
Probiotic Oxalate-Degrading Bacteria: New Insight of Environmental Variables and Expression of the oxc and frc Genes on Oxalate Degradation Activity



Abstract
:1. Introduction
Chemistry of Oxalate
2. Oxalate Sources in the Body
3. Oxalate Content Estimation Methods
4. Degradation of Oxalate by Obligate and Facultative Anaerobic Gut Bacteria
5. Variables Affecting the Activity of Oxalate Degrading Bacteria
5.1. pH
5.2. Glucose and Sucrose Concentration
5.3. Yeast Extract
5.4. Bacterial Age
5.5. Inulin
5.6. Antibiotics
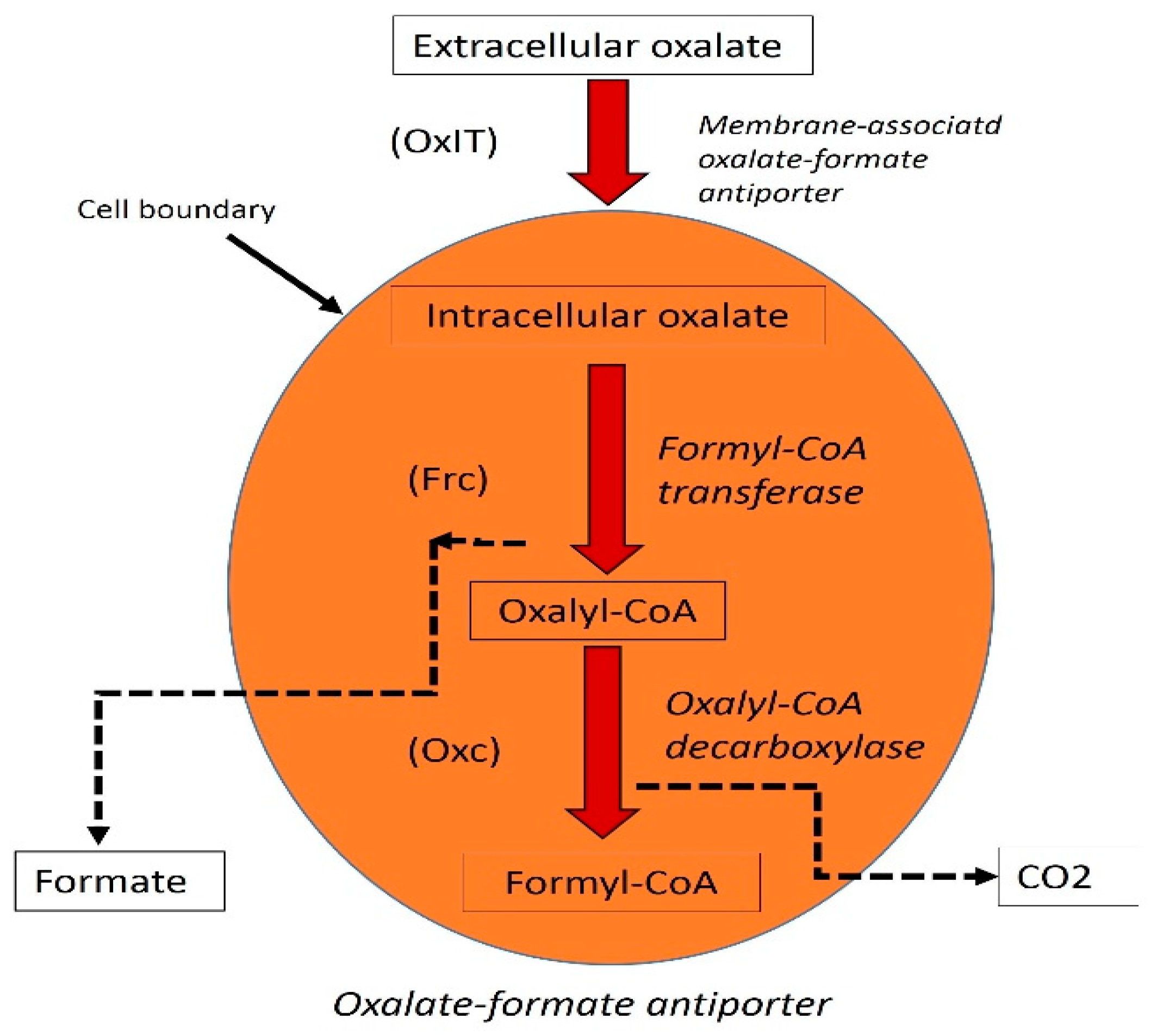
6. Analysis of Transcription and Function of the oxc and frc Genes in LAB and O. formigenes
7. Discussion
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kullin, B.R.; Reid, S.J.; Abratt, V.R. The use of probiotic bacteria to treat recurrent calcium oxalate kidney stone disease. In The Role of Bacteria in Urology; Lange, D., Chew, B., Eds.; Springer: Cham, Switzerland, 2016; pp. 63–75. [Google Scholar] [CrossRef]
- Batagello, C.A.; Monga, M.; Miller, A.W. Calcium oxalate urolithiasis: A case of missing microbes? J. Endourol. 2018, 32, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Shee, K.; Stoller, L. Perspectives in primary hyperoxaluria—Historical, current and future clinical interventions. Nat. Rev. Urol. 2022, 19, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Falk, P.G.; Hooper, L.V.; Midtvedt, T.; Gordon, J.I. Creating and maintaining the gastrointestinal ecosystem: What we know and need to know from gnotobiology. Microbiol. Mol. Biol. Rev. 1998, 62, 1157–1170. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Peck, A.B.; Khan, S.R. NADPH oxidase as a therapeutic target for oxalate induced injury in kidneys. Oxid. Med. Cell Longev. 2013, 2013, 462361. [Google Scholar] [CrossRef]
- Figueres, L.; Hourmant, M.; Lemoine, S. Understanding and managing hypercalciuria in adults with nephrolithiasis: Keys for nephrologists. Nephrol. Dial. Transplant. 2020, 35, 573–575. [Google Scholar] [CrossRef]
- Castellaro, A.M.; Tonda, A.; Cejas, H.H.; Ferreyra, H.; Caputto, B.L.; Pucci, O.A.; Gil, G.A. Oxalate induces breast cancer. BMC Cancer 2015, 15, 761. [Google Scholar] [CrossRef]
- Rahman, M.M.; Abdullah, R.B.; Khadijah, W.E.W. A review of oxalate poisoning in domestic animals: Tolerance and performance aspects. J. Anim. Physiol. Anim. Nutr. 2013, 97, 605–619. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, Y.H.; Chi, Z.P.; Huang, R.; Huang, H.; Liu, G.; Zhang, Y.; Yang, H.; Lin, J.; Yang, T.; et al. The handling of oxalate in the body and the origin of oxalate in calcium oxalate stones. Urol. Int. 2020, 104, 167–176. [Google Scholar] [CrossRef]
- Oranusi, S.; Adedeji, O.M.; Olopade, B.K. Probiotics in management of diseases: A review. Int. J. Curr. Res. Acad. Rev. 2014, 2, 138–158. [Google Scholar]
- Fargue, S.; Milliner, D.S.; Knight, J.; Olson, J.B.; Lowther, W.T.; Holmes, R.P. Hydroxyproline Metabolism and Oxalate Syn-thesis in Primary Hyperoxaluria. J. Am. Soc. Nephrol. 2018, 29, 1615–1623. [Google Scholar] [CrossRef] [Green Version]
- Gianmoena, K.; Gasparoni, N.; Jashari, A.; Gabrys, P.; Grgas, K.; Ghallab, A.; Nordström, K.; Gasparoni, G.; Reinders, J.; Ed-lund, K.; et al. Epigenomic and transcriptional profiling identifies impaired glyoxylate detoxification in NAFLD as a risk fac-tor for hyperoxaluria. Cell Rep. 2021, 36, 109526. [Google Scholar] [CrossRef]
- Lange, J.N.; Wood, K.D.; Knight, J.; Assimos, D.G.; Holmes, R.P. Glyoxal formation and its role in endogenous oxalate syn-thesis. Adv. Urol. 2012, 2012, 819202. [Google Scholar] [CrossRef]
- Turroni, S.; Vitali, B.; Bendazzoli, C.; Candela, M.; Gotti, R.; Federici, F.; Pirovano, F.; Brigidi, P. Oxalate consumption by lac-tobacilli: Evaluation of oxalyl-CoA decarboxylase and formyl-CoA transferase activity in Lactobacillus acidophilus. J. Appl. Microbiol. 2007, 103, 1600–1609. [Google Scholar] [CrossRef]
- Miller, A.W.; Dearing, D. The metabolic and ecological interactions of oxalate-degrading bacteria in mammalian gut. Pathogens 2013, 2, 636–652. [Google Scholar] [CrossRef]
- Wigner, P.; Bijak, M.; Saluk-Bijak, J. Probiotics in the Prevention of the Calcium Oxalate Urolithiasis. Cells 2022, 11, 284. [Google Scholar] [CrossRef]
- Hatch, M.; Freel, R.W.; Vaziri, N.D. Mechanisms of oxalate absorption and secretion across the rabbit distal colon. Pflug. Arch. 1994, 426, 101–109. [Google Scholar] [CrossRef]
- Ohlweiler, O.A.; Schneider, M.H. Standardization of potassium permanganate by titration of sodium oxalate in presence of perchloric acid and manganese(II) sulfate. Anal. Chim. Acta 1972, 58, 477–480. [Google Scholar] [CrossRef]
- Karamad, D.; Khosravi-Darani, K.; Hosseini, H.; Tavasoli, S. Analytical procedures and methods validation for oxalate con-tent estimation. Biointerface Res. Appl. Chem. 2019, 9, 4305–4310. [Google Scholar] [CrossRef]
- Murru, N.; Blaiotta, G.; Peruzy, M.F.; Santonicola, S.; Mercogliano, R.; Aponte, M. Screening of oxalate degrading lactic acid bacteria of food origin. Ital. J. Food Saf. 2017, 6, 6345. [Google Scholar] [CrossRef]
- Allison, M.J.; Dawson, K.A.; Mayberry, W.R.; Foss, J.G. Oxalobacter formigenes gen. nov.; sp. nov.: Oxalate-degrading anaer-obes that inhabit the gastrointestinal tract. Arch. Microbiol. 1985, 141, 147. [Google Scholar] [CrossRef]
- Nazzal, L.; Francois, F.; Henderson, N.; Liu, M.; Li, H.; Koh, H.; Wang, C.; Gao, Z.; Perez, G.P.; Asplin, J.R. Effect of antibiotic treatment on Oxalobacter formigenes colonization of the gut microbiome and urinary oxalate excretion. Sci. Rep. 2021, 11, 16428. [Google Scholar] [CrossRef]
- Stewart, C.S.; Duncan, S.H.; Cave, D.R. Oxalobacter formigenes and its role in oxalate metabolism in the human gut. FEMS Microbiol. Lett. 2004, 230, 1–7. [Google Scholar] [CrossRef]
- Arasu, M.V.; Al-Dhabi, N.A.; Ilavenil, S.; Choi, K.C.; Srigopalram, S. In vitro importance of probiotic Lactobacillus planta-rum related to medical field. Saudi J. Biol. Sci. 2016, 23, S6–S10. [Google Scholar] [CrossRef]
- PeBenito, A.; Nazzal, L.; Wang, C.; Li, H.; Jay, M.; Noya-Alarcon, O.; Contreras, M.; Lander, O.; Leach, J.; Dominguez-Bello, M.G.; et al. Comparative prevalence of Oxalobacter formigenes in three human populations. Sci. Rep. 2019, 9, 574. [Google Scholar] [CrossRef]
- Jiang, T.; Chen, W.; Cao, L.; He, Y.; Zhou, H.; Mao, H. Abundance, Functional, and evolutionary analysis of ox-alyl-coenzyme a decarboxylase in human microbiota. Front. Microbiol. 2020, 11, 672. [Google Scholar] [CrossRef]
- Campieri, C.; Campieri, M.; Bertuzzi, V.; Swennen, E.; Matteuzzi, D.; Stefoni, S.; Pirovano, F.; Centi, C.; Ulisse, S.; Famularo, G.; et al. Reduction of oxaluria after an oral course of lactic acid bacteria at high concentration. Kidney Int. 2001, 60, 1097–1105. [Google Scholar] [CrossRef]
- Federici, F.; Vitali, B.; Gotti, R.; Pasca, M.R.; Gobbi, S.; Peck, A.B.; Brigidi, P. Characterization and heterologous expression of the oxalyl coenzyme A decarboxylase gene from Bifidobacterium lactis. Appl. Environ. Microbiol. 2004, 70, 5066–5073. [Google Scholar] [CrossRef]
- Chamberlain, C.A.; Hatch, M.; Garrett, T.J. Metabolomic profiling of oxalate-degrading probiotic Lactobacillus acidophilus and Lactobacillus gasseri. PLoS ONE 2019, 14, e0222393. [Google Scholar] [CrossRef]
- Azcarate-Peril, M.A.; Bruno-Bárcena, J.M.; Hassan, H.M.; Klaenhammer, T.R. Transcriptional and functional analysis of ox-alyl-coenzyme A (CoA) decarboxylase and formyl-CoA transferase genes from Lactobacillus acidophilus. Appl. Environ. Microbiol. 2006, 72, 1891–1899. [Google Scholar] [CrossRef]
- Lewanika, T.R.; Reid, S.J.; Abratt, V.R.; Macfarlane, G.T.; Macfarlane, S. Lactobacillus gasseri Gasser AM63(T) de-grades oxa-late in a multistage continuous culture simulator of the human colonic microbiota. FEMS Microbiol. Ecol. 2007, 61, 110–120. [Google Scholar] [CrossRef] [Green Version]
- Karamad, D.; Khosravi-Darani, K.; Hosseini, H.; Tavasoli, S.; Miller, A.W. Assessment of the process variables for degrada-tion of oxalate by Lactobacillus acidophilus ATCC 4356 using simulated rumen fluid media and tea. Appl. Food Biotechnol. 2020, 7, 195–204. [Google Scholar] [CrossRef]
- Karamad, D.; Khosravi-Darani, K.; Hosseini, H.; Tavasoli, S.; Miller, A.W. Evaluation of Oxalobacter formigenes DSM 4420 biodegradation activity for high oxalate media content: An in vitro model. Biocatal. Agric. Biotechnol. 2019, 22, 101378. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.W.; Choy, D.; Penniston, K.L.; Lange, D. Inhibition of urinary stone disease by a multi-species bacterial network ensures healthy oxalate homeostasis. Kidney Int. 2019, 96, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Dawson, K.A.; Milton, A.; Hartman, P. Characteristics of anaerobic oxalate-degrading enrichment cultures from the rumen. Appl. Environ. Microbiol. 1980, 40, 840–846. [Google Scholar] [CrossRef]
- Gholami, Z.; Khosravi-Darani, K. An overview of conjugated linoleic acid: Microbial production and application. Mini Rev. Med. Chem. 2014, 14, 734–746. [Google Scholar] [CrossRef]
- Balthazar, C.F.; Silva, H.L.; Esmerino, E.A.; Rocha, R.S.; Moraes, J.; Carmo, M.A.; Azevedo, L.; Camps, I.; Abud, Y.K.; Sant’Anna, C.; et al. The addition of inulin and Lactobacillus casei 01 in sheep milk ice cream. Food Chem. 2018, 246, 464–472. [Google Scholar] [CrossRef]
- Stepanova, N.; Akulenko, I.; Serhiichuk, T.; Dovbynchuk, T.; Savchenko, S.; Tolstanova, G. Synbiotic supplementation and oxalate homeostasis in rats: Focus on microbiota oxalate-degrading activity. Urolithiasis 2022, 50, 249–258. [Google Scholar] [CrossRef]
- Darilmaz, D.; Sonmez, O.S.; Beyatli, Y. The effects of inulin as a prebiotic supplement and the symbiotic interactions of probi-otics to improve oxalate degrading activity. Int. J. Food Sci. 2018, 54, 121–131. [Google Scholar] [CrossRef]
- Lange, J.N.; Wood, K.D.; Wong, H.; Otto, R.; Mufarrij, P.W.; Knight, J.; Akpinar, H.; Holmes, R.P.; Assimos, D.G. Sensitivity of human strains of Oxalobacter formigenes to commonly prescribed antibiotics. Urology 2012, 79, 1286–1289. [Google Scholar] [CrossRef]
- Scotland, K.; Lange, D. The link between antibiotic exposure and kidney stone disease. Ann. Transl. Med. 2018, 6, 371. [Google Scholar] [CrossRef]
- Ricagno, S.; Jonsson, S.; Richards, N.; Lindqvis, Y. Formyl-CoA transferase encloses the CoA binding site at the interface of an interlocked dimer. EMBO J. 2003, 22, 3210–3219. [Google Scholar] [CrossRef]
- Knight, J.; Deora, R.; Assimos, D.G.; Holmes, R.P. The genetic composition of Oxalobacter formigenes and its relationship to colonization and calcium oxalate stone disease. Urolithiasis 2013, 41, 187–196. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in hu-mans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Nazzal, L.; Ho, M.; Wu, M.; Charytan, D.M. Oxalate Nephropathy in an Oxalobacter formigenes-Negative Subject. Kidney Int. Rep. 2020, 5, 754–757. [Google Scholar] [CrossRef]
- Liu, M.; Devlin, J.C.; Hu, J.; Volkova, A.; Battaglia, T.W.; Ho, M.; Asplin, J.R.; Byrd, A.; Loke, P.; Li, H.; et al. Microbial genetic and transcriptional contributions to oxalate degradation by the gut microbiota in health and disease. eLife 2021, 10, e63642. [Google Scholar] [CrossRef]
- Jafari, G.A.; Ardakani, R.F.; Sepahi, M.A.; Nowroozi, J.; Soltanpour, M.S. Development of an innovative method by optimizing qpcr technique for isolating and determining Oxalobacter formigenes microbial load in the stool of patients with urolithiasis. Iran J. Kidney Dis. 2021, 15, 190–198. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Sources | Reference |
|---|---|---|
| Lactiplantibacillus plantarum | YOMO Research Centre | [27] |
| Lactobacillus brevis | ||
| Lactobacillus acidophilus | ||
| Bifidobacterium infantis | ||
| Bifidobacterium animalis ATCC 27536 | ||
| Bifidobacterium breve MB 283 | Gut intestine human | [28] |
| Bifidobacterium longum MB 282 | ||
| Bifidobacterium infantis MB 57 | ||
| Bifidobacterium adolescentis MB 238 | ||
| Lacticaseibacillus casei | Gut intestine human | [29] |
| Lactobacillus acidophilus | Gut intestine human | [30] |
| Lactobacillus gasseri Gasser AM63T | Gut intestine human | [31] |
| Lactobacillus acidophilus | ||
| Lactobacillus gasseri | ||
| Lactiplantibacillus plantarum | ||
| Lacticaseibacillus casei | ||
| Lacticaseibacillus rhamnosus | ||
| Ligiactobacillus salivarius |
| Antibiotic | HC1 | Va3 | Cc13 | OxK |
|---|---|---|---|---|
| Amoxicillin | R | R | R | R |
| Amoxicillin/Clavulanate | R | R | R | R |
| Azithromycin | S | S | S | S |
| Ceftriaxone | R | R | R | R |
| Cephalexin | R | R | R | R |
| Ciprofloxacin | S | S | S | S |
| Clarithromycin | S | S | S | S |
| Clindamycin | S | S | S | S |
| Doxycycline | S | S | S | S |
| Gentamicin | S | S | S | S |
| Levofloxacin | S | S | S | S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karamad, D.; Khosravi-Darani, K.; Khaneghah, A.M.; Miller, A.W. Probiotic Oxalate-Degrading Bacteria: New Insight of Environmental Variables and Expression of the oxc and frc Genes on Oxalate Degradation Activity. Foods 2022, 11, 2876. https://doi.org/10.3390/foods11182876
Karamad D, Khosravi-Darani K, Khaneghah AM, Miller AW. Probiotic Oxalate-Degrading Bacteria: New Insight of Environmental Variables and Expression of the oxc and frc Genes on Oxalate Degradation Activity. Foods. 2022; 11(18):2876. https://doi.org/10.3390/foods11182876
Chicago/Turabian StyleKaramad, Dina, Kianoush Khosravi-Darani, Amin Mousavi Khaneghah, and Aaron W. Miller. 2022. "Probiotic Oxalate-Degrading Bacteria: New Insight of Environmental Variables and Expression of the oxc and frc Genes on Oxalate Degradation Activity" Foods 11, no. 18: 2876. https://doi.org/10.3390/foods11182876