8, 810460. [Google Scholar] [CrossRef] [PubMed]
- Du, H.C.; Yang, J.; Lu, X.H.; Lu, Z.X.; Bie, X.M.; Zhao, H.Z.; Zhang, C.; Lu, F.X. Purification, Characterization, and Mode of Action of Plantaricin GZ1-27, a Novel Bacteriocin against Bacillus cereus. J. Agric. Food. Chem. 2018, 66, 18. [Google Scholar] [CrossRef] [PubMed]
- González-Quijano, G.K.; Dorantes-Alvarez, L.; Hernández-Sánchez, H.; Jaramillo-Flores, M.E.; Jesús, P.M.; Vera-Ponce, L.A.; Hernández-Rodríguez, C. Halotolerance and survival kinetics of lactic acid bacteria isolated from jalapeño pepper (Capsicum annuum L.) fermentation. J. Food Sci. 2014, 79, M1545–M1553. [Google Scholar] [CrossRef]
- Venkatraman, K.L.; Mehta, A. Health Benefits and Pharmacological Effects of Porphyra Species. Plant Foods Hum. Nutr. 2019, 74, 10–17. [Google Scholar] [CrossRef]
- Patricia, S. Sensory evaluation of food principles and practices. J. Wine. Res. 2013, 24, 1. [Google Scholar]
- Sowmya, B.H.; Usha, A.D. Quantification of Total Phenolics, Flavonoids and Evaluation of in vitro Free Radical Scavenging Activities in Psidium guajava L. Indian J. Pharm. Sci. 2020, 82, 578–585. [Google Scholar] [CrossRef]
- Kim, D.; Jeong, S.W.; Lee, C.Y. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, J.; Niu, X. Simultaneous determination of five organic acids in beef heart persimmon juice based on HPLC. Chin. Brew. 2022, 41, 232–237. [Google Scholar]
- Adeyeye, E.I. Amino acid composition of three species of Nigerian fish: Clarias anguillaris, Oreochromis niloticus and Cynoglossus senegalensis. Food Chem. 2009, 113, 43–46. [Google Scholar] [CrossRef]
- Natalia, A.M.; Andrés, M.G.; Natividad, J.C.; Rocío, G.D.; Cristina, A.; Lourdes, A. Target vs spectral fingerprint data analysis of Iberian ham samples for avoiding labelling fraud using headspace–gas chromatography–ion mobility spectrometry. Food Chem. 2018, 246, 65–73. [Google Scholar]
- Cai, W.C.; Wang, Y.R.; Wang, W.P.; Shu, N.; Hou, Q.C.; Tang, F.X.; Shan, C.H.; Yang, X.Q.; Guo, Z. Insights into the Aroma Profile of Sauce-Flavor Baijiu by GC-IMS Combined with Multivariate Statistical Analysis. J. Anal. Methods Chem. 2022, 2022, 4614330. [Google Scholar] [CrossRef]
- Lan, H.Y.; Teng, Q.; Yang, H.; Deng, L.L.; Zeng, K.F. Screening of bacteriocin-producing lactic acid bacteria in Chinese homemade pickle and dry-cured meat, and bacteriocin identification by genome sequencing. LWT 2020, 125, 109177. [Google Scholar]
- Nawangsih, E.N.; Kusnawan, P.H.; Djamaludin, M. The Growth Curve and Total Colonies Number of Lactobacillus spp on the Pink Guava (Psidium guajava Linnaeus) Juice Medium. In 12th Annual Scientific Meeting, Medical Faculty, Universitas Jenderal Achmad Yani, International Symposium on “Emergency Preparedness and Disaster Response during COVID 19 Pandemic” (ASMC 2021); Atlantis Press: Paris, France, 2021. [Google Scholar]
- Hiroshi, H.; Osamu, S.; Hiroaki, T. Taxonomic homogeneity of a salt-tolerant lactic acid bacteria isolated from shoyu mash. J. Gen. Appl. Microbiol. 2003, 49, 95–100. [Google Scholar]
- Cheong, E.Y.L.; Sandhu, A.; Jayabalan, J. Isolation of lactic acid bacteria with antifungal activity against the common cheese spoilage mould Penicillium commune and their potential as biopreservatives in cheese. Food Control 2014, 46, 91–97. [Google Scholar] [CrossRef]
- Zangeneh, M.; Khorrami, S.; Khaleghi, M. Bacteriostatic activity and partial characterization of the bacteriocin produced by L. plantarum sp. isolated from traditional sourdough. Food Sci. Nutr. 2020, 8, 6023–6030. [Google Scholar] [CrossRef]
- Leggieri, M.C.; Decontardi, S.; Bertuzzi, T.; Battilani, P. Modeling Growth and Toxin Production of Toxigenic Fungi Signaled in Cheese under Different Temperature and Water Activity Regimes. Toxins 2016, 9, 4. [Google Scholar] [CrossRef]
- Li, X.N. Research on Preparation and Storage Characteristics of Sandwich Nori. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2020. [Google Scholar]
- Jabło ´nska-Ry´s, E.; Skrzypczak, K.; Sławińska, A.; Radzki, W.; Gustaw, W. Lactic acid fermentation of edible mushrooms: Tradition, technology, current state of research: A review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 655–669. [Google Scholar] [CrossRef]
- Rakin, M.; Vukasinovic, M.; Siler-marinkovic, S.; Maksimovic, M. Contribution of lactic acid fermentation to improved nutritive quality vegetable juices enriched with brewer’s yeast autolysate. Food Chem. 2007, 100, 599–602. [Google Scholar] [CrossRef]
- Di Cagno, R.; Filannino, P.; Cavoski, I.; Lanera, A.; Mamdouh, B.M.; Gobbetti, M. Bioprocessing technology to exploit organic palm date (Phoenix dactylifera L. cultivar Siwi) fruit as a functional dietary supplement. J. Funct. Foods 2017, 31, 9–19. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Xiao, L.; Wu, M. Effect of lactic acid fermentation on the phytochemical, volatile profile and sensory attributes of mulberry juice. Int. J. Food Prop. 2017, 56, 305–317. [Google Scholar]
- Curiel, J.A.; Pinto, D.; Marzani, B.; Filannino, P.; Farris, G.A.; Gobbetti, M. Lactic acid fermentation as a tool to enhance the antioxidant properties of Myrtus communis berries. Microb. Cell Fact. 2015, 14, 67. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Wen, J.J.; Hu, J.L.; Nie, Q.X.; Chen, H.H.; Xie, M.Y. Momordica charantia juice with Lactobacillus plantarum fermentation: Chemical composition, antioxidant properties and aroma profile. Food Biosci. 2019, 29, 62–72. [Google Scholar] [CrossRef]
- Tu, C.H.; Tang, S.J.; Azi, F.; Hu, W.X.; Dong, M.S. Use of kombucha consortium to transform soy whey into a novel functional beverage. J. Funct. Foods 2019, 52, 81–89. [Google Scholar] [CrossRef]
- Vitas, J.S.; Cvetanovic, A.D.; Maskovic, P.Z.; Svarc-Gajic, J.V.; Malbasa, R.V. Chemical composition and biological activity of novel types of kombucha beverages with yarrow. J. Funct. Foods. 2018, 44, 95–102. [Google Scholar] [CrossRef]
- Chen, Y.L.; Wang, Y.L.; Chen, J.X.; Tang, H.; Wang, C.H.; Li, Z.J.; Xiao, Y. Bioprocessing of soybeans (Glycine max L.) by solid-state fermentation with Eurotium cristatum YL-1 improves total phenolic content, isoflavone aglycones, and antioxidant activity. RSC Adv. 2020, 10, 16928–16941. [Google Scholar] [CrossRef]
- Ge, X.J.; Tang, N.Y.; Huang, Y.X.; Chen, X.H.; Dong, M.S.; Rui, X.; Li, W. Fermentative and physicochemical properties of fermented milk supplemented with sea buckthorn (Hippophae eleagnaceae L.). LWT 2022, 153, 112484. [Google Scholar] [CrossRef]
- Kato, H.; Rhue, M.R.; Nishimura, T. Role of Free Amino Acids and Peptides in Food Taste; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1989. [Google Scholar]
- Schoenberger, C.; Krottenthaler, M.; Back, W. Sensory and analytical characterization of nonvolatile taste-active compounds in bottom-fermented beers. Technol. Quar. 2002, 39, 210–217. [Google Scholar]
- Liu, Y.; Xu, X.L.; Zhou, G.H. Changes in taste compounds of duck during processing. Food Chem. 2007, 102, 22–26. [Google Scholar] [CrossRef]
- Song, J.; Yan, Y.; Wang, X.; Li, X.; Li, W. Characterization of fatty acids, amino acids and organic acids in three colored quinoas based on untargeted and targeted metabolomics. LWT-Food Sci. Technol. 2020, 140, 110690. [Google Scholar] [CrossRef]
- Juliana, M.; Habtu, S.; Jolien, D.; Joachim, J.S.; Xavier, G.; Ann, D.W.; Katleen, R. Effect of Lactic Acid Fermentation on Volatile Compounds and Sensory Characteristics of Mango (Mangifera indica) Juices. Foods 2022, 11, 383. [Google Scholar]
- Ying, W.; Jiang, Y.T.; Cao, J.X. Study on lipolysis-oxidation and volatile flavour compounds of dry-cured goose with different curing salt content during production. Food Chem. 2016, 190, 33–40. [Google Scholar] [CrossRef]
- Huang, L.; Wu, Z.; Chen, X.Q.; Weng, P.F.; Zhang, X. Characterization of flavour and volatile compounds of fermented squid using electronic nose and HPMS in combination with GC-MS. Int. J. Food Prop. 2018, 21, 760–770. [Google Scholar] [CrossRef]
- Sidira, M.; Kandylis, P.; Kanellaki, M. Effect of immobilized Lactobacillus casei on the evolution of flavor compounds in probiotic dry-fermented sausages during ripening. Meat Sci. 2015, 100, 41–51. [Google Scholar] [CrossRef]
- Niu, Y.W.; Chen, X.M.; Xiao, Z.B.; Ma, N.; Zhu, J.C. Characterization of aroma-active compounds in three Chinese Moutai liquors by gas chromatography-olfactometry, gas chromatography-mass spectrometry and sensory evaluation. Nat. Prod. Res. 2017, 31, 938–944. [Google Scholar] [CrossRef]
- Xiao, Z.B.; Yu, D.; Niu, Y.W.; Chen, F.; Song, S.Q.; Zhu, J.C.; Zhu, G.Y. Characterization of aroma compounds of Chinese famous liquors by gas chromatography-mass spectrometry and flash GC electronic-nose. J. Chromatogr. B 2014, 11, 945–946. [Google Scholar] [CrossRef]
- Noguerol-Pato, P.; González-Álvarez, M.; González-Barreiro, C.; Cancho-Grande, B.; Simal-Gándara, J. Evolution of the aromatic profile in Garnacha Tintorera grapes during raisining and comparison with that of the naturally sweet wine obtained. Food Chem. 2013, 139, 1–4. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Names | Result |
|---|---|
| seven leaf glycosides | + |
| fiber the second pond | + |
| maltose | + |
| mannitol | + |
| salicin | + |
| sorbitol | + |
| sucrose | + |
| raffinose | + |
| Indicators | Fermentation Time/h | MMB-05 | FJAT-7928 | Mix | Control |
|---|---|---|---|---|---|
| Water activity (aw) | 0 | 0.76 ± 0.00 aB | 0.80 ± 0.00 aA | 0.80 ± 0.00 bB | 0.80 ± 0.00 aA |
| 24 | 0.75 ± 0.00 bC | 0.78 ± 0.00 bB | 0.78 ± 0.00 cC | 0.79 ± 0.00 bA | |
| 72 | 0.74 ± 0.00 bD | 0.75 ± 0.00 cC | 0.75 ± 0.00 aB | 0.81 ± 0.00 aA | |
| Chromatic aberration (L*) | 0 | 23.87 ± 0.32 bC | 28.39 ± 0.08 bA | 28.53 ± 0.12 bA | 27.44 ± 0.17 bB |
| 24 | 25.94 ± 0.84 aC | 30.92 ± 0.25 aA | 29.64 ± 0.43 aAB | 28.67 ± 0.13 aB | |
| 72 | 25.78 ± 0.04 aB | 27.48 ± 0.58 bA | 26.42 ± 0.39 cB | 28.30 ± 0.15 aA | |
| Viscosity (unit: cp) | 0 | 142.18 ± 1.91 aC | 162.73 ± 2.40 aB | 158.42 ± 0.67 aB | 683.48 ± 0.78 aA |
| 24 | 74.45 ± 1.09 bD | 154.52 ± 1.08 bB | 110.59 ± 0.35 bC | 664.10 ± 2.36 bA | |
| 72 | 67.05 ± 1.68 cC | 113.28 ± 0.67 cB | 67.02 ± 1.62 cC | 494.69 ± 0.57 cA |
| Groups | Taste | Fermentation Time/h | MMB-05 | FJAT-7928 | Mix | Control |
|---|---|---|---|---|---|---|
| Asp | sweet; umami | 24 | 0.16 ± 0.00 cC | 0.24 ± 0.00 cB | 0.14 ± 0.00 cD | 0.50 ± 0.01 aA |
| 48 | 0.18 ± 0.00 bD | 0.34 ± 0.00 bB | 0.18 ± 0.00 bC | 0.36 ± 0.00 bA | ||
| 72 | 0.22 ± 0.00 aC | 0.52 ± 0.00 aA | 0.21 ± 0.00 aD | 0.22 ± 0.00 cB | ||
| Thr | sweet | 24 | 0.16 ± 0.00 cA | 0.05 ± 0.01 cC | 0.15 ± 0.00 cB | 0.05 ± 0.01 cC |
| 48 | 0.18 ± 0.00 bA | 0.08 ± 0.00 bC | 0.15 ± 0.00 bB | 0.07 ± 0.00 bD | ||
| 72 | 0.23 ± 0.00 aA | 0.12 ± 0.00 aC | 0.22 ± 0.00 aB | 0.09 ± 0.00 aD | ||
| Ser | sweet | 24 | ND | 0.06 ± 0.00 cA | ND | 0.06 ± 0.00 aA |
| 48 | ND | 0.10 ± 0.00 bA | ND | 0.03 ± 0.00 bB | ||
| 72 | ND | 0.12 ± 0.00 aA | ND | 0.03 ± 0.00 bB | ||
| Glu | umami; sour | 24 | 2.62 ± 0.00 cA | 2.22 ± 0.00 cC | 2.27 ± 0.00 cB | 2.21 ± 0.02 aC |
| 48 | 2.76 ± 0.00 bB | 2.71 ± 0.02 bC | 3.08 ± 0.00 bA | 2.07 ± 0.00 bD | ||
| 72 | 3.67 ± 0.01 aB | 3.86 ± 0.00 aA | 3.47 ± 0.00 aC | 1.76 ± 0.00 cD | ||
| Gly | sweet | 24 | 0.08 ± 0.00 cC | 0.08 ± 0.00 cD | 0.10 ± 0.00 bB | 0.10 ± 0.00 aA |
| 48 | 0.11 ± 0.00 bA | 0.10 ± 0.00 bB | 0.10 ± 0.00 bB | 0.07 ± 0.00 cC | ||
| 72 | 0.15 ± 0.00 aB | 0.15 ± 0.00 aB | 0.15 ± 0.00 aA | 0.07 ± 0.00 bC | ||
| Ala | sweet | 24 | 3.75 ± 0.00 cA | 2.68 ± 0.03 cC | 3.30 ± 0.00 cB | 2.68 ± 0.03 bC |
| 48 | 3.90 ± 0.00 bB | 3.68 ± 0.03 bC | 3.97 ± 0.00 bA | 2.64 ± 0.00 bD | ||
| 72 | 5.08 ± 0.01 aA | 4.34 ± 0.01 aC | 5.00 ± 0.00 aB | 2.85 ± 0.00 aD | ||
| Cys | 24 | 0.03 ± 0.00 cC | 0.04 ± 0.00 cB | 0.03 ± 0.00 cD | 0.09 ± 0.00 aA | |
| 48 | 0.04 ± 0.00 aC | 0.09 ± 0.00 bA | 0.03 ± 0.00 bD | 0.08 ± 0.00 aB | ||
| 72 | 0.04 ± 0.00 bD | 0.19 ± 0.01 aA | 0.06 ± 0.00 aC | 0.08 ± 0.00 bB | ||
| Val | sweet; bitter | 24 | 0.01 ± 0.00 bC | 0.09 ± 0.00 cB | 0.01 ± 0.00 bC | 0.11 ± 0.00 aA |
| 48 | 0.02 ± 0.00 aB | 0.11 ± 0.00 bA | 0.01 ± 0.00 bC | 0.11 ± 0.00 aA | ||
| 72 | 0.02 ± 0.00 aC | 0.23 ± 0.00 aA | 0.02 ± 0.01 aC | 0.11 ± 0.00 aB | ||
| Met | bitter | 24 | ND | ND | ND | ND |
| 48 | 0.01 ± 0.00 abB | 0.01 ± 0.00 bAB | 0.01 ± 0.00 aB | 0.01 ± 0.00 bA | ||
| 72 | 0.01 ± 0.00 aC | 0.02 ± 0.00 aB | 0.01 ± 0.00 aC | 0.08 ± 0.00 aA | ||
| Ile | bitter | 24 | 0.02 ± 0.00 cB | 0.06 ± 0.00 cA | 0.02 ± 0.00 bB | 0.06 ± 0.00 aA |
| 48 | 0.03 ± 0.00 bC | 0.08 ± 0.00 bA | 0.03 ± 0.00 aB | 0.03 ± 0.00 bB | ||
| 72 | 0.03 ± 0.00 aB | 0.09 ± 0.00 aA | 0.03 ± 0.00 aB | 0.03 ± 0.00 cB | ||
| Leu | bitter | 24 | 0.02 ± 0.00 cD | 0.10 ± 0.00 bB | 0.04 ± 0.00 bC | 0.16 ± 0.00 cA |
| 48 | 0.05 ± 0.00 bD | 0.16 ± 0.00 aB | 0.06 ± 0.00 aC | 0.16 ± 0.00 bA | ||
| 72 | 0.06 ± 0.00 aB | 0.05 ± 0.00 cC | 0.06 ± 0.00 aB | 0.19 ± 0.00 aA | ||
| Tyr | bitter | 24 | 0.35 ± 0.00 cA | 0.36 ± 0.00 cA | 0.31 ± 0.01 cB | 0.03 ± 0.00 cC |
| 48 | 0.37 ± 0.00 bC | 0.43 ± 0.00 bB | 0.45 ± 0.00 bA | 0.13 ± 0.00 bD | ||
| 72 | 0.47 ± 0.00 aB | 0.49 ± 0.00 aA | 0.47 ± 0.00 aC | 0.27 ± 0.00 aD | ||
| Phe | sweet; bitter | 24 | 0.06 ± 0.00 cB | 0.04 ± 0.00cC | 0.20 ± 0.00 cA | 0.04 ± 0.00 cC |
| 48 | 0.07 ± 0.00 bC | 0.06 ± 0.00 bD | 0.27 ± 0.00 bA | 0.14 ± 0.00 bB | ||
| 72 | 0.08 ± 0.00 aC | 0.07 ± 0.00 aD | 0.30 ± 0.00 aA | 0.30 ± 0.00 aB | ||
| Lys | bitter | 24 | 0.01 ± 0.00 cB | 0.01 ± 0.00 cB | 0.01 ± 0.00 cA | 0.01 ± 0.00 cB |
| 48 | 0.02 ± 0.00 bA | 0.02 ± 0.00 bA | 0.02 ± 0.00 bB | 0.02 ± 0.00 bB | ||
| 72 | 0.03 ± 0.00 aC | 0.08 ± 0.00 aA | 0.03 ± 0.00 aC | 0.05 ± 0.00 aB | ||
| His | sour; bitter | 24 | 1.37 ± 0.00 cB | 1.28 ± 0.00 cC | 1.42 ± 0.00 cA | 0.03 ± 0.00 bD |
| 48 | 1.80 ± 0.00 aA | 1.65 ± 0.00 bC | 1.70 ± 0.00 bB | 0.03 ± 0.00 abD | ||
| 72 | 1.75 ± 0.00 bC | 2.01 ± 0.00 aA | 2.00 ± 0.01 aB | 0.04 ± 0.00 aD | ||
| Arg | bitter | 24 | 0.17 ± 0.00 cA | 0.05 ± 0.00 cB | ND | 0.02 ± 0.00 cC |
| 48 | 0.17 ± 0.00 bA | 0.07 ± 0.00 bB | ND | 0.02 ± 0.00 bC | ||
| 72 | 0.22 ± 0.00 aA | 0.18 ± 0.00 aB | 0.01 ± 0.00 aD | 0.02 ± 0.00 aC | ||
| Pro | sweet | 24 | 0.28 ± 0.01 cA | 0.13 ± 0.00 cC | 0.23 ± 0.00 cB | ND |
| 48 | 0.31 ± 0.00 bA | 0.18 ± 0.00 bC | 0.30 ± 0.01 bB | ND | ||
| 72 | 0.36 ± 0.00 aA | 0.26 ± 0.00 aB | 0.34 ± 0.01 aA | ND |
| Compounds | CAS# | RI | Rt | Dt | Relative Amount (%) | |||
|---|---|---|---|---|---|---|---|---|
| MM B-05 | FJAT-7928 | Mix | Control | |||||
| Aldehydes | ||||||||
| Phenylacetaldehyde | C122781 | 1763.10 | 2249.53 | 1.26 | 0.18 ± 0.03 A | 0.17 ± 0.01 A | 0.15 ± 0.01 A | 0.22 ± 0.02 A |
| 2-Decenal | C3913711 | 1766.10 | 2264.39 | 1.49 | 0.15 ± 0.01 A | 0.16 ± 0.01 A | 0.17 ± 0.03 A | 0.20 ± 0.01 A |
| (E,E)-2,4-Octadienal | C30361285 | 1665.00 | 1818.42 | 1.28 | 0.21 ± 0.01 A | 0.22 ± 0.01 A | 0.21 ± 0.01 A | 0.29 ± 0.02 A |
| Benzaldehyde | C100527 | 1549.30 | 1414.40 | 1.16 | 0.45 ± 0.01 C | 0.46 ± 0.03 C | 0.48 ± 0.02 B | 1.08 ± 0.02 A |
| (E)-2-Octenal-M | C2548870 | 1436.10 | 1106.41 | 1.34 | 0.56 ± 0.01 AB | 0.54 ± 0.12 B | 0.73 ± 0.10 A | 0.90 ± 0.14 A |
| (E)-2-Octenal-D | C2548870 | 1437.50 | 1109.70 | 1.82 | 0.07 ± 0.01 AB | 0.06 ± 0.01 B | 0.09 ± 0.02 A | 0.11 ± 0.01 A |
| Nonanal | C124196 | 1400.90 | 1025.03 | 1.49 | 0.20 ± 0.01 B | 0.14 ± 0.01 C | 0.20 ± 0.01 B | 0.58 ± 0.03 A |
| (E)-2-Heptenal-M | C18829555 | 1328.90 | 876.80 | 1.26 | 1.25 ± 0.03 C | 1.32 ± 0.14 C | 1.41 ± 0.04 B | 2.01 ± 0.10 A |
| (E)-2-Heptenal-D | C18829555 | 1329.70 | 878.28 | 1.67 | 1.62 ± 0.06 B | 1.40 ± 0.45 B | 2.43 ± 0.44 A | 3.23 ± 0.55 A |
| (E)-2-Hexenal-M | C6728263 | 1230.20 | 712.37 | 1.18 | 0.46 ± 0.02 C | 0.47 ± 0.04 C | 0.55 ± 0.02 B | 0.91 ± 0.03 A |
| 3-Methyl-2-butenal | C107868 | 1211.60 | 685.43 | 1.09 | 0.20 ± 0.00 C | 0.23 ± 0.00 B | 0.23 ± 0.00 A | 0.14 ± 0.01 D |
| Cyclopentanone | C120923 | 1196.30 | 664.06 | 1.11 | 0.17 ± 0.00 B | 0.24 ± 0.01 A | 0.17 ± 0.00 B | 0.16 ± 0.00 C |
| Heptanal-D | C111717 | 1194.30 | 661.28 | 1.69 | 0.02 ± 0.00 BC | 0.01 ± 0.00 C | 0.03 ± 0.00 B | 0.21 ± 0.02 A |
| 2-Heptanone-M | C110430 | 1191.70 | 657.56 | 1.26 | 0.35 ± 0.01 B | 0.49 ± 0.01 A | 0.35 ± 0.01 B | 0.36 ± 0.01 C |
| 2-Methyl-2-pentenal-M | C623369 | 1172.40 | 616.69 | 1.16 | 0.57 ± 0.00 A | 0.60 ± 0.00 A | 0.57 ± 0.02 A | 0.72 ± 0.00 A |
| 2-Methyl-2-pentenal-D | C623369 | 1172.90 | 617.61 | 1.50 | 0.93 ± 0.03 B | 0.81 ± 0.04 C | 0.80 ± 0.02 C | 1.69 ± 0.03 A |
| (E)-2-Pentenal-M | C1576870 | 1148.30 | 569.31 | 1.11 | 0.76 ± 0.01 C | 0.81 ± 0.03 BC | 0.82 ± 0.01 A | 1.00 ± 0.01 AB |
| (E)-2-Pentenal-D | C1576870 | 1148.30 | 569.31 | 1.36 | 1.30 ± 0.03 BC | 1.18 ± 0.16 C | 1.43 ± 0.09 AB | 1.90 ± 0.11 A |
| (Z)-2-Pentenal-M | C1576869 | 1114.50 | 508.92 | 1.09 | 0.65 ± 0.00 C | 0.73 ± 0.03 B | 0.65 ± 0.03 BC | 1.19 ± 0.02 A |
| (Z)-2-Pentenal-D | C1576869 | 1113.40 | 507.07 | 1.35 | 2.90 ± 0.01 A | 2.58 ± 0.05 C | 2.59 ± 0.02 B | 3.20 ± 0.01 B |
| Hexanal-M | C66251 | 1098.70 | 482.91 | 1.28 | 0.46 ± 0.03 B | 0.19 ± 0.03 C | 0.55 ± 0.03 B | 1.04 ± 0.10 A |
| Hexanal-D | C66251 | 1098.70 | 482.91 | 1.56 | 0.07 ± 0.01 BC | 0.02 ± 0.00 C | 0.11 ± 0.01 B | 0.39 ± 0.11 A |
| Pentanal | C110623 | 1001.10 | 361.32 | 1.43 | 1.40 ± 0.10 C | 0.22 ± 0.06 D | 1.54 ± 0.08 B | 3.00 ± 0.06 A |
| 3-Methylbutanal | C590863 | 928.20 | 305.51 | 1.40 | 1.14 ± 0.05 B | 0.84 ± 0.10 C | 1.07 ± 0.03 B | 2.90 ± 0.02 A |
| Butanal | C123728 | 891.90 | 281.52 | 1.28 | 0.39 ± 0.01 B | 0.33 ± 0.06 C | 0.44 ± 0.02 B | 1.03 ± 0.03 A |
| Diethyl acetal | C105577 | 902.70 | 288.44 | 1.03 | 0.60 ± 0.02 B | 0.66 ± 0.03 B | 0.63 ± 0.03 B | 0.90 ± 0.01 A |
| Acrolein | C107028 | 862.60 | 263.53 | 1.06 | 0.44 ± 0.01 B | 0.47 ± 0.01 B | 0.47 ± 0.02 A | 0.33 ± 0.02 C |
| Propanal | C123386 | 812.10 | 235.20 | 1.14 | 2.97 ± 0.03 B | 3.23 ± 0.04 A | 2.85 ± 0.02 C | 3.76 ± 0.03 B |
| Acetaldehyde | C75070 | 756.90 | 207.69 | 1.02 | 0.82 ± 0.05 A | 0.58 ± 0.04 B | 0.79 ± 0.09 A | 0.57 ± 0.03 B |
| 2-Methylpropanal | C78842 | 828.40 | 243.99 | 1.28 | 0.01 ± 0.00 BC | 0.01 ± 0.00 C | 0.02 ± 0.00 B | 0.08 ± 0.00 A |
| Octanal-M | C124130 | 1294.20 | 813.28 | 1.41 | 0.09 ± 0.00 B | 0.10 ± 0.01 B | 0.10 ± 0.00 B | 0.47 ± 0.26 A |
| Octanal-D | C124130 | 1294.30 | 813.41 | 1.83 | 0.02 ± 0.00 B | 0.02 ± 0.00 B | 0.02 ± 0.00 B | 0.07 ± 0.04 A |
| (E)-2-Hexenal-D | C6728263 | 1230.10 | 712.13 | 1.52 | 0.32 ± 0.00 A | 0.25 ± 0.05 B | 0.33 ± 0.02 A | 0.44 ± 0.04 A |
| Methacrolein | C78853 | 891.30 | 281.10 | 1.22 | 0.26 ± 0.01 B | 0.24 ± 0.03 BC | 0.33 ± 0.01 A | 0.26 ± 0.02 C |
| Ketones | ||||||||
| 2-Cyclohexen-1-one | C930687 | 1423.60 | 1076.91 | 1.10 | 0.03 ± 0.00 A | 0.03 ± 0.00 A | 0.03 ± 0.00 A | 0.02 ± 0.00 B |
| 6-Methyl-5-hepten-2-one | C110930 | 1345.00 | 907.92 | 1.18 | 0.21 ± 0.01 B | 0.17 ± 0.01 C | 0.24 ± 0.02 A | 0.16 ± 0.01 D |
| 1-Hydroxy-2-propanone-M | C116096 | 1314.10 | 849.09 | 1.07 | 1.87 ± 0.02 A | 1.93 ± 0.04 A | 1.90 ± 0.03 A | 1.92 ± 0.25 B |
| 1-Hydroxy-2-propanone-D | C116096 | 1314.80 | 850.42 | 1.23 | 0.94 ± 0.07 A | 0.98 ± 0.02 A | 0.93 ± 0.02 A | 1.04 ± 0.07 B |
| 3-Hydroxy-2-butanone | C513860 | 1298.70 | 821.10 | 1.33 | 6.05 ± 0.17 A | 5.84 ± 0.09A | 6.00 ± 0.10 A | 1.68 ± 1.74 B |
| 1-Octen-3-one-M | C4312996 | 1312.70 | 846.42 | 1.28 | 0.88 ± 0.03 A | 0.79 ± 0.01 B | 0.87 ± 0.02 A | 0.85 ± 0.11 B |
| 1-Octen-3-one-D | C4312996 | 1308.30 | 838.43 | 1.68 | 0.29 ± 0.01 AB | 0.24 ± 0.11 B | 0.43 ± 0.11 A | 0.56 ± 0.17 A |
| Cyclohexanone | C108941 | 1311.90 | 845.09 | 1.46 | 0.27 ± 0.03 B | 0.3 ± 0.06 B | 0.34 ± 0.03 AB | 0.49 ± 0.04 A |
| Heptanal-M | C111717 | 1195.00 | 662.20 | 1.34 | 0.21 ± 0.01 C | 0.13 ± 0.01 D | 0.23 ± 0.01 B | 0.60 ± 0.02 A |
| 2-Heptanone-D | C110430 | 1191.30 | 656.63 | 1.63 | 0.18 ± 0.01 B | 0.28 ± 0.03 A | 0.16 ± 0.01 B | 0.22 ± 0.01 B |
| 2,3-Pentanedione | C600146 | 1061.30 | 431.90 | 1.20 | 0.62 ± 0.05 BC | 0.53 ± 0.10 C | 0.72 ± 0.07 AB | 1.03 ± 0.10 A |
| 1-Penten-3-one-M | C1629589 | 1038.90 | 404.22 | 1.08 | 0.46 ± 0.01 B | 0.51 ± 0.00 A | 0.46 ± 0.01 B | 0.63 ± 0.00 A |
| 1-Penten-3-one-D | C1629589 | 1038.50 | 403.76 | 1.31 | 1.49 ± 0.04 AB | 1.42 ± 0.20 B | 1.66 ± 0.10 A | 1.93 ± 0.10 AB |
| 2-Pentanone | C107879 | 997.20 | 357.17 | 1.36 | 1.44 ± 0.01 B | 1.80 ± 0.10 A | 1.49 ± 0.03 B | 1.51 ± 0.02 C |
| 2-Butanone | C78933 | 914.60 | 296.28 | 1.24 | 3.48 ± 0.04 A | 3.67 ± 0.07 A | 3.47 ± 0.01 A | 4.32 ± 0.04 A |
| Acetone | C67641 | 835.90 | 248.15 | 1.11 | 5.42 ± 0.18 AB | 6.09 ± 0.38 A | 5.27 ± 0.07 B | 7.13 ± 0.04 AB |
| Alcohols | ||||||||
| 1-Octanol | C111875 | 1652.70 | 1770.52 | 1.48 | 0.18 ± 0.04 A | 0.18 ± 0.01 A | 0.18 ± 0.02 A | 0.19 ± 0.00 A |
| Linalool | C78706 | 1633.00 | 1696.19 | 1.22 | 0.15 ± 0.02 A | 0.17 ± 0.01 A | 0.15 ± 0.01 A | 0.14 ± 0.01 B |
| 1-Octen-3-ol | C3391864 | 1483.40 | 1225.97 | 1.17 | 0.14 ± 0.01 BC | 0.13 ± 0.02 C | 0.18 ± 0.03 AB | 0.26 ± 0.04 A |
| 1-Heptanol | C111706 | 1486.60 | 1234.50 | 1.41 | 0.07 ± 0.00 B | 0.08 ± 0.00 A | 0.08 ± 0.00 AB | 0.06 ± 0.00 C |
| (Z)-Hex-3-enol | C928961 | 1418.70 | 1065.36 | 1.25 | 0.11 ± 0.01 A | 0.09 ± 0.01 B | 0.12 ± 0.01 A | 0.05 ± 0.01 C |
| 1-Hexanol-M | C111273 | 1369.20 | 956.84 | 1.33 | 1.21 ± 0.01 B | 1.39 ± 0.05 A | 1.12 ± 0.02 C | 0.19 ± 0.03 D |
| 1-Hexanol-D | C111273 | 1368.40 | 955.36 | 1.64 | 0.22 ± 0.00 B | 0.28 ± 0.02 A | 0.20 ± 0.00 C | 0.04 ± 0.01 D |
| (Z)-2-Penten-1-ol | C1576950 | 1337.40 | 893.10 | 0.95 | 0.59 ± 0.00 A | 0.52 ± 0.02 B | 0.57 ± 0.01 A | 0.50 ± 0.00 C |
| 1-Pentanol-M | C71410 | 1264.70 | 765.11 | 1.26 | 0.98 ± 0.02 B | 1.24 ± 0.04A | 0.98 ± 0.02 B | 1.51 ± 0.04 A |
| 1-Pentanol-D | C71410 | 1263.90 | 763.78 | 1.51 | 0.37 ± 0.02 B | 0.62 ± 0.06 A | 0.39 ± 0.03 B | 0.77 ± 0.06 A |
| 3-Methyl-3-buten-1-ol-M | C763326 | 1261.30 | 759.75 | 1.17 | 0.35 ± 0.01 A | 0.23 ± 0.03 B | 0.33 ± 0.01 A | 0.07 ± 0.00 C |
| 3-Methyl-3-buten-1-ol-D | C763326 | 1261.30 | 759.75 | 1.44 | 0.34 ± 0.01 A | 0.27 ± 0.01 B | 0.35 ± 0.01 A | 0.12 ± 0.00 C |
| 3-Methyl-1-butanol-M | C123513 | 1220.10 | 697.51 | 1.24 | 1.19 ± 0.01 A | 1.29 ± 0.02 A | 1.21 ± 0.00 A | 0.40 ± 0.07 B |
| 3-Methyl-1-butanol-D | C123513 | 1219.40 | 696.58 | 1.49 | 3.43 ± 0.03 A | 3.04 ± 0.01 B | 2.71 ± 0.01 C | 0.15 ± 0.02 D |
| 1-Penten-3-ol-M | C616251 | 1176.50 | 625.05 | 0.95 | 0.82 ± 0.01 B | 0.86 ± 0.00 B | 0.86 ± 0.02 A | 0.98 ± 0.01 C |
| 1-Penten-3-ol-D | C616251 | 1175.10 | 622.26 | 1.37 | 0.58 ± 0.01 B | 0.55 ± 0.01 C | 0.53 ± 0.00 C | 0.80 ± 0.03 A |
| 1-Butanol-M | C71363 | 1160.80 | 593.46 | 1.18 | 1.02 ± 0.01 B | 1.12 ± 0.03 A | 1.01 ± 0.01 B | 0.81 ± 0.01 C |
| 1-Butanol-D | C71363 | 1160.80 | 593.46 | 1.38 | 0.75 ± 0.01 B | 0.87 ± 0.01 A | 0.75 ± 0.01 B | 0.27 ± 0.01 C |
| 2-Methyl-1-propanol-M | C78831 | 1105.60 | 494.06 | 1.17 | 0.55 ± 0.01 B | 0.64 ± 0.01 A | 0.60 ± 0.01 A | 0.27 ± 0.01 C |
| 2-Methyl-1-propanol-D | C78831 | 1107.90 | 497.78 | 1.37 | 1.00 ± 0.02 A | 0.93 ± 0.03 B | 0.79 ± 0.02 C | 0.13 ± 0.02 D |
| 1-Propanol-M | C71238 | 1051.40 | 419.45 | 1.11 | 0.49 ± 0.01 B | 0.80 ± 0.04A | 0.45 ± 0.01 B | 0.38 ± 0.02 C |
| 1-Propanol-D | C71238 | 1052.90 | 421.29 | 1.25 | 0.27 ± 0.00 B | 0.76 ± 0.02 A | 0.26 ± 0.01 B | 0.15 ± 0.01 C |
| 2-Butanol | C78922 | 1034.70 | 399.15 | 1.15 | 0.15 ± 0.00 A | 0.15 ± 0.00 B | 0.14 ± 0.00 C | 0.10 ± 0.00 D |
| Ethanol | C64175 | 941.40 | 314.73 | 1.13 | 4.62 ± 0.04 AB | 4.83 ± 0.07 A | 4.49 ± 0.04 B | 3.42 ± 0.03 C |
| Esters | ||||||||
| γ-Butyrolactone | C96480 | 1709.70 | 2003.42 | 1.09 | 0.75 ± 0.03 A | 0.69 ± 0.02 B | 0.65 ± 0.06 B | 0.78 ± 0.03 B |
| Ethyl pyruvate | C617356 | 1249.40 | 741.17 | 1.15 | 0.24 ± 0.00 B | 0.23 ± 0.02 B | 0.27 ± 0.01 B | 1.68 ± 0.07 A |
| Ethyl hexanoate | C123660 | 1244.50 | 733.73 | 1.34 | 0.17 ± 0.01 B | 0.19 ± 0.01 B | 0.18 ± 0.01 AB | 0.25 ± 0.00 A |
| Ethyl butanoate-M | C105544 | 1049.50 | 417.14 | 1.20 | 0.42 ± 0.01 A | 0.32 ± 0.01 C | 0.37 ± 0.00 B | 0.44 ± 0.02 B |
| Ethyl butanoate-D | C105544 | 1049.20 | 416.68 | 1.56 | 0.09 ± 0.01 B | 0.08 ± 0.02 BC | 0.07 ± 0.01 C | 0.17 ± 0.01 A |
| Ethyl Acetate | C141786 | 894.10 | 282.91 | 1.34 | 0.47 ± 0.01 A | 0.39 ± 0.01 B | 0.38 ± 0.01 B | 0.38 ± 0.01 C |
| Methyl acetate | C79209 | 878.50 | 273.15 | 1.19 | 0.34 ± 0.01 A | 0.31 ± 0.04 B | 0.34 ± 0.02 A | 0.17 ± 0.01 C |
| Ethyl isobutyrate | C97621 | 963.80 | 331.00 | 1.19 | 0.36 ± 0.01 B | 0.3 ± 0.03 C | 0.37 ± 0.01 B | 0.65 ± 0.02 A |
| Acids | ||||||||
| 2-Methylpropanoic acid | C79312 | 1690.30 | 1920.83 | 1.17 | 0.15 ± 0.01 A | 0.16 ± 0.01 A | 0.15 ± 0.00 A | 0.09 ± 0.02 B |
| Propanoic acid | C79094 | 1637.00 | 1711.06 | 1.12 | 1.20 ± 0.11 B | 1.62 ± 0.04 A | 1.31 ± 0.02 B | 0.5 ± 0.11C |
| Acetic acid-M | C64197 | 1500.50 | 1272.51 | 1.06 | 9.55 ± 0.14 A | 9.76 ± 0.15 A | 9.13 ± 0.15 A | 7.93 ± 1.23 B |
| Acetic acid-D | C64197 | 1502.20 | 1277.05 | 1.15 | 10.63 ± 0.26 A | 10.54 ± 0.03 A | 11.29 ± 0.59 A | 4.53 ± 2.05 B |
| Furans | ||||||||
| Dihydro-5-methyl-2(3H)-furanone | C108292 | 1735.90 | 2120.69 | 1.13 | 0.18 ± 0.02 A | 0.17 ± 0.02 AB | 0.16 ± 0.01 AB | 0.19 ± 0.01 B |
| 2-Acetylfuran | C1192627 | 1539.20 | 1383.76 | 1.13 | 0.33 ± 0.00 A | 0.26 ± 0.01 C | 0.25 ± 0.00 C | 0.36 ± 0.01 B |
| 5-Methyl-2(3H)-furanone | C591128 | 1442.90 | 1122.86 | 1.13 | 0.05 ± 0.00 A | 0.05 ± 0.00 B | 0.05 ± 0.00 A | 0.04 ± 0.00 C |
| 2-Pentylfuran | C3777693 | 1239.00 | 725.37 | 1.25 | 0.23 ± 0.02 A | 0.21 ± 0.02 A | 0.16 ± 0.02 B | 0.07 ± 0.01 C |
| 2-Ethylfuran | C3208160 | 968.60 | 334.57 | 1.04 | 0.89 ± 0.04 B | 1.02 ± 0.04 A | 0.82 ± 0.03 B | 0.77 ± 0.03 C |
| Furfural | C98011 | 1491.90 | 1248.75 | 1.09 | 0.74 ± 0.02 A | 0.74 ± 0.01 A | 0.66 ± 0.01 B | 0.91 ± 0.04 A |
| Pyrazines | ||||||||
| 2,6-Dimethyl-3-ethylpyrazine | C13925070 | 1557.00 | 1438.52 | 1.24 | 0.14 ± 0.01 A | 0.17 ± 0.01 A | 0.14 ± 0.00 A | 0.11 ± 0.02 B |
| 2-Ethyl-5-methylpyrazine | C13360640 | 1393.50 | 1008.72 | 1.15 | 0.05 ± 0.00 A | 0.04 ± 0.00 B | 0.05 ± 0.00 A | 0.03 ± 0.00 C |
| Ethylpyrazine | C13925003 | 1369.20 | 956.84 | 1.11 | 0.12 ± 0.00 B | 0.11 ± 0.00 C | 0.12 ± 0.00 BC | 0.18 ± 0.02 A |
| 2,5-Dimethylpyrazine | C123320 | 1350.20 | 918.30 | 1.12 | 0.26 ± 0.01 B | 0.28 ± 0.01 B | 0.26 ± 0.01 B | 0.37 ± 0.02 A |
| 2-Methylpyrazine | C109080 | 1274.70 | 781.11 | 1.08 | 0.58 ± 0.01 A | 0.53 ± 0.01 B | 0.51 ± 0.01 B | 0.59 ± 0.01 C |
| Aromatic hydrocarbons | ||||||||
| p-Xylene | C106423 | 1143.90 | 560.95 | 1.08 | 0.12 ± 0.01 B | 0.11 ± 0.00 C | 0.12 ± 0.00 BC | 0.18 ± 0.00 A |
| Toluene | C108883 | 1062.30 | 433.19 | 1.04 | 0.60 ± 0.01 B | 0.60 ± 0.01 C | 0.60 ± 0.00 B | 0.77 ± 0.01 A |
| Pyridines | ||||||||
| 3-Ethylpyridine | C536787 | 1384.60 | 989.45 | 1.10 | 0.06 ± 0.00 A | 0.06 ± 0.00 A | 0.06 ± 0.00 A | 0.05 ± 0.00 B |
| Others | ||||||||
| α-Terpinene | C99865 | 1189.60 | 652.91 | 1.23 | 0.18 ± 0.01 A | 0.16 ± 0.00 B | 0.14 ± 0.00 C | 0.19 ± 0.00 B |
| α-Phellandrene | C99832 | 1175.10 | 622.26 | 1.67 | 0.14 ± 0.00 B | 0.13 ± 0.00 C | 0.13 ± 0.01 C | 0.21 ± 0.01 A |
| β-Pinene | C127913 | 1143.90 | 560.95 | 1.22 | 0.29 ± 0.00 A | 0.25 ± 0.00 C | 0.23 ± 0.01 C | 0.32 ± 0.00 B |
| Tetrahydrothiophene-M | C110010 | 1121.10 | 520.07 | 1.05 | 0.58 ± 0.01 B | 0.61 ± 0.01 B | 0.59 ± 0.01 B | 0.78 ± 0.01 A |
| Tetrahydrothiophene-D | C110010 | 1121.10 | 520.07 | 1.31 | 0.35 ± 0.00 A | 0.31 ± 0.01 B | 0.31 ± 0.01 B | 0.44 ± 0.03 A |
| Dimethyl disulfide | C624920 | 1081.60 | 458.76 | 1.14 | 0.11 ± 0.01 BC | 0.09 ± 0.02 C | 0.15 ± 0.03 B | 0.34 ± 0.04 A |
| Thiophene | C110021 | 1028.00 | 391.31 | 1.04 | 1.58 ± 0.01 A | 1.65 ± 0.03 A | 1.45 ± 0.06 B | 1.70 ± 0.05 C |
| Acrylonitrile | C107131 | 1011.70 | 372.86 | 1.08 | 0.39 ± 0.00 B | 0.43 ± 0.02 A | 0.39 ± 0.00 B | 0.42 ± 0.00 C |
| Dimethyl sulfide | C75183 | 788.30 | 222.92 | 1.09 | 2.24 ± 0.02 A | 1.92 ± 0.11 B | 2.21 ± 0.00 A | 2.22 ± 0.05 B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, T.; Chen, J.; Xu, J.; Gu, H.; Zhao, P.; Wang, W.; Pan, S.; Tao, Y.; Wang, H.; Yang, J. Screening of a Novel Lactiplantibacillus plantarum MMB-05 and Lacticaseibacillus casei Fermented Sandwich Seaweed Scraps: Chemical Composition, In Vitro Antioxidant, and Volatile Compounds Analysis by GC-IMS. Foods 2022, 11, 2875. https://doi.org/10.3390/foods11182875
Gao T, Chen J, Xu J, Gu H, Zhao P, Wang W, Pan S, Tao Y, Wang H, Yang J. Screening of a Novel Lactiplantibacillus plantarum MMB-05 and Lacticaseibacillus casei Fermented Sandwich Seaweed Scraps: Chemical Composition, In Vitro Antioxidant, and Volatile Compounds Analysis by GC-IMS. Foods. 2022; 11(18):2875. https://doi.org/10.3390/foods11182875
Chicago/Turabian StyleGao, Tengqi, Jinling Chen, Jie Xu, Han Gu, Pengpeng Zhao, Wenbin Wang, Saikun Pan, Yang Tao, Hongli Wang, and Jie Yang. 2022. "Screening of a Novel Lactiplantibacillus plantarum MMB-05 and Lacticaseibacillus casei Fermented Sandwich Seaweed Scraps: Chemical Composition, In Vitro Antioxidant, and Volatile Compounds Analysis by GC-IMS" Foods 11, no. 18: 2875. https://doi.org/10.3390/foods11182875