
Synergistic Antibacterial Mechanism of Mannosylerythritol Lipid-A and Lactic Acid on Listeria monocytogenes Based on Transcriptomic Analysis
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Chemical Reagents
2.2. Synergistic Antibacterial Activity Assessment
2.3. RNA Isolation and Sequencing
2.4. Differential Expression Analysis
2.5. GO and KEGG Analysis of Differential Gene Expression
2.6. Real-Time qPCR Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Synergistic Antibacterial Activity of MEL-A and LA on L. monocytogenes
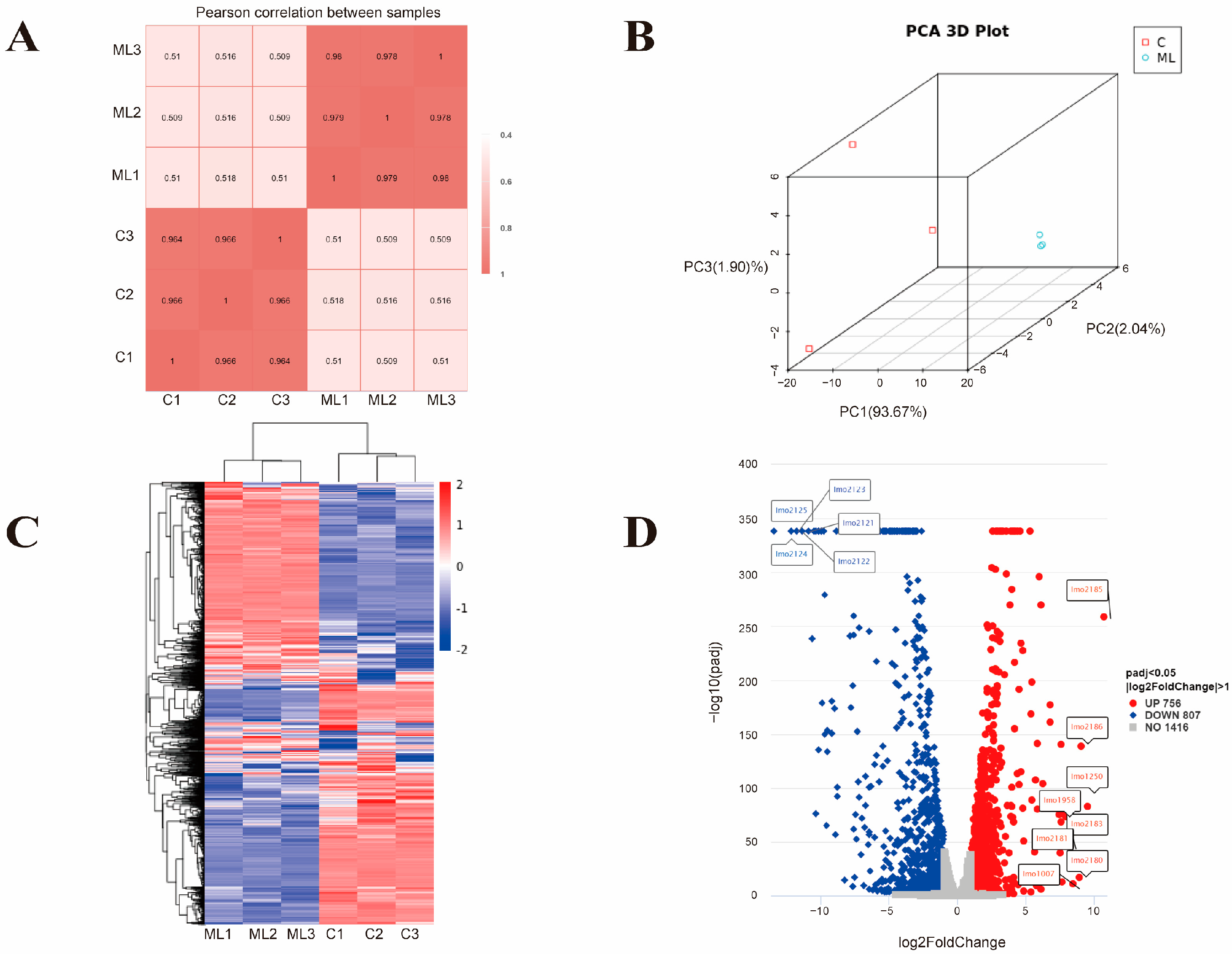
3.2. Effect of Synergistic Effect of MEL-A and LA on the Transcriptome Profile of L. monocytogenes
3.2.1. Global Response of L. monocytogenes to the Synergistic Treatment (ML Group)
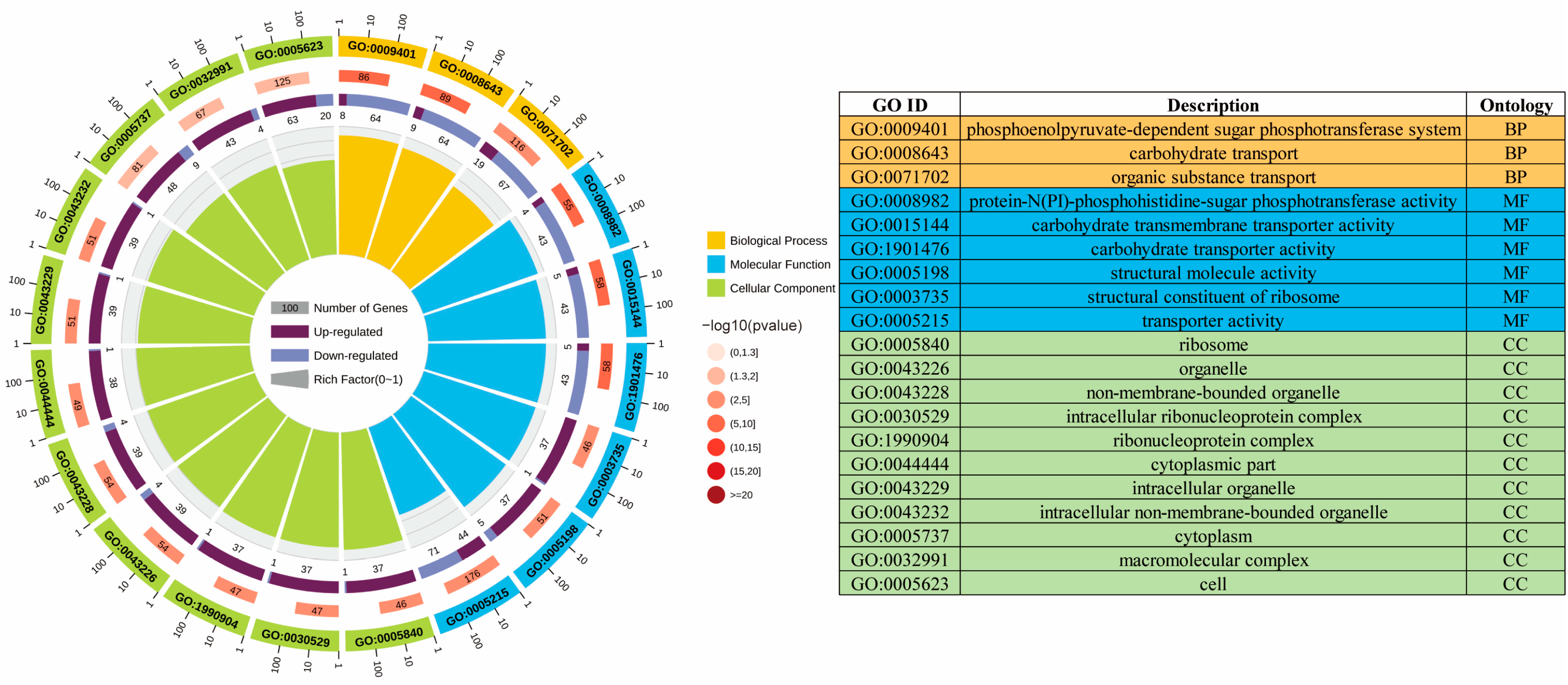
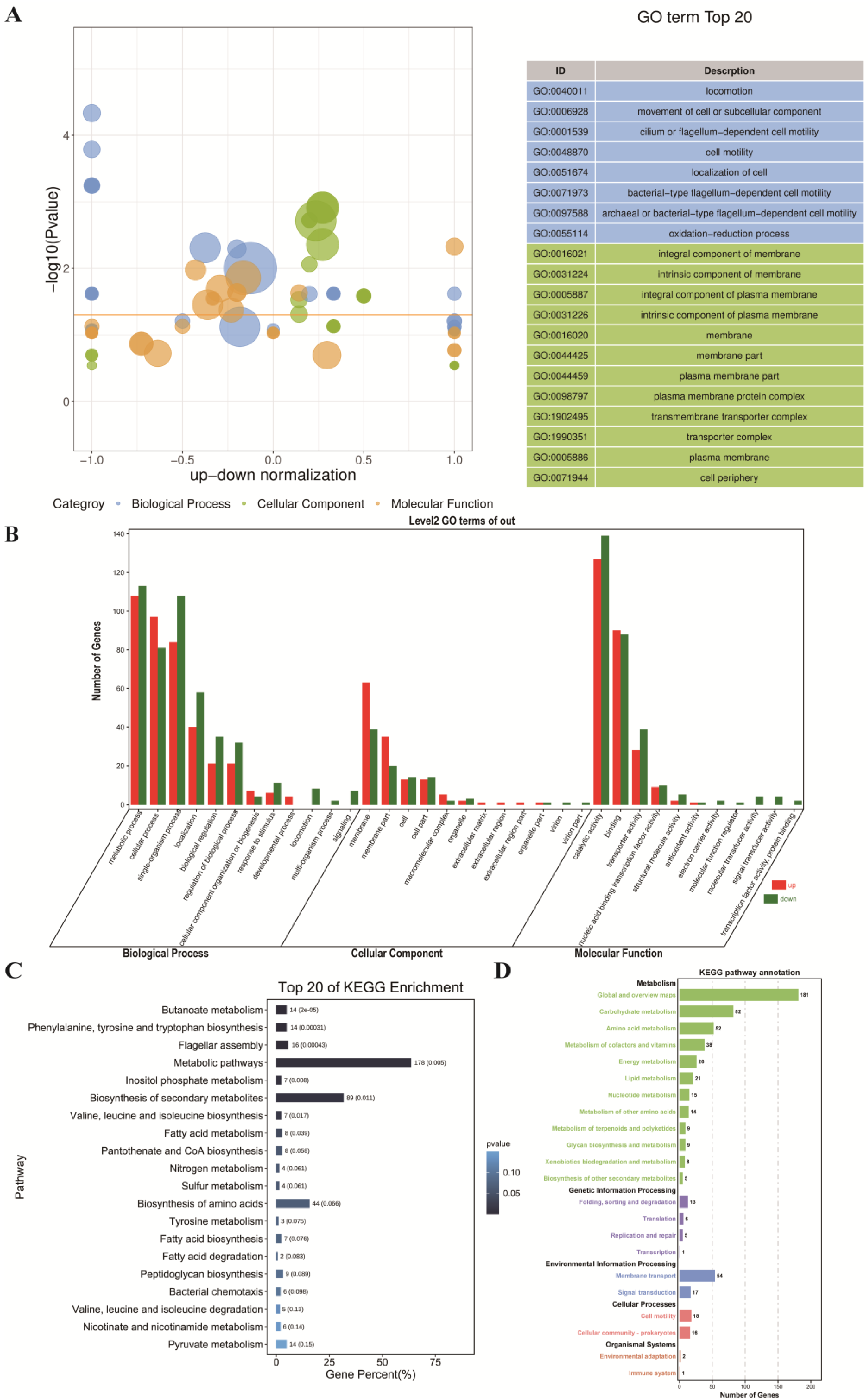
3.2.2. GO Enrichment Analysis of the ML Group and the Control Group
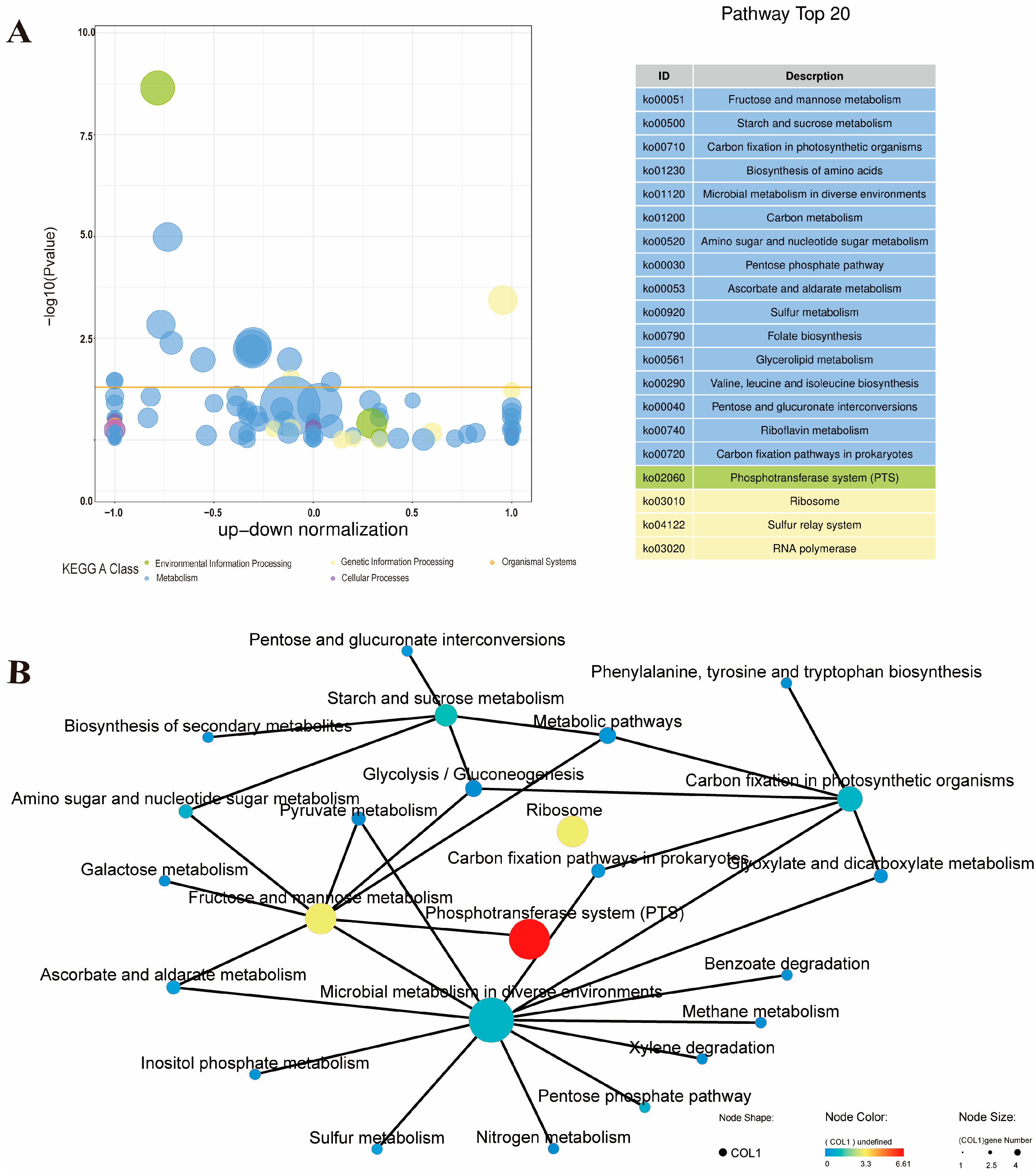
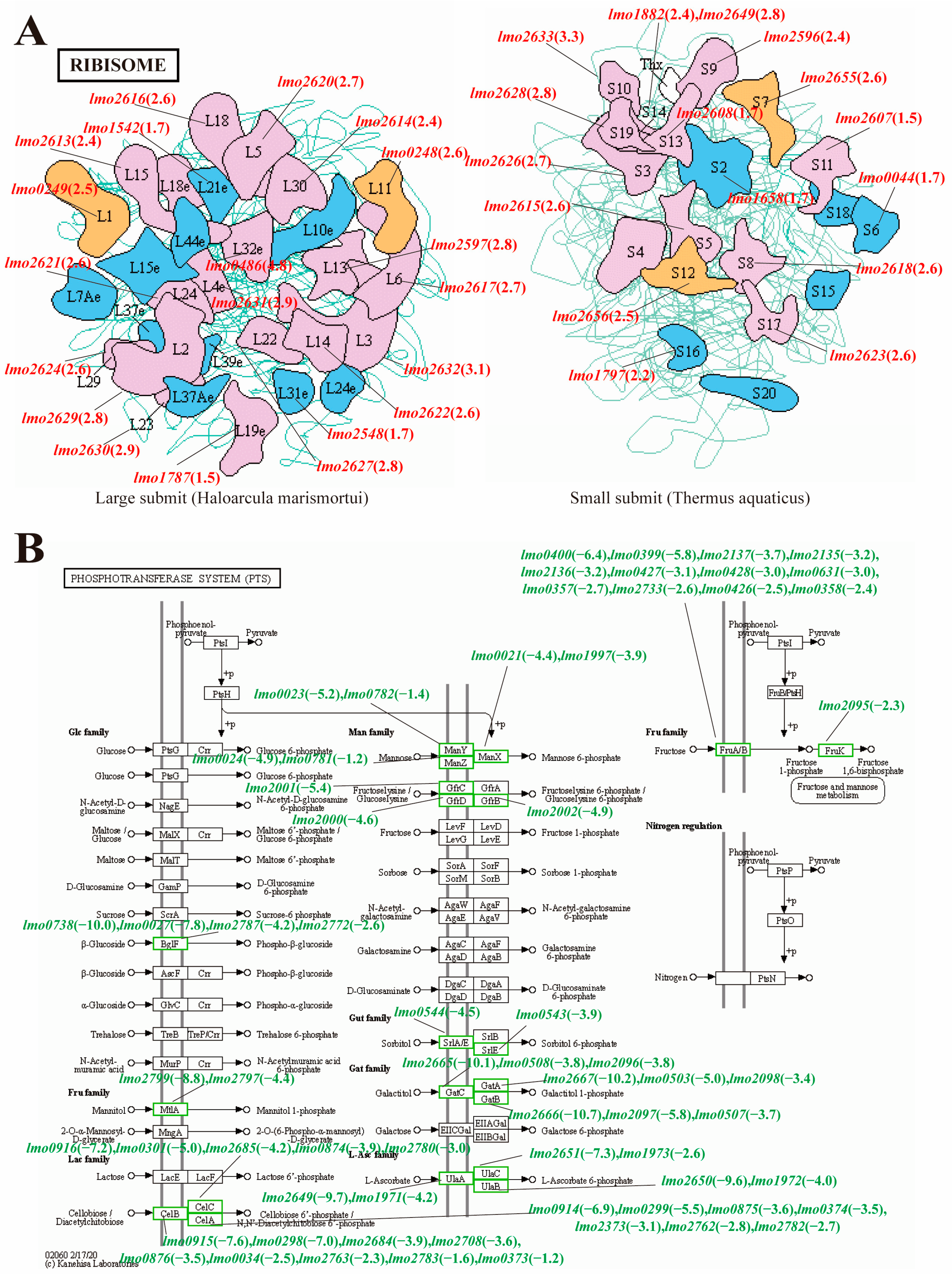
3.2.3. KEGG Pathway Enrichment Analysis of the ML Group and the Control Group
3.3. Comparative Transcriptome Analysis of the ML Co-Treatment Group and the LA Treatment Group Alone
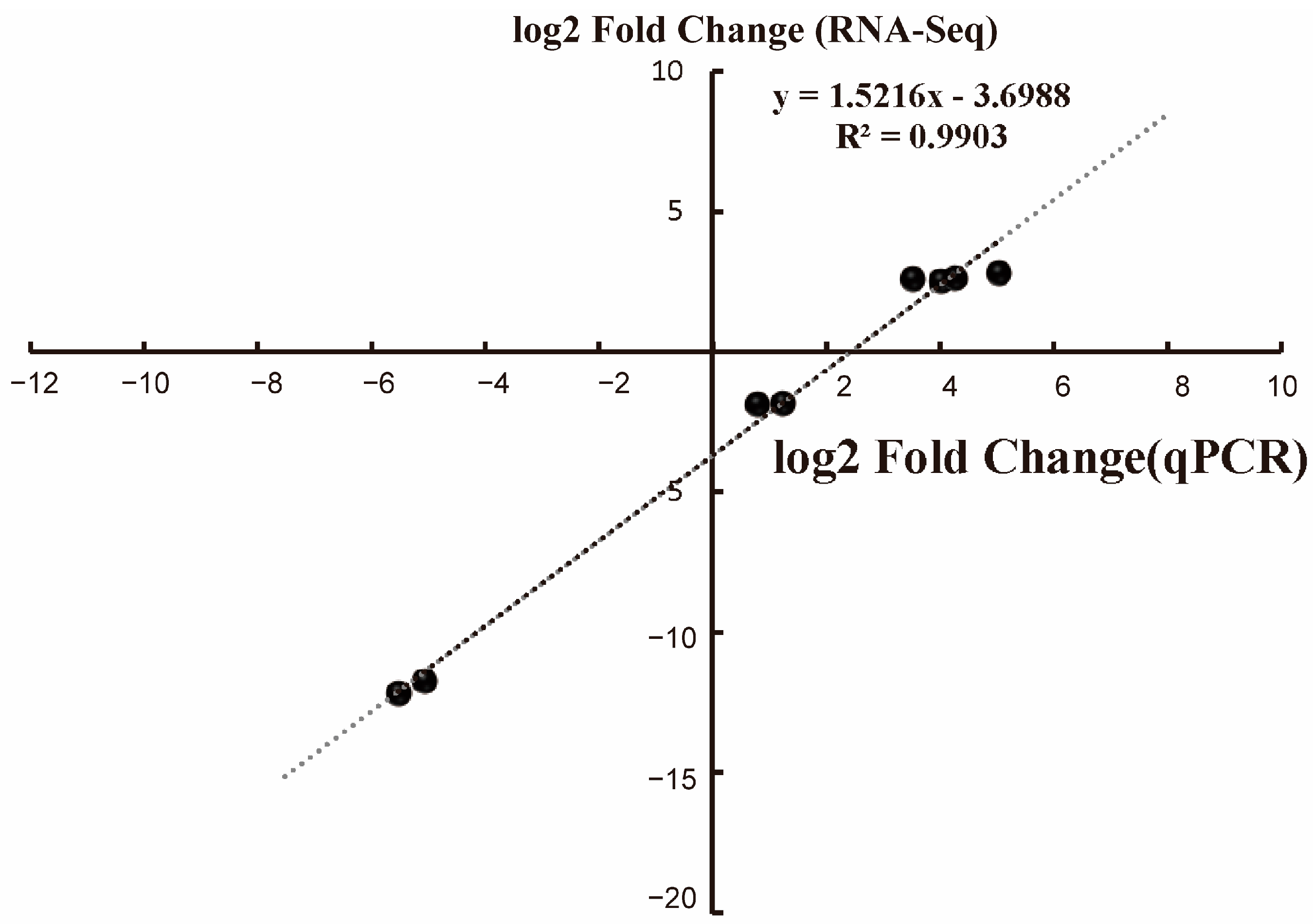
3.4. RT-qPCR Validation of RNA-Seq Data
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, X.; Liu, G.; Wu, Y.; Pang, X.; Wu, Y.; Qinshu; Niu, J.; Chen, Q.; Zhang, X. Transposon sequencing: A powerful tool for the functional genomic study of food-borne pathogens. Trends Food Sci. Technol. 2021, 118, 679–687. [Google Scholar] [CrossRef]
- Radoshevich, L.; Cossart, P. Listeria monocytogenes: Towards a complete picture of its physiology and pathogenesis. Nat. Rev. Microbiol. 2018, 16, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Gooneratne, R.; Hussain, M.A. Listeria monocytogenes in Fresh Produce: Outbreaks, Prevalence and Contamination Levels. Foods 2017, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Cossart, P. Illuminating the landscape of host-pathogen interactions with the bacterium Listeria monocytogenes. Proc. Natl. Acad. Sci. USA 2011, 108, 19484–19491. [Google Scholar] [CrossRef]
- Ozdemir, H.; Yildirim, Y.; Kuplulu, O.; Koluman, A.; Goncuoglu, M.; Inat, G. Effects of lactic acid and hot water treatments on Salmonella Typhimurium and Listeria monocytogenes on beef. Food Control 2006, 17, 299–303. [Google Scholar] [CrossRef]
- Byelashov, O.A.; Daskalov, H.; Geornaras, I.; Kendall, P.A.; Belk, K.E.; Scanga, J.A.; Smith, G.C.; Sofos, J.N. Reduction of Listeria monocytogenes on frankfurters treated with lactic acid solutions of various temperatures. Food Microbiol. 2010, 27, 783–790. [Google Scholar] [CrossRef]
- Wang, C.; Chang, T.; Yang, H.; Cui, M. Antibacterial mechanism of lactic acid on physiological and morphological properties of Salmonella Enteritidis, Escherichia coli and Listeria monocytogenes. Food Control 2015, 47, 231–236. [Google Scholar] [CrossRef]
- Lin, C.; Qin, L.; Xue, Z.; Hongfei, Z.; Xinyi, P.; Hongshun, Y. Inactivation efficacies of lactic acid and mild heat treatments against Escherichia coli strains in organic broccoli sprouts. Food Control 2022, 133 Pt A, 108577. [Google Scholar]
- Gonzalez-Fandos, E.; Martinez-Laorden, A.; Perez-Arnedo, I. Efficacy of combinations of lactic acid and potassium sorbate against Listeria monocytogenes in chicken stored under modified atmospheres. Food Microbiol. 2021, 93, 103596. [Google Scholar] [CrossRef]
- Capita, R.; Alonso-Calleja, C.; Rodriguez-Perez, R.; Moreno, B.; Garcia-Fernandez, M.D. Influence of poultry carcass skin sample site on the effectiveness of trisodium phosphate against Listeria monocytogenes. J Food Protect. 2002, 65, 853–856. [Google Scholar] [CrossRef]
- Kozak, S.M.; Margison, K.M.; D’Amico, D.J. Synergistic Antimicrobial Combinations Inhibit and Inactivate Listeria monocytogenes in Neutral and Acidic Broth Systems. J. Food Protect. 2017, 80, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Nashida, J.; Nishi, N.; Takahashi, Y.; Hayashi, C.; Igarashi, M.; Takahashi, D.; Toshima, K. Systematic and Stereoselective Total Synthesis of Mannosylerythritol Lipids and Evaluation of Their Antibacterial Activity. J. Org. Chem. 2018, 83, 7281–7289. [Google Scholar] [CrossRef] [PubMed]
- Silva Coelho, A.L.; Feuser, P.E.; Mattar Carciofi, B.A.; de Andrade, C.J.; de Oliveira, D. Mannosylerythritol lipids: Antimicrobial and biomedical properties. Appl. Microbiol. Biot. 2020, 104, 2297–2318. [Google Scholar] [CrossRef]
- Shu, Q.; Lou, H.; Wei, T.; Liu, X.; Chen, Q. Contributions of Glycolipid Biosurfactants and Glycolipid-Modified Materials to Antimicrobial Strategy: A Review. Pharmaceutics 2021, 13, 227. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.; Wei, T.; Lu, H.; Niu, Y.; Chen, Q. Mannosylerythritol lipids: Dual inhibitory modes against Staphylococcus aureus through membrane-mediated apoptosis and biofilm disruption. Appl. Microbiol. Biot. 2020, 104, 5053–5064. [Google Scholar] [CrossRef]
- Shu, Q.; Niu, Y.W.; Zhao, W.J.; Chen, Q.H. Antibacterial activity and mannosylerythritol lipids against vegetative cells and spores of Bacillus cereus. Food Control 2019, 106, 106711. [Google Scholar] [CrossRef]
- Qin, S.; Tianyu, W.; Xiayu, L.; Siyu, L.; Qihe, C. The dough-strengthening and spore-sterilizing effects of mannosylerythritol lipid-A in frozen dough and its application in bread making. Food Chem. 2022, 369, 131011. [Google Scholar]
- Liu, X.; Shu, Q.; Chen, Q.; Pang, X.; Wu, Y.; Zhou, W.; Wu, Y.; Niu, J.; Zhang, X. Antibacterial Efficacy and Mechanism of Mannosylerythritol Lipids-A on Listeria monocytogenes. Molecules 2020, 25, 4857. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, L.; Pang, X.; Wu, Y.; Wu, Y.; Shu, Q.; Chen, Q.; Zhang, X. Synergistic antibacterial effect and mechanism of high hydrostatic pressure and mannosylerythritol Lipid-A on Listeria monocytogenes. Food Control 2022, 135, 108797. [Google Scholar] [CrossRef]
- Shu, Q.; Lou, H.; Wei, T.; Zhang, X.; Chen, Q. Synergistic antibacterial and antibiofilm effects of ultrasound and MEL-A against methicillin-resistant Staphylococcus aureus. Ultrason. Sonochemistry 2021, 72, 105452. [Google Scholar] [CrossRef]
- Qian, J.; Zhang, M.; Dai, C.; Huo, S.; Ma, H. Transcriptomic analysis of Listeria monocytogenes under pulsed magnetic field treatment. Food Res. Int. 2020, 133, 109195. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Hu, X.; Gao, Z.; Li, G.; Fu, F.; Shang, X.; Liang, Z.; Shan, Y. Global transcriptomic response of Listeria monocytogenes exposed to Fingered Citron (Citrus medica L. var. sarcodactylis Swingle) essential oil. Food Res. Int. 2021, 143, 110274. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, S.; Xu, Y.; Mi, X.; Xing, T.; Li, J.; Zhang, L.; Gao, F.; Jiang, Y. Acid resistance of E. coli O157:H7 and O26:H11 exposure to lactic acid revealed by transcriptomic analysis. LWT-Food Sci. Technol. 2021, 136, 110352. [Google Scholar] [CrossRef]
- Becavin, C.; Bouchier, C.; Lechat, P.; Archambaud, C.; Creno, S.; Gouin, E.; Zongfu, W.; Kuhbacher, A.; Brisse, S.; Pucciarelli, M.G.; et al. Comparison of widely used Listeria monocytogenes strains EGD, 10403S, and EGD-e highlights genomic differences underlying variations in pathogenicity. MBio 2014, 5, e00969-14. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Li, H.; Niu, Y.; Chen, Q. Characterization and Inducing Melanoma Cell Apoptosis Activity of Mannosylerythritol Lipids-A Produced from Pseudozyma aphidis. PLoS ONE 2016, 11, e0148198. [Google Scholar]
- Kafantaris, I.; Tsadila, C.; Nikolaidis, M.; Tsavea, E.; Dimitriou, T.G.; Iliopoulos, I.; Amoutzias, G.D.; Mossialos, D. Transcriptomic Analysis of Pseudomonas aeruginosa Response to Pine Honey via RNA Sequencing Indicates Multiple Mechanisms of Antibacterial Activity. Foods 2021, 10, 936. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C-T method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Dog-Hwan, O.; Marshall, D.L. Antimicrobial activity of ethanol, glycerol monolaurate or lactic acid against Listeria monocytogenes. Int. J. Food Microbiol. 1993, 20, 239–246. [Google Scholar]
- Patro, R.; Mount, S.M.; Kingsford, C. Sailfish enables alignment-free isoform quantification from RNA-seq reads using lightweight algorithms. Nat. Biotechnol. 2014, 32, 462–464. [Google Scholar] [CrossRef]
- Ledala, N.; Sengupta, M.; Muthaiyan, A.; Wilkinson, B.J.; Jayaswal, R.K. Transcriptomic Response of Listeria monocytogenes to Iron Limitation and fur Mutation. Appl. Environ. Microb. 2010, 76, 406–416. [Google Scholar] [CrossRef]
- McLaughlin, H.P.; Hill, C.; Gahan, C.G.M. The impact of iron on Listeria monocytogenes; inside and outside the host. Curr. Opin. Biotechnol. 2011, 22, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Quistgaard, E.M.; Low, C.; Guettou, F.; Nordlund, P. Understanding transport by the major facilitator superfamily (MFS): Structures pave the way. Nat. Rev. Mol. Cell Biol. 2016, 17, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsen, C.; Brede, D.A.; Hernandez, P.E.; Nes, I.F.; Diep, D.B. The Maltose ABC Transporter in Lactococcus lactis Facilitates High-Level Sensitivity to the Circular Bacteriocin Garvicin ML. Antimicrob. Agents Chemother. 2012, 56, 2908–2915. [Google Scholar] [CrossRef] [PubMed]
- Lengeler, J.W.; Jahreis, K. Bacterial PEP-Dependent Carbohydrate: Phosphotransferase Systems Couple Sensing and Global Control Mechanisms. Bact. Sens. Signal. 2009, 16, 65–87. [Google Scholar]
- Gabor, E.; Goehler, A.-K.; Kosfeld, A.; Staab, A.; Kremling, A.; Jahreis, K. The phosphoenolpyruvate-dependent glucose-phosphotransferase system from Escherichia coli K-12 as the center of a network regulating carbohydrate flux in the cell. Eur. J. Cell Biol. 2011, 90, 711–720. [Google Scholar] [CrossRef]
- Metselaar, K.I.; den Besten, H.M.W.; Boekhorst, J.; van Hijum, S.A.F.T.; Zwietering, M.H.; Abee, T. Diversity of acid stress resistant variants of Listeria monocytogenes and the potential role of ribosomal protein S21 encoded by rpsU. Front. Microbiol. 2015, 6, 422. [Google Scholar] [CrossRef]
- Wu, Y.; Pang, X.; Liu, X.; Wu, Y.; Zhang, X. Functional Genomics Identified Novel Genes Involved in Growth at Low Temperatures in Listeria monocytogenes. Microbiol. Spectr. 2022, e00710-22. [Google Scholar] [CrossRef]
- Ramakrishnan, V. Ribosome structure and the mechanism of translation. Cell 2002, 108, 557–572. [Google Scholar] [CrossRef]
- Pechmann, S.; Willmund, F.; Frydman, J. The Ribosome as a Hub for Protein Quality Control. Mol. Cell 2013, 49, 411–421. [Google Scholar] [CrossRef]
- Poehlsgaard, J.; Douthwaite, S. The bacterial ribosome as a target for antibiotics. Nat. Rev. Microbiol. 2005, 3, 870–881. [Google Scholar] [CrossRef]
- Lin, J.; Zhou, D.; Steitz, T.A.; Polikanov, Y.S.; Gagnon, M.G. Ribosome-Targeting Antibiotics: Modes of Action, Mechanisms of Resistance, and Implications for Drug Design. Annu. Rev. Biochem. 2018, 87, 451–478. [Google Scholar] [CrossRef] [PubMed]
- Saier, M.H., Jr. The Bacterial Phosphotransferase System: New Frontiers 50 Years after Its Discovery. J. Mol. Microbiol. Biotechnol. 2015, 25, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Hua, X.; Xue, Z.; Ma, J. Cajanin Stilbene Acid Inhibited Vancomycin-Resistant Enterococcus by Inhibiting Phosphotransferase System. Front. Pharmacol. 2020, 11, 473. [Google Scholar] [CrossRef] [PubMed]
- Snyder, H.; Kellogg, S.L.; Skarda, L.M.; Little, J.L.; Kristich, C.J. Nutritional Control of Antibiotic Resistance via an Interface between the Phosphotransferase System and a Two-Component Signaling System. Antimicrob. Agents Chemother. 2014, 58, 957–965. [Google Scholar] [CrossRef]
- Huang, K.; Zeng, J.; Liu, X.; Jiang, T.; Wang, J. Structure of the mannose phosphotransferase system (man-PTS) complexed with microcin E492, a pore-forming bacteriocin. Cell Discov. 2021, 7, 1–5. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Log (CFU/mL) | Log Reduction (CFU/mL) | Antibacterial Rate (%) |
|---|---|---|---|
| Control | 9.44 ± 0.01 a | — | — |
| MEL-A (32 μg/mL) | 8.70 ± 0.03 c | 0.74 | 81.8 |
| LA (1 mg/mL) | 9.13 ± 0.10 b | 0.31 | 50.0 |
| LA (1 mg/mL) + MEL-A (32 μg/mL) | 7.14 ± 0.26 e | 2.30 | 99.4 |
| LA (2 mg/mL) | 8.46 ± 0.04 d | 0.98 | 89.4 |
| LA (2 mg/mL) + MEL-A (32 μg/mL) | 6.45 ± 0.09 f | 2.99 | 99.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Pang, X.; Wu, Y.; Wu, Y.; Shi, Y.; Zhang, X.; Chen, Q. Synergistic Antibacterial Mechanism of Mannosylerythritol Lipid-A and Lactic Acid on Listeria monocytogenes Based on Transcriptomic Analysis. Foods 2022, 11, 2660. https://doi.org/10.3390/foods11172660
Liu X, Pang X, Wu Y, Wu Y, Shi Y, Zhang X, Chen Q. Synergistic Antibacterial Mechanism of Mannosylerythritol Lipid-A and Lactic Acid on Listeria monocytogenes Based on Transcriptomic Analysis. Foods. 2022; 11(17):2660. https://doi.org/10.3390/foods11172660
Chicago/Turabian StyleLiu, Xiayu, Xinxin Pang, Yansha Wu, Yajing Wu, Ying Shi, Xinglin Zhang, and Qihe Chen. 2022. "Synergistic Antibacterial Mechanism of Mannosylerythritol Lipid-A and Lactic Acid on Listeria monocytogenes Based on Transcriptomic Analysis" Foods 11, no. 17: 2660. https://doi.org/10.3390/foods11172660