
NMR-Based Metabolomics to Decipher the Molecular Mechanisms in the Action of Gut-Modulating Foods



Abstract
:1. Introduction
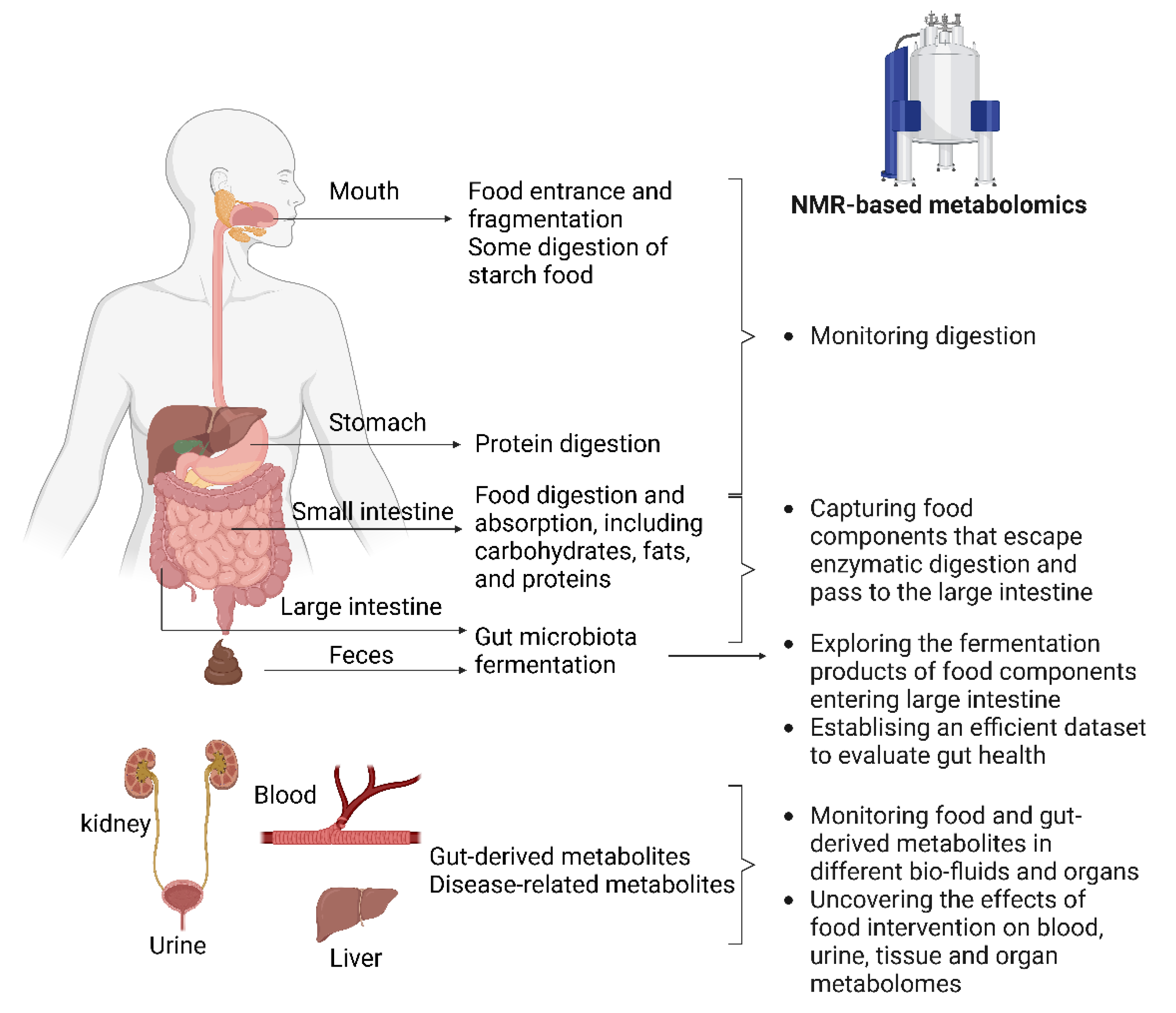
2. Advantages and Limitations of NMR-Based Metabolomics
3. Multiple Purposes of NMR-Based Metabolomics in Food and Nutrition Science
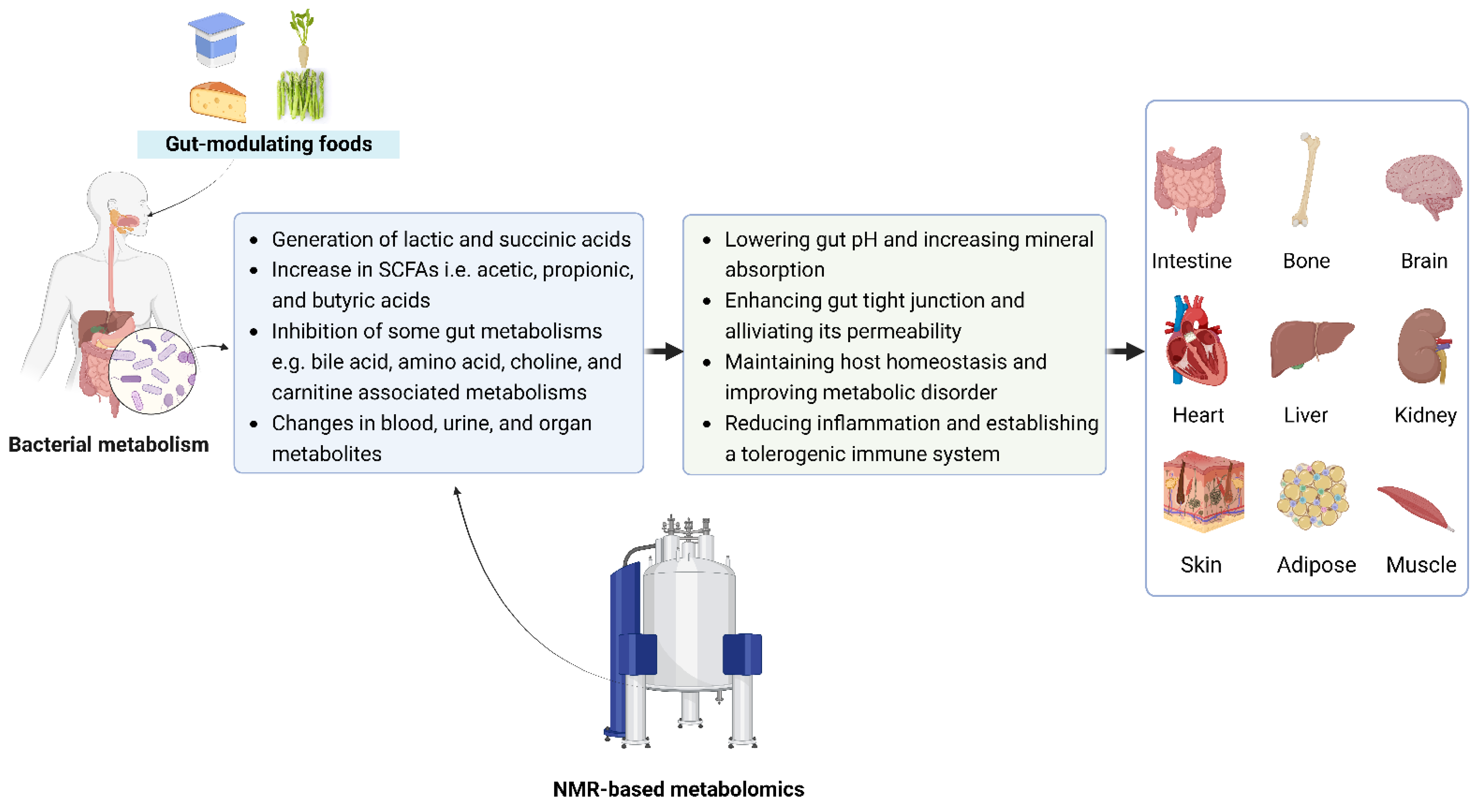
4. Metabolomics Reveals the Beneficial Effects of Saccharolytic Activity on Gut-Organ Axis
5. Gut Modulating Foods in the Suppression of Unfavorable Metabolites
6. Calcium as a New Functional Food Ingredient?
7. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wörheide, M.A.; Krumsiek, J.; Kastenmüller, G.; Arnold, M. Multi-omics integration in biomedical research—A metabolomics-centric review. Anal. Chim. Acta 2021, 1141, 144–162. [Google Scholar] [CrossRef]
- Brennan, L.; Hu, F.B.; Sun, Q. Metabolomics meets nutritional epidemiology: Harnessing the potential in metabolomics data. Metabolites 2021, 11, 709. [Google Scholar] [CrossRef]
- Lamichhane, S.; Sen, P.; Dickens, A.M.; Orešič, M.; Bertram, H.C. Gut metabolome meets microbiome: A methodological perspective to understand the relationship between host and microbe. Methods 2018, 149, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Lanng, S.K.; Zhang, Y.; Christensen, K.R.; Hansen, A.K.; Nielsen, D.S.; Kot, W.; Bertram, H.C. Partial substitution of meat with insect (Alphitobius diaperinus) in a carnivore diet changes the gut microbiome and metabolome of healthy rats. Foods 2021, 10, 1814. [Google Scholar] [CrossRef] [PubMed]
- Thøgersen, R.; Castro-Mejía, J.L.; Kræmer Sundekilde, U.; Hansen, L.H.; Gray, N.; Kuhnle, G.; Rye Jørgensen, N.; Kornerup Hansen, A.; Sandris Nielsen, D.; Bertram, H.C. Inulin and milk mineral fortification of a pork sausage exhibits distinct effects on the microbiome and biochemical activity in the gut of healthy rats. Food Chem. 2020, 331, 127291. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhong, D.; Ren, Y.Y.; Meng, Z.K.; Pegg, R.B.; Zhong, G. Effect of konjac glucomannan on metabolites in the stomach, small intestine and large intestine of constipated mice and prediction of the KEGG pathway. Food Funct. 2021, 12, 3044–3056. [Google Scholar] [CrossRef]
- Cui, M.; Trimigno, A.; Aru, V.; Khakimov, B.; Engelsen, S.B. Human Faecal 1H NMR Metabolomics: Evaluation of Solvent and Sample Processing on Coverage and Reproducibility of Signature Metabolites. Anal. Chem. 2020, 92, 9546–9555. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, S.; Yde, C.C.; Schmedes, M.S.; Jensen, H.M.; Meier, S.; Bertram, H.C. Strategy for nuclear-magnetic-resonance-based metabolomics of human feces. Anal. Chem. 2015, 87, 5930–5937. [Google Scholar] [CrossRef] [PubMed]
- Thøgersen, R.; Castro-Mejía, J.L.; Sundekilde, U.K.; Hansen, L.H.; Hansen, A.K.; Nielsen, D.S.; Bertram, H.C. Ingestion of an inulin-enriched pork sausage product positively modulates the gut microbiome and metabolome of healthy rats. Mol. Nutr. Food Res. 2018, 62, 1800608. [Google Scholar] [CrossRef] [PubMed]
- Thøgersen, R.; Rasmussen, M.K.; Sundekilde, U.K.; Goethals, S.A.; Van Hecke, T.; Vossen, E.; De Smet, S.; Bertram, H.C. Background diet influences TMAO concentrations associated with red meat intake without influencing apparent hepatic TMAO-related activity in a porcine model. Metabolites 2020, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowda, G.N.; Raftery, D. NMR-Based Metabolomics Methods and Protocols Methods; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Wishart, D.S. NMR metabolomics: A look ahead. J. Magn. Reson. 2019, 306, 155–161. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Timbrell, J.A.; Sadler, P.J. Proton NMR spectra of urine as indicators of renal damage. Mercury-induced nephrotoxicity in rats. Mol. Pharmacol. 1985, 27, 644–651. [Google Scholar]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. “Metabonomics”: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Emwas, A.H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Nagana Gowda, G.A.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR spectroscopy for metabolomics research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.G.; Park, J.Y.; Lee, J.; Bang, E.; Kim, S.R.; Lee, E.K.; Yun, H.J.; Kang, C.M.; Hwang, G.S. Investigation of relative metabolic changes in the organs and plasma of rats exposed to X-ray radiation using HR-MAS 1H NMR and solution 1H NMR. NMR Biomed. 2016, 29, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Jenne, A.; Bermel, W.; Michal, C.A.; Gruschke, O.; Soong, R.; Ghosh Biswas, R.; Bastawrous, M.; Simpson, A.J. DREAMTIME NMR Spectroscopy: Targeted Multi-Compound Selection with Improved Detection Limits. Angew. Chemie Int. Ed. 2022, 61, e202110044. [Google Scholar] [CrossRef]
- Appiah-Amponsah, E.; Owusu-Sarfo, K.; Gowda, G.A.N.; Ye, T.; Raftery, D. Combining hydrophilic interaction chromatography (HILIC) and isotope tagging for off-line LC-NMR applications in metabolite analysis. Metabolites 2013, 3, 575–591. [Google Scholar] [CrossRef]
- Khakimov, B.; Mobaraki, N.; Trimigno, A.; Aru, V.; Engelsen, S.B. Signature Mapping (SigMa): An efficient approach for processing complex human urine 1H NMR metabolomics data. Anal. Chim. Acta 2020, 1108, 142–151. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Xie, Z.; Thøgersen, R.; Rasmussen, M.K.; Zachariassen, L.F.; Jørgensen, N.R.; Nørgaard, J.V.; Andersen, H.J.; Nielsen, D.S.; Hansen, A.K.; et al. Effects of Calcium Source, Inulin, and Lactose on Gut-Bone Associations in an Ovarierectomized Rat Model. Mol. Nutr. Food Res. 2022, 66, 2100883. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, X.; Li, H.; Ren, Y.; Geng, Y.; Lu, Z.; Shi, J.; Xu, Z. Similarities and differences of oligo/poly-saccharides’ impact on human fecal microbiota identified by in vitro fermentation. Appl. Microbiol. Biotechnol. 2021, 105, 7475–7486. [Google Scholar] [CrossRef]
- Harris, H.C.; Morrison, D.J.; Edwards, C.A. Impact of the source of fermentable carbohydrate on SCFA production by human gut microbiota in vitro—A systematic scoping review and secondary analysis. Crit. Rev. Food Sci. Nutr. 2021, 61, 3892–3903. [Google Scholar] [CrossRef] [PubMed]
- Schmedes, M.; Brejnrod, A.D.; Aadland, E.K.; Kiilerich, P.; Kristiansen, K.; Jacques, H.; Lavigne, C.; Graff, I.E.; Eng, Ø.; Holthe, A.; et al. The Effect of Lean-Seafood and Non-Seafood Diets on Fecal Metabolites and Gut Microbiome: Results from a Randomized Crossover Intervention Study. Mol. Nutr. Food Res. 2019, 63, 1700976. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Bergeron, N.; Levison, B.S.; Li, X.S.; Chiu, S.; Xun, J.; Koeth, R.A.; Lin, L.; Wu, Y.; Tang, W.H.W.; et al. Impact of chronic dietary red meat, white meat, or non-meat protein on trimethylamine N-oxide metabolism and renal excretion in healthy men and women. Eur. Heart J. 2019, 40, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Ma, C.; Liu, C.; Wang, Z.; Yang, J.; Liu, X.; Shen, Z.; Wu, R. NMR-based fecal metabolomics fingerprinting as predictors of earlier diagnosis in patients with colorectal cancer. Oncotarget 2016, 7, 29454–29464. [Google Scholar] [CrossRef] [PubMed]
- Vernocchi, P.; Del Chierico, F.; Putignani, L. Gut microbiota profiling: Metabolomics based approach to unravel compounds affecting human health. Front. Microbiol. 2016, 7, 1144. [Google Scholar] [CrossRef]
- Smirnov, K.S.; Maier, T.V.; Walker, A.; Heinzmann, S.S.; Forcisi, S.; Martinez, I.; Walter, J.; Schmitt-Kopplin, P. Challenges of metabolomics in human gut microbiota research. Int. J. Med. Microbiol. 2016, 306, 266–279. [Google Scholar] [CrossRef]
- Bervoets, L.; Ippel, J.H.; Smolinska, A.; Van Best, N.; Savelkoul, P.H.M.; Mommers, M.A.H.; Penders, J. Practical and Robust NMR-Based Metabolic Phenotyping of Gut Health in Early Life. J. Proteome Res. 2021, 20, 5079–5087. [Google Scholar] [CrossRef]
- Alinaghi, M.; Nguyen, D.N.; Bertram, H.C.; Sangild, P.T. Direct implementation of intestinal permeability test in nmr metabolomics for simultaneous biomarker discovery—A feasibility study in a preterm piglet model. Metabolites 2020, 10, 22. [Google Scholar] [CrossRef]
- Carvalho, D.V.; Silva, L.M.A.; Alves Filho, E.G.; Santos, F.A.; Lima, R.P.; De Viana, A.F.S.C.; Nunes, P.I.G.; Fonseca, S.G.D.C.; De Melo, T.S.; Viana, D.D.A.; et al. Cashew apple fiber prevents high fat diet-induced obesity in mice: An NMR metabolomic evaluation. Food Funct. 2019, 10, 1671–1683. [Google Scholar] [CrossRef]
- Hanifa, M.A.; Skott, M.; Maltesen, R.G.; Rasmussen, B.S.; Nielsen, S.; Frøkiær, J.; Ring, T.; Wimmer, R. Tissue, urine and blood metabolite signatures of chronic kidney disease in the 5/6 nephrectomy rat model. Metabolomics 2019, 15, 1–16. [Google Scholar] [CrossRef]
- Stepnowski, P.; Kaczor, M.; Proczko-Stepaniak, M.; Kaska, L.; Sledzinski, T.; Mika, A.; Kaczynski, Z. Potential Application of 1H NMR for Routine Serum Lipidome Analysis -Evaluation of Effects of Bariatric Surgery. Sci. Rep. 2017, 7, 15530. [Google Scholar]
- Khakimov, B.; Hoefsloot, H.C.J.; Mobaraki, N.; Aru, V.; Kristensen, M.; Lind, M.V.; Holm, L.; Castro-Mejía, J.L.; Nielsen, D.S.; Jacobs, D.M.; et al. Human Blood Lipoprotein Predictions from 1H NMR Spectra: Protocol, Model Performances, and Cage of Covariance. Anal. Chem. 2022, 94, 628–636. [Google Scholar] [CrossRef]
- Kostara, C.E.; Lekkas, P.; Vezyraki, P.; Angelidis, C.; Deligiannis, I.K.; Bairaktari, E.T.; Kalfakakou, V. Lipidome of plasma lipoproteins and liver is zinc- modulated in High fat diet treated mice. J. Trace Elem. Med. Biol. 2018, 50, 268–275. [Google Scholar] [CrossRef]
- ten Hove, M.; Pater, L.; Storm, G.; Weiskirchen, S.; Weiskirchen, R.; Lammers, T.; Bansal, R. The hepatic lipidome: From basic science to clinical translation. Adv. Drug Deliv. Rev. 2020, 159, 180–197. [Google Scholar] [CrossRef]
- Smilde, A.K.; Jansen, J.J.; Hoefsloot, H.C.J.; Lamers, R.J.A.N.; van der Greef, J.; Timmerman, M.E. ANOVA-simultaneous component analysis (ASCA): A new tool for analyzing designed metabolomics data. Bioinformatics 2005, 21, 3043–3048. [Google Scholar] [CrossRef]
- Yde, C.C.; Clausen, M.R.; Ditlev, D.B.; Lillefosse, H.; Madsen, L.; Kristiansen, K.; Liaset, B.; Bertram, H.C. Multi-block PCA and multi-compartmental study of the metabolic responses to intake of hydrolysed versus intact casein in C57BL/6J mice by NMR-based metabolomics. Metabolomics 2014, 10, 938–949. [Google Scholar] [CrossRef]
- Alinaghi, M.; Bertram, H.C.; Brunse, A.; Smilde, A.K.; Westerhuis, J.A. Common and distinct variation in data fusion of designed experimental data. Metabolomics 2020, 16, 1–11. [Google Scholar] [CrossRef]
- Liu, H.M.; Lin, X.; Meng, X.H.; Zhao, Q.; Shen, J.; Xiao, H.M.; Deng, H.W. Integrated metagenome and metabolome analyses of blood pressure studies in early postmenopausal Chinese women. J. Hypertens. 2021, 39, 1800–1809. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Kläring, K.; Heinzmann, S.S.; Platz, S.; Scholz, B.; Engel, K.H.; Schmitt-Kopplin, P.; Haller, D.; Rohn, S.; Skurk, T.; et al. Gut metabolites and bacterial community networks during a pilot intervention study with flaxseeds in healthy adult men. Mol. Nutr. Food Res. 2015, 59, 1614–1628. [Google Scholar] [CrossRef]
- Antharam, V.C.; McEwen, D.C.; Garrett, T.J.; Dossey, A.T.; Li, E.C.; Kozlov, A.N.; Mesbah, Z.; Wang, G.P. An integrated metabolomic and microbiome analysis identified specific gut microbiota associated with fecal cholesterol and coprostanol in Clostridium difficile infection. PLoS ONE 2016, 11, e0148824. [Google Scholar] [CrossRef]
- Das, Q.; Islam, M.R.; Lepp, D.; Tang, J.; Yin, X.; Mats, L.; Liu, H.; Ross, K.; Kennes, Y.M.; Yacini, H.; et al. Gut Microbiota, Blood Metabolites, and Spleen Immunity in Broiler Chickens Fed Berry Pomaces and Phenolic-Enriched Extractives. Front. Vet. Sci. 2020, 7, 150. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Min, L.; Zhang, W.; Liu, J.; Hou, Z.; Chu, M.; Li, L.; Shen, W.; Zhao, Y.; Zhang, H. Zinc oxide nanoparticles influence microflora in ileal digesta and correlate well with blood metabolites. Front. Microbiol. 2017, 8, 992. [Google Scholar] [CrossRef]
- Ma, J.; Sun, L.; Liu, Y.; Ren, H.; Shen, Y.; Bi, F.; Zhang, T.; Wang, X. Alter between gut bacteria and blood metabolites and the anti-tumor effects of Faecalibacterium prausnitzii in breast cancer. J. Clin. Oncol. 2020, 38, e12575. [Google Scholar] [CrossRef]
- Clos-Garcia, M.; Andrés-Marin, N.; Fernández-Eulate, G.; Abecia, L.; Lavín, J.L.; van Liempd, S.; Cabrera, D.; Royo, F.; Valero, A.; Errazquin, N.; et al. Gut microbiome and serum metabolome analyses identify molecular biomarkers and altered glutamate metabolism in fibromyalgia. EBioMedicine 2019, 46, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Li, Y.; Zhang, W.; Fan, M.; Qian, H.; Zhang, H.; Qi, X.; Wang, L. Effects of cereal fibers on short-chain fatty acids in healthy subjects and patients: A meta-analysis of randomized clinical trials. Food Funct. 2021, 12, 7040–7053. [Google Scholar] [CrossRef] [PubMed]
- Hills, R.D.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut microbiome: Profound implications for diet and disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Ahlawat, S.; Asha; Sharma, K.K. Gut–organ axis: A microbial outreach and networking. Lett. Appl. Microbiol. 2021, 72, 636–668. [Google Scholar] [CrossRef] [PubMed]
- Frampton, J.; Murphy, K.G.; Frost, G.; Chambers, E.S. Short-chain fatty acids as potential regulators of skeletal muscle metabolism and function. Nat. Metab. 2020, 2, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wang, Y.; Yang, G.; Zhang, Q.; Meng, L.; Xin, Y.; Jiang, X. The role of short-chain fatty acids in intestinal barrier function, inflammation, oxidative stress, and colonic carcinogenesis. Pharmacol. Res. 2021, 165, 105420. [Google Scholar] [CrossRef] [PubMed]
- Couto, M.R.; Gonçalves, P.; Magro, F.; Martel, F. Microbiota-derived butyrate regulates intestinal inflammation: Focus on inflammatory bowel disease. Pharmacol. Res. 2020, 159, 104947. [Google Scholar] [CrossRef] [PubMed]
- Salvi, P.S.; Cowles, R.A. Butyrate and the intestinal epithelium: Modulation of proliferation and inflammation in homeostasis and disease. Cells 2021, 10, 1775. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Davis, B.; Zhu, W.; Zheng, N.; Di Meng; Walker, W.A. Short-chain fatty acid butyrate, a breast milk metabolite, enhances immature intestinal barrier function genes in response to inflammation in vitro and in vivo. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 320, G521–G530. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Cao, M.M.; Li, Y.J.; Dai, G.C.; Lu, P.P.; Zhang, M.; Bai, L.Y.; Chen, X.X.; Zhang, C.; Shi, L.; et al. The regulative effect and repercussion of probiotics and prebiotics on osteoporosis: Involvement of brain-gut-bone axis. Crit. Rev. Food Sci. Nutr. 2022, 2, 1–19. [Google Scholar] [CrossRef]
- Bakirhan, H.; Karabudak, E. Effects of inulin on calcium metabolism and bone health. Int. J. Vitam. Nutr. Res. 2021, 1, 1–12. [Google Scholar] [CrossRef]
- Slevin, M.M.; Allsopp, P.J.; Magee, P.J.; Bonham, M.P.; Naughton, V.R.; Strain, J.J.; Duffy, M.E.; Wallace, J.M.; McSorley, E.M. Supplementation with calcium and short-chain fructo-oligosaccharides affects markers of bone turnover but not bone mineral density in postmenopausal women. J. Nutr. 2014, 144, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Geiger, S.; Patra, A.K.; Schrapers, K.T.; Braun, H.S.; Aschenbach, J.R. Menthol stimulates calcium absorption in the rumen but not in the jejunum of sheep. J. Dairy Sci. 2021, 104, 3067–3081. [Google Scholar] [CrossRef]
- Lucas, S.; Omata, Y.; Hofmann, J.; Böttcher, M.; Iljazovic, A.; Sarter, K.; Albrecht, O.; Schulz, O.; Krishnacoumar, B.; Krönke, G.; et al. Short-chain fatty acids regulate systemic bone mass and protect from pathological bone loss. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zaiss, M.M.; Jones, R.M.; Schett, G.; Pacifici, R. The gut-bone axis: How bacterial metabolites bridge the distance. J. Clin. Investig. 2019, 129, 3018–3028. [Google Scholar] [CrossRef]
- Li, S.; Mao, Y.; Zhou, F.; Yang, H.; Shi, Q.; Meng, B. Gut microbiome and osteoporosis. Bone Jt. Res. 2020, 9, 524–530. [Google Scholar] [CrossRef]
- Takahashi, D.; Hoshina, N.; Kabumoto, Y.; Maeda, Y.; Suzuki, A.; Tanabe, H.; Isobe, J.; Yamada, T.; Muroi, K.; Yanagisawa, Y.; et al. Microbiota-derived butyrate limits the autoimmune response by promoting the differentiation of follicular regulatory T cells. EBioMedicine 2020, 58, 102913. [Google Scholar] [CrossRef]
- Li, J.Y.; Yu, M.; Pal, S.; Tyagi, A.M.; Dar, H.; Adams, J.; Neale Weitzmann, M.; Jones, R.M.; Pacifici, R. Parathyroid hormone-dependent bone formation requires butyrate production by intestinal microbiota. J. Clin. Investig. 2020, 130, 1767–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Takakura, A.; Zandi-Nejad, K.; Charles, J.F. Mechanisms of gut microbiota-mediated bone remodeling. Gut Microbes 2018, 9, 84–92. [Google Scholar] [CrossRef]
- Jaimes, J.D.; Slavíčkova, A.; Hurych, J.; Cinek, O.; Nichols, B.; Vodolanova, L.; Cerny, K.; Havlik, J. Stool metabolome-microbiota evaluation among children and adolescents with obesity, overweight, and normal-weight using 1H NMR and 16S rRNA gene profiling. PLoS ONE 2021, 16, e0247378. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef]
- Chain, S.; Acids, F.; Microbiota, F. Short Chain Fatty Acids and Fecal Microbiota Abundance in Humans with Obesity: A Systematic Review and Meta-Analysis. Nutrients 2019, 11, 2512. [Google Scholar]
- Yamamura, R.; Nakamura, K.; Ukawa, S.; Okada, E.; Nakagawa, T.; Imae, A.; Kunihiro, T.; Kimura, T.; Hirata, T.; Tamakoshi, A. Fecal short-chain fatty acids and obesity in a community-based Japanese population: The DOSANCO Health Study. Obes. Res. Clin. Pract. 2021, 15, 345–350. [Google Scholar] [CrossRef]
- Canfora, E.E.; Blaak, E.E. Acetate: A diet-derived key metabolite in energy metabolism: Good or bad in context of obesity and glucose homeostasis? Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A double-edged sword for health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Boulangé, C.L.; Neves, A.L.; Chilloux, J.; Nicholson, J.K.; Dumas, M.E. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Vojinovic, D.; Radjabzadeh, D.; Kurilshikov, A.; Amin, N.; Wijmenga, C.; Franke, L.; Ikram, M.A.; Uitterlinden, A.G.; Zhernakova, A.; Fu, J.; et al. Relationship between gut microbiota and circulating metabolites in population-based cohorts. Nat. Commun. 2019, 10, 5813. [Google Scholar] [CrossRef]
- Bartel, J.; Krumsiek, J.; Schramm, K.; Adamski, J.; Gieger, C.; Herder, C.; Carstensen, M.; Peters, A.; Rathmann, W.; Roden, M.; et al. The Human Blood Metabolome-Transcriptome Interface. PLoS Genet. 2015, 11, e1005274. [Google Scholar] [CrossRef] [PubMed]
- Fechine, C.P.N.D.S.; Monteiro, M.G.C.A.; Tavares, J.F.; Souto, A.L.; Luna, R.C.P.; da Silva, C.S.O.; da Silva, J.A.; Dos Santos, S.G.; de Carvalho Costa, M.J.; Persuhn, D.C. Choline metabolites, hydroxybutyrate and hdl after dietary fiber supplementation in overweight/obese hypertensive women: A metabolomic study. Nutrients 2021, 13, 1437. [Google Scholar] [CrossRef]
- Dai, B.; Huang, S.; Deng, Y. Modified insoluble dietary fibers in okara affect body composition, serum metabolic properties, and fatty acid profiles in mice fed high-fat diets: An NMR investigation. Food Res. Int. 2019, 116, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Crovesy, L.; El-Bacha, T.; Rosado, E.L. Modulation of the gut microbiota by probiotics and symbiotics is associated with changes in serum metabolite profile related to a decrease in inflammation and overall benefits to metabolic health: A double-blind randomized controlled clinical trial in wom. Food Funct. 2021, 12, 2161–2170. [Google Scholar] [CrossRef]
- Sivanesan, S.; Taylor, A.; Zhang, J.; Bakovic, M. Betaine and Choline Improve Lipid Homeostasis in Obesity by Participation in Mitochondrial Oxidative Demethylation. Front. Nutr. 2018, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Randell, E.; Zhou, H.; Sun, G. Higher serum choline and betaine levels are associated with better body composition in male but not female population. PLoS ONE 2018, 13, e0193114. [Google Scholar] [CrossRef] [PubMed]
- Mey, J.T.; Erickson, M.L.; Axelrod, C.L.; King, W.T.; Flask, C.A.; McCullough, A.J.; Kirwan, J.P. β-Hydroxybutyrate is reduced in humans with obesity-related NAFLD and displays a dose-dependent effect on skeletal muscle mitochondrial respiration in vitro. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E184–E195. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.C.; Candow, D.G.; Krentz, J.R.; Roberts, M.D.; Young, K.C. Changes in fat mass following creatine supplementation and resistance training in adults ≥50 years of age: A meta-analysis. J. Funct. Morphol. Kinesiol. 2019, 4, 62. [Google Scholar] [CrossRef]
- Sharma, V.; Sharma, N.; Sheikh, I.; Kumar, V.; Sehrawat, N.; Yadav, M.; Ram, G.; Sankhyan, A.; Sharma, A.K. Probiotics and Prebiotics Having Broad Spectrum Anticancer Therapeutic Potential: Recent Trends and Future Perspectives. Curr. Pharmacol. Reports 2021, 7, 67–79. [Google Scholar] [CrossRef]
- Zheng, Y.; He, J.-Q. Pathogenic Mechanisms of Trimethylamine N-Oxide-induced Atherosclerosis and Cardiomyopathy. Curr. Vasc. Pharmacol. 2021, 20, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Qiu, J.; Lian, J.; Yang, X.; Zhou, J. Gut Metabolite Trimethylamine-N-Oxide in Atherosclerosis: From Mechanism to Therapy. Front. Cardiovasc. Med. 2021, 8, 723886. [Google Scholar] [CrossRef] [PubMed]
- Arayici, M.E.; Mert-Ozupek, N.; Yalcin, F.; Basbinar, Y.; Ellidokuz, H. Soluble and Insoluble Dietary Fiber Consumption and Colorectal Cancer Risk: A Systematic Review and Meta-Analysis. Nutr. Cancer 2021, 74, 2412–2425. [Google Scholar] [CrossRef] [PubMed]
- Conlon, M.A.; Kerr, C.A.; Mcsweeney, C.S.; Dunne, R.A.; Shaw, J.M.; Kang, S.; Bird, A.R.; Morell, M.K.; Lockett, T.J.; Molloy, P.L.; et al. Resistant starches protect against colonic DNA damage and alter microbiota and gene expression in rats fed a western diet. J. Nutr. 2012, 142, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Thøgersen, R.; Bertram, H.C. Reformulation of processed meat to attenuate potential harmful effects in the gastrointestinal tract—A review of current knowledge and evidence of health prospects. Trends Food Sci. Technol. 2021, 108, 111–118. [Google Scholar] [CrossRef]
- Thøgersen, R.; Gray, N.; Kuhnle, G.; Van Hecke, T.; De Smet, S.; Young, J.F.; Sundekilde, U.K.; Hansen, A.K.; Bertram, H.C. Inulin-fortification of a processed meat product attenuates formation of nitroso compounds in the gut of healthy rats. Food Chem. 2020, 302, 125339. [Google Scholar] [CrossRef]
- Fuhren, J.; Schwalbe, M.; Boekhorst, J.; Rösch, C.; Schols, H.A.; Kleerebezem, M. Dietary calcium phosphate strongly impacts gut microbiome changes elicited by inulin and galacto-oligosaccharides consumption. Microbiome 2021, 9, 218. [Google Scholar] [CrossRef]
- Cha, K.H.; Yang, J.S.; Kim, K.A.; Yoon, K.Y.; Song, D.G.; Erdene-Ochir, E.; Kang, K.; Pan, C.H.; Ko, G.P. Improvement in host metabolic homeostasis and alteration in gut microbiota in mice on the high-fat diet: A comparison of calcium supplements. Food Res. Int. 2020, 136, 109495. [Google Scholar] [CrossRef]
- Gilbert, M.S.; Ijssennagger, N.; Kies, A.K.; van Mil, S.W.C. Protein fermentation in the gut; implications for intestinal dysfunction in humans, pigs, and poultry. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G159–G170. [Google Scholar] [CrossRef]
- Cai, J.; Chen, Z.; Wu, W.; Lin, Q.; Liang, Y. High animal protein diet and gut microbiota in human health. Crit. Rev. Food Sci. Nutr. 2021, 62, 6225–6237. [Google Scholar] [CrossRef]
- Tan, B.; Chen, J.; Qin, S.; Liao, C.; Zhang, Y.; Wang, D.; Li, S.; Zhang, Z.; Zhang, P.; Xu, F. Tryptophan Pathway-Targeted Metabolomics Study on the Mechanism and Intervention of Cisplatin-Induced Acute Kidney Injury in Rats. Chem. Res. Toxicol. 2021, 34, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, S.; Inagi, R. Harnessing Metabolomics to Describe the Pathophysiology Underlying Progression in Diabetic Kidney Disease. Curr. Diab. Rep. 2021, 21, 21. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Y.; Chen, D.-Q.; Chen, L.; Liu, J.-R.; Vaziri, N.D.; Guo, Y.; Zhao, Y.-Y. Microbiome–metabolome reveals the contribution of gut–kidney axis on kidney disease. J. Transl. Med. 2019, 17, 5. [Google Scholar] [CrossRef] [Green Version]
- Gill, P.A.; van Zelm, M.C.; Muir, J.G.; Gibson, P.R. Review article: Short chain fatty acids as potential therapeutic agents in human gastrointestinal and inflammatory disorders. Aliment. Pharmacol. Ther. 2018, 48, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, R.; Wang, S.; Solberg Woods, L.C.; Seshie, O.; Chung, S.T.; Shively, C.A.; Register, T.C.; Craft, S.; McClain, D.A.; Yadav, H. Comparative microbiome signatures and short-chain fatty acids in mouse, rat, non-human primate, and human feces. Front. Microbiol. 2018, 9, 2897. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Metabolite Class | Metabolites |
|---|---|
| Branched-chain amino acids | Leucine, Isoleucine, Valine |
| Aromatic amino acids | Phenylalanine, Tyrosine, Tryptophan |
| Other amino acids | Alanine, Arginine, Asparagine, Aspartate, Glutamate, Glutamine, Glycine, Histidine, Lysine, Methionine, Proline, Threonine, Sarcosine, Serine |
| Amino acid derivatives | p-cresol, 4-hydroxyphenyllactate, N-phenylacetylglycine, Acetoin, Creatine, Creatinine |
| Short-chain fatty acids | Acetate, Butyrate, Formate, Propionate, Butyrate, Isobutyrate, Valerate |
| Other organic acids | Bile acids, Caprylate, Citrate, Fumarate, Glycolate, Isocaproate, Lactate, Malate, Malonate, Phenylacetate, Pyruvate, Succinate, Taurine |
| Mono and disaccharides | Fructose, Glucose, Galactose, Lactose, Ribose, Xylose |
| Nucleotides and derivatives | Cytidine, Hypoxanthine, Thymine, Uracil, Xanthine |
| Alcohols | Glycerol, Ethanol, Methanol |
| Esters | 2-methylbutyrate, 3-hydoxyphenlyacetate,4-hydroxyphenylacetate, Guanidoacetate, Methyl succinate |
| Alkaloids | Betaine, Carnitine, Choline, Dimethylamine, Ethanolamine, |
| Biogenic amines | Trimethylamine |
| Ketones | Acetone, Dihydroxyacetone |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, W.; Bertram, H.C. NMR-Based Metabolomics to Decipher the Molecular Mechanisms in the Action of Gut-Modulating Foods. Foods 2022, 11, 2707. https://doi.org/10.3390/foods11172707
He W, Bertram HC. NMR-Based Metabolomics to Decipher the Molecular Mechanisms in the Action of Gut-Modulating Foods. Foods. 2022; 11(17):2707. https://doi.org/10.3390/foods11172707
Chicago/Turabian StyleHe, Weiwei, and Hanne Christine Bertram. 2022. "NMR-Based Metabolomics to Decipher the Molecular Mechanisms in the Action of Gut-Modulating Foods" Foods 11, no. 17: 2707. https://doi.org/10.3390/foods11172707