
Polysaccharides from Bamboo Shoot (Leleba oldhami Nakal) Byproducts Alleviate Antibiotic-Associated Diarrhea in Mice through Their Interactions with Gut Microbiota
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Extraction of the Polysaccharides
2.3. Chemical Component Analysis
2.4. Determination of Molecular Weight of BSP
2.5. Animal Experimental Design
2.6. Analysis of Mice Bodyweight and Fecal pH
2.7. Analysis of Short Chain Fatty Acids (SCFAs)
2.8. Histologic Observation of Colon and Small Intestine
2.9. Total DNA Extraction and Illumina High Throughput Sequencing Analysis
2.10. Bio-Informatics Analysis
2.11. Statistical Analysis
3. Results
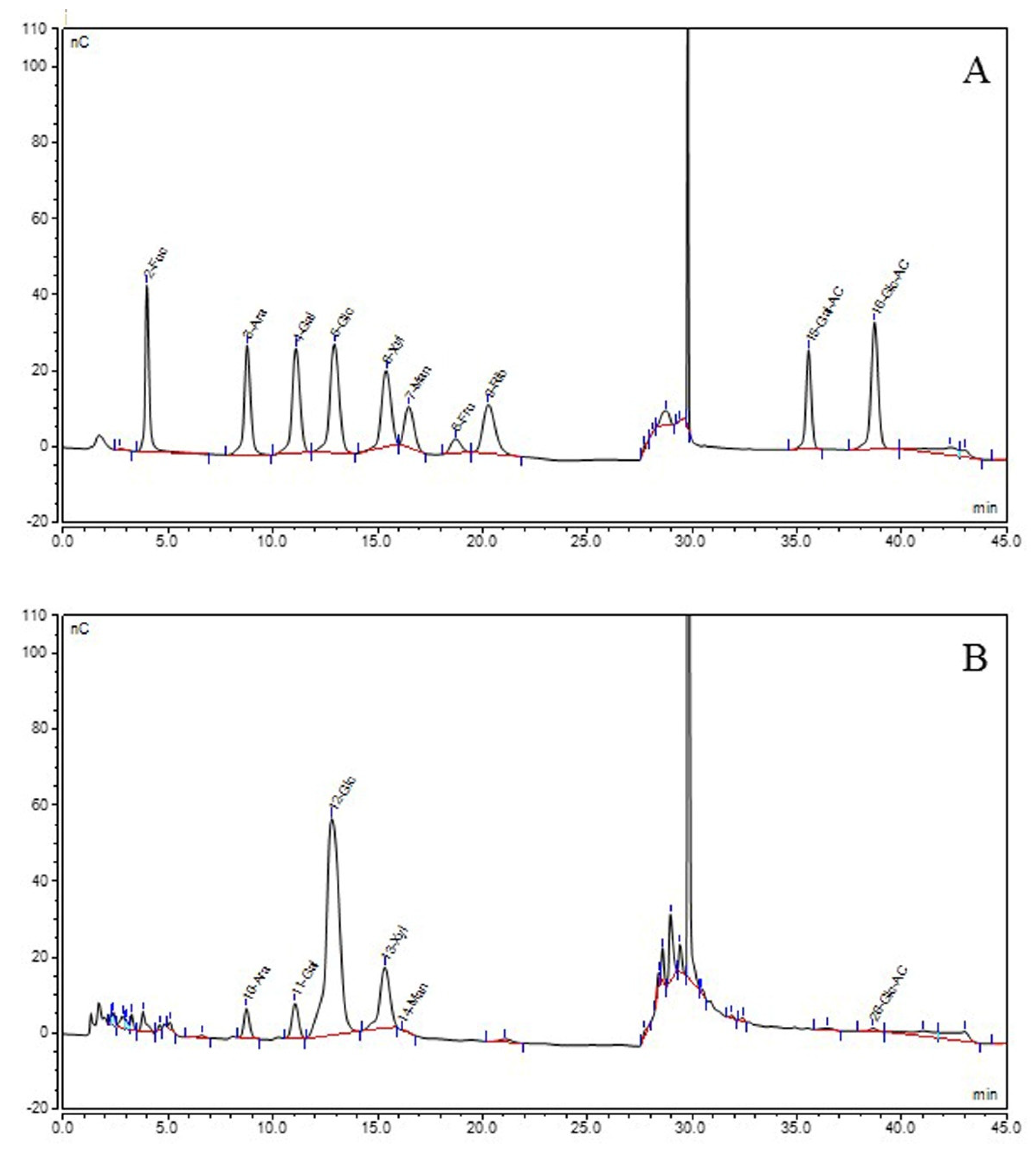
3.1. Composition and Molecular Weight of BSP
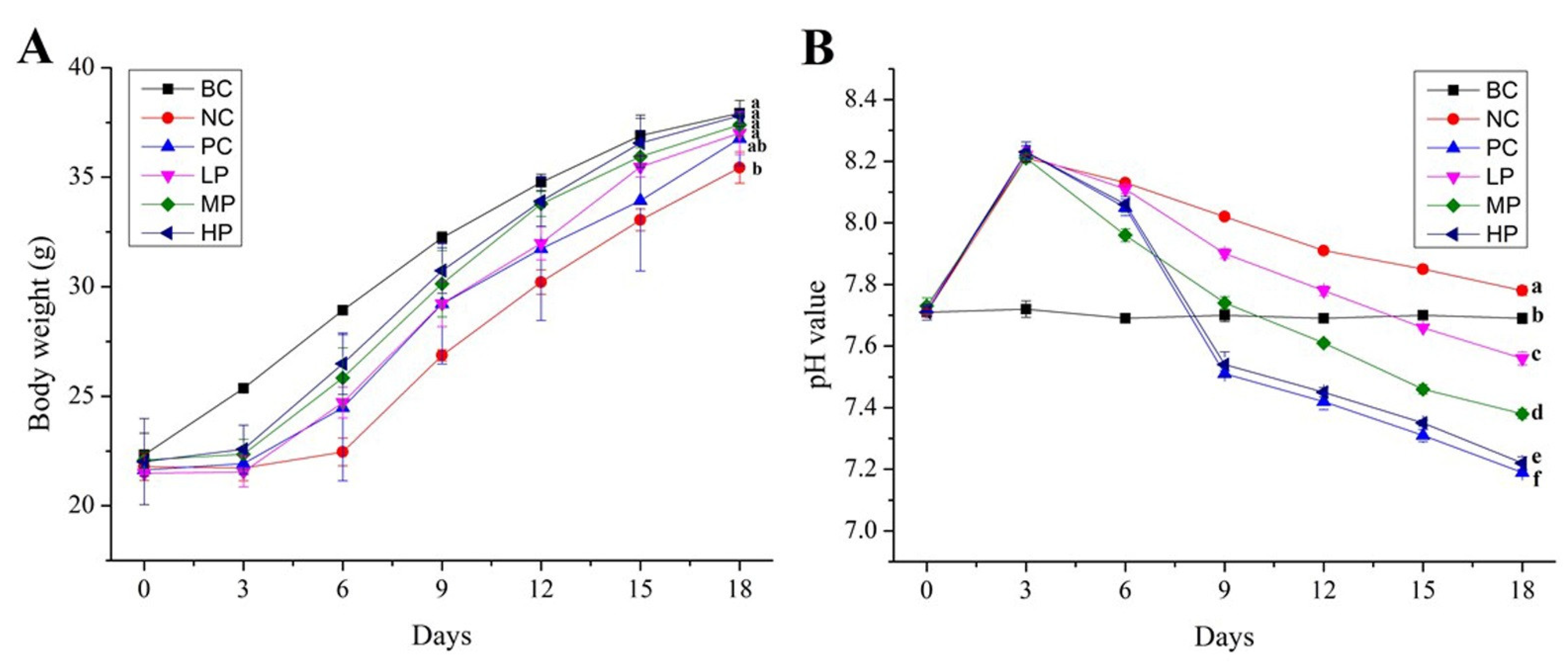
3.2. Effect of BSP on Bodyweight of Mice
3.3. Effect of BSP on pH Value and SCFAs of Mice Feces
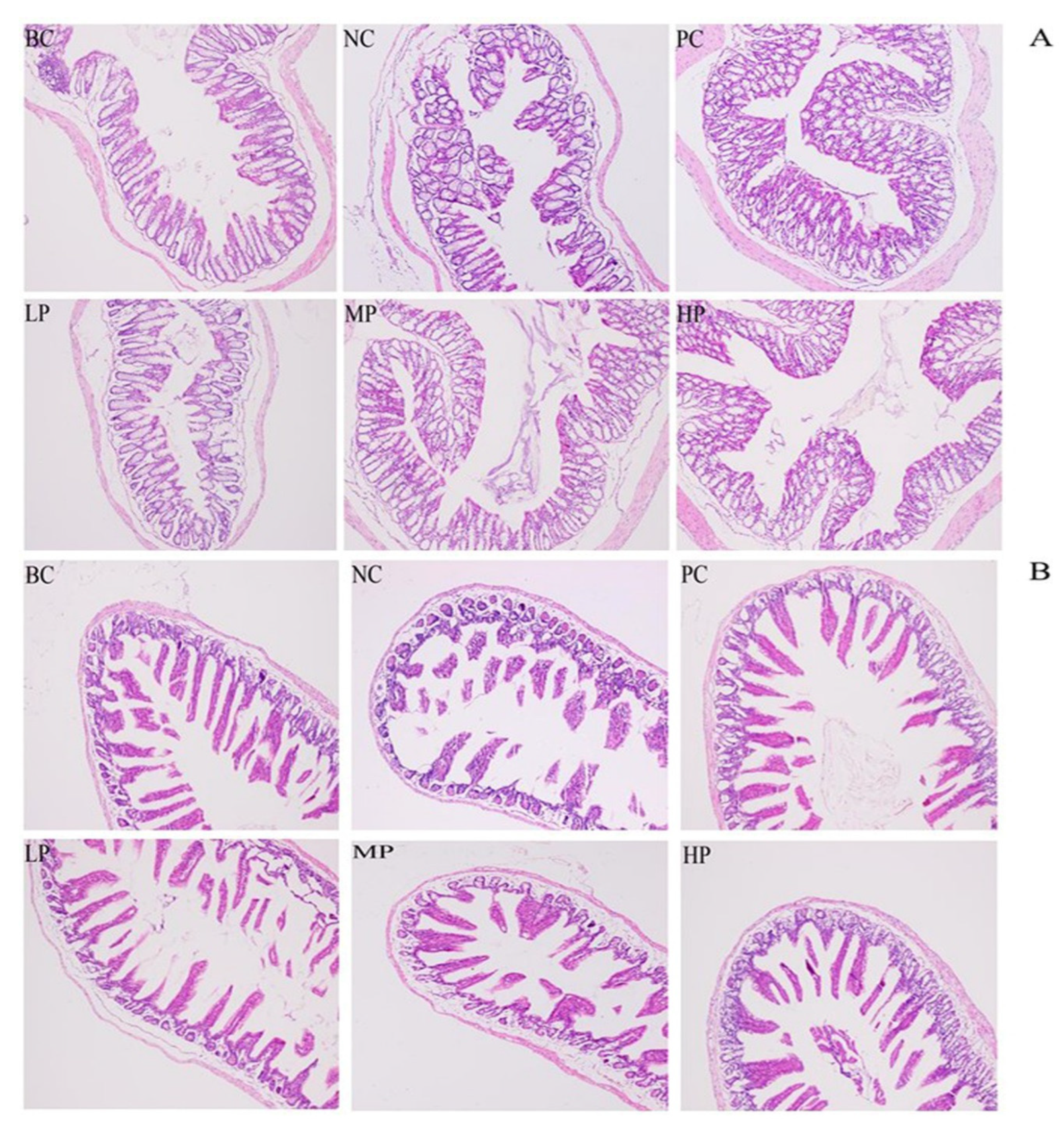
3.4. Histologic Observation of Colon and Small Intestine
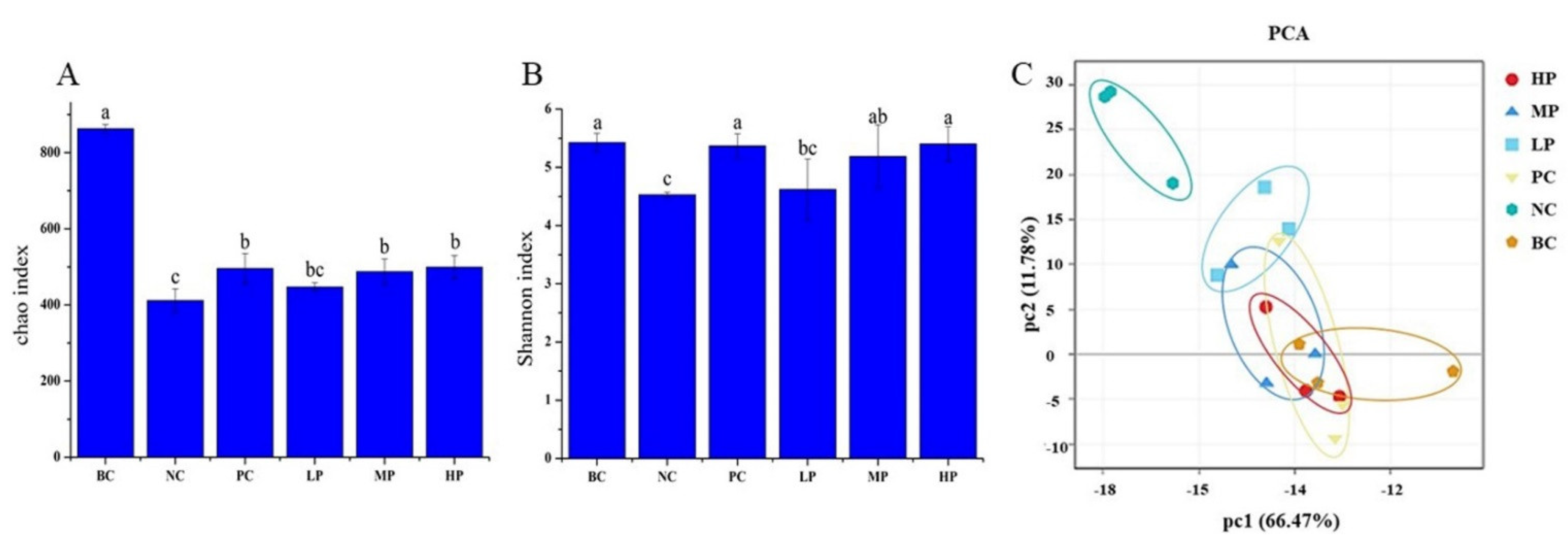
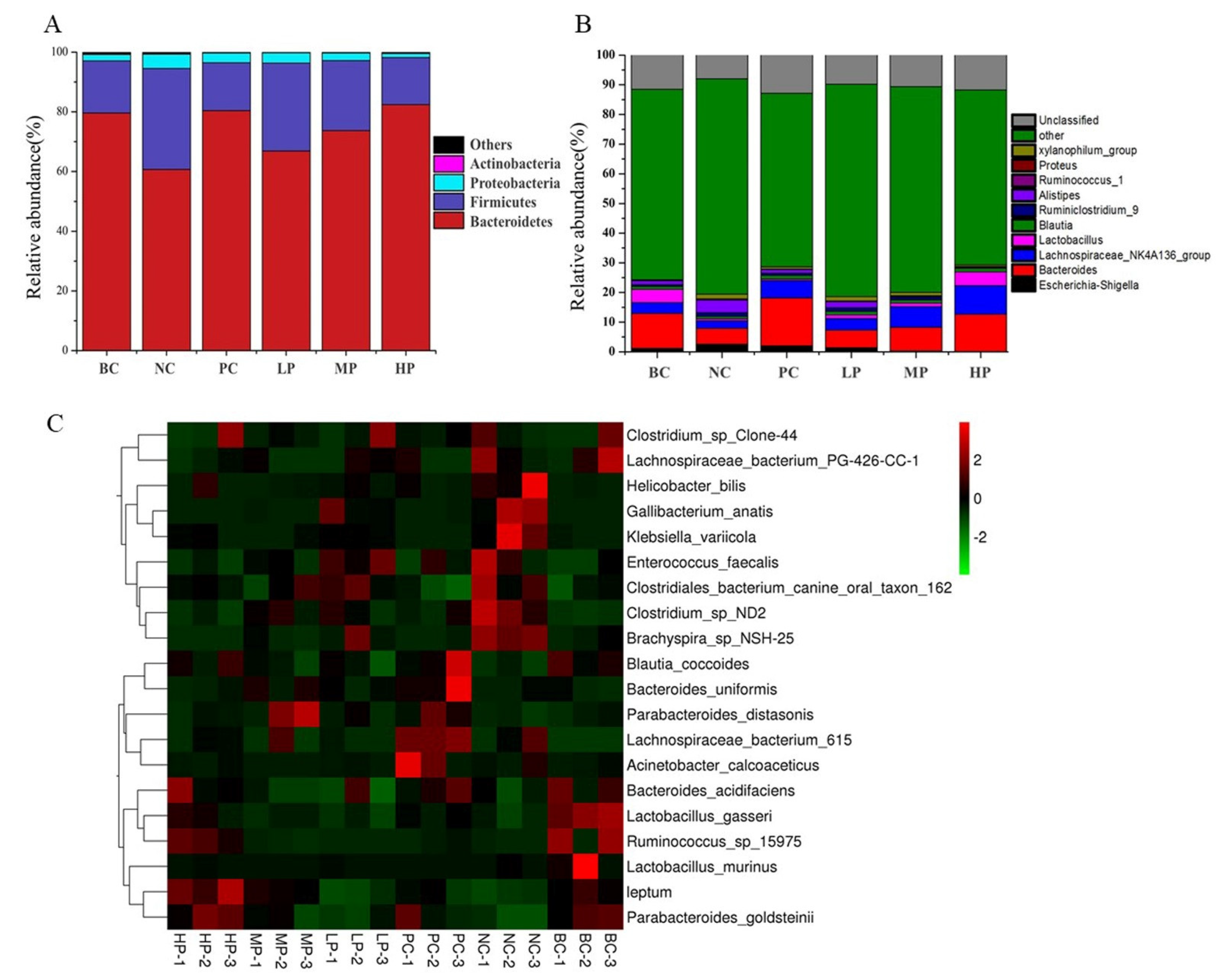
3.5. Overall Structural Characteristics of the Mice Microbiome
3.6. Composition Analysis of the Gut Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, X.; Guo, J.; Ji, K.; Zhang, P. Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota. Sci. Rep. 2016, 6, 32953. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Zhang, J.; Li, S.; Zheng, B.; Hu, J. Polysaccharides isolated from laminaria japonica attenuates gestational diabetes mellitus by regulating the gut microbiota in mice. Food Front. 2021, 2, 208–217. [Google Scholar] [CrossRef]
- Lowry, C.A.; Smith, D.G.; Siebler, P.H.; Schmidt, D.; Stamper, C.E.; Hassell, J.E.; Yamashita, P.S.; Fox, J.H.; Reber, S.O.; Brenner, L.A.; et al. The microbiota, immunoregulation, and mental health: Implications for public health. Curr. Environ. Health Rep. 2016, 3, 270–286. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, A.; Butt, M.S.; Afzaal, M.; van Baak, M.; Nadeem, M.T.; Shahid, M.Z. Prebiotics, gut microbiota and metabolic risks: Unveiling the relationship. J. Funct. Foods 2015, 17, 189–201. [Google Scholar] [CrossRef]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar] [CrossRef]
- Jakobsdottir, G.; Xu, J.; Molin, G.; Ahrne, S.; Nyman, M. High-fat diet reduces the formation of butyrate, but increases succinate, inflammation, liver fat and cholesterol in rats, while dietary fibre counteracts these effects. PLoS ONE 2013, 8, e80476. [Google Scholar] [CrossRef]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.J.; Wareham, K.; Wang, D.; Bradley, C.; Sewell, B.; Hutchings, H.; Harris, W.; Dhar, A.; Brown, H.; Foden, A.; et al. A high-dose preparation of lactobacilli and bifidobacteria in the prevention of antibiotic-associated and clostridium difficile diarrhoea in older people admitted to hospital: A multicentre, randomised, double-blind, placebo-controlled, parallel arm trial (placide). Health Technol. Assess 2013, 17, 1–140. [Google Scholar]
- Lv, W.; Liu, C.; Ye, C.; Sun, J.; Tan, X.; Zhang, C.; Qu, Q.; Shi, D.; Guo, S. Structural modulation of gut microbiota during alleviation of antibiotic-associated diarrhea with herbal formula. Int. J. Biol. Macromol. 2017, 105, 1622–1629. [Google Scholar] [CrossRef]
- Surawicz, C.M. Probiotics, antibiotic-associated diarrhoea and clostridium difficile diarrhoea in humans. Best Pract. Res. Clin. Gastroenterol. 2003, 17, 775–783. [Google Scholar] [CrossRef]
- Hamaker, B.R.; Tuncil, Y.E. A perspective on the complexity of dietary fiber structures and their potential effect on the gut microbiota. J. Mol. Biol. 2014, 426, 3838–3850. [Google Scholar] [CrossRef] [PubMed]
- Azmi, A.F.; Mustafa, S.; Hashim, D.M.; Manap, Y.A. Prebiotic activity of polysaccharides extracted from gigantochloa levis (buluh beting) shoots. Molecules 2012, 17, 1635–1651. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Chen, L.; Gao, K.; Shao, Z.; Huo, X.; Hua, M.; Liu, S.; Sun, Y.; Li, S. Effects of schisandra chinensis polysaccharides on rats with antibiotic-associated diarrhea. Int. J. Biol. Macromol. 2019, 124, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Qi, Y.; Chen, L.; Qu, D.; Li, Z.; Gao, K.; Chen, J.; Sun, Y. Effects of panax ginseng polysaccharides on the gut microbiota in mice with antibiotic-associated diarrhea. Int. J. Biol. Macromol. 2019, 124, 931–937. [Google Scholar] [CrossRef] [PubMed]
- He, M.X.; Wang, J.L.; Qin, H.; Shui, Z.X.; Zhu, Q.L.; Wu, B.; Tan, F.R.; Pan, K.; Hu, Q.C.; Dai, L.C.; et al. Bamboo: A new source of carbohydrate for biorefinery. Carbohydr. Polym. 2014, 111, 645–654. [Google Scholar] [CrossRef]
- Yang, Q.; Duan, Z.-B.; Wang, Z.-L.; He, K.-H.; Sun, Q.-X.; Peng, Z.-H. Bamboo resources, utilization and ex-situ conservation in xishuangbanna, south-eastern china. J. For. Res. 2008, 19, 79–83. [Google Scholar] [CrossRef]
- Chen, G.; Li, C.; Wang, S.; Mei, X.; Zhang, H.; Kan, J. Characterization of physicochemical properties and antioxidant activity of polysaccharides from shoot residues of bamboo (chimonobambusa quadrangularis): Effect of drying procedures. Food Chem. 2019, 292, 281–293. [Google Scholar] [CrossRef]
- Park, E.J.; Jhon, D.Y. Effects of bamboo shoot consumption on lipid profiles and bowel function in healthy young women. Nutrition 2009, 25, 723–728. [Google Scholar] [CrossRef]
- Choudhury, D.; Sahu, J.K.; Sharma, G.D. Value addition to bamboo shoots: A review. J. Food Sci. Technol. 2012, 49, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Satya, S.; Bal, L.M.; Singhal, P.; Naik, S.N. Bamboo shoot processing: Food quality and safety aspect (a review). Trends Food Sci. Technol. 2010, 21, 181–189. [Google Scholar] [CrossRef]
- Chen, G.; Bu, F.; Chen, X.; Li, C.; Wang, S.; Kan, J. Ultrasonic extraction, structural characterization, physicochemical properties and antioxidant activities of polysaccharides from bamboo shoots (chimonobambusa quadrangularis) processing by-products. Int. J. Biol. Macromol. 2018, 112, 656–666. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Wang, X.; Zhang, Y.; Wang, J.; Sun, H.; Wang, J.; Cao, X.; Ye, Y. Isolation and prebiotic activity of water-soluble polysaccharides fractions from the bamboo shoots (phyllostachys praecox). Carbohydr. Polym. 2016, 151, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Chen, X.; Yang, B.; Yu, Q.; Wei, X.; Ding, Y.; Kan, J. New insight into bamboo shoot (chimonobambusa quadrangularis) polysaccharides: Impact of extraction processes on its prebiotic activity. Food Hydrocoll. 2019, 95, 367–377. [Google Scholar] [CrossRef]
- Guo, Z.; Zhao, B.; Li, H.; Miao, S.; Zheng, B. Optimization of ultrasound-microwave synergistic extraction of prebiotic oligosaccharides from sweet potatoes (Ipomoea batatas L.). Innov. Food Sci. Emerg. Technol. 2019, 54, 51–63. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, S.; Wang, Q.; Lu, X.; Lin, L.; Tian, Y.; Xiao, J.; Zheng, B. Characterization and hypoglycemic activity of a β-pyran polysaccharides from bamboo shoot (leleba oldhami nakal) shells. Carbohydr. Polym. 2016, 144, 438–446. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Kemp, P.F.; Aller, J.Y. Bacterial diversity in aquatic and other environments: What 16s rdna libraries can tell us. FEMS Microbiol. Ecol. 2004, 47, 161–177. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, X.; Yu, S.; Zhao, M. Isolation and antioxidant activities of polysaccharides extracted from the shoots of phyllostachys edulis (carr.). Int. J. Biol. Macromol. 2011, 49, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Fung, K.Y.C.; Cosgrove, L.; Lockett, T.; Head, R.; Topping, D.L. A review of the potential mechanisms for the lowering of colorectal oncogenesis by butyrate. Br. J. Nutr. 2012, 108, 820–831. [Google Scholar] [CrossRef]
- Lu, S.; Zuo, T.; Zhang, N.; Shi, H.; Liu, F.; Wu, J.; Wang, Y.; Xue, C.; Tang, Q.-J. High throughput sequencing analysis reveals amelioration of intestinal dysbiosis by squid ink polysaccharide. J. Funct. Foods 2016, 20, 506–515. [Google Scholar] [CrossRef]
- Guo, T.; Song, D.; Ho, C.T.; Zhang, X.; Zhang, C.; Cao, J.; Wu, Z. Omics analyses of gut microbiota in a circadian rhythm disorder mouse model fed with oolong tea polyphenols. J. Agric. Food Chem. 2019, 67, 8847–8854. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-induced extinctions in the gut microbiota compound over generations. Nature 2016, 529, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Boutard, M.; Cerisy, T.; Nogue, P.Y.; Alberti, A.; Weissenbach, J.; Salanoubat, M.; Tolonen, A.C. Functional diversity of carbohydrate-active enzymes enabling a bacterium to ferment plant biomass. PLoS Genet 2014, 10, e1004773. [Google Scholar] [CrossRef]
- MoezArdalan, K.; Zali, M.R.; Dallal, M.M.S.; Hemami, M.R.; Salmanzadeh-Ahrabi, S. Prevalence and pattern of antimicrobial resistance of shigella species among patients with acute diarrhoea in karaj, tehran, iran. J. Health Popul. Nutr. 2003, 21, 96–102. [Google Scholar]
- Bird, A.R.; Vuaran, M.; Crittenden, R.; Hayakawa, T.; Playne, M.J.; Brown, I.L.; Topping, D.L. Comparative effects of a high-amylose starch and a fructooligosaccharide on fecal bifidobacteria numbers and short-chain fatty acids in pigs fed bifidobacterium animalis. Dig. Dis. Sci. 2009, 54, 947–954. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Acetic Acid | Propionic Acid | Butyric Acid | Valeric Acid |
|---|---|---|---|---|
| BC | 94.78 ± 3.06 c | 11.69 ± 0.47 c | 8.70 ± 0.33 e | 0.98 ± 0.05 c |
| NC | 84.23 ± 1.92 d | 7.78 ± 0.37 d | 6.29 ± 0.10 f | 0.75 ± 0.07 d |
| PC | 109.67 ± 3.17 a | 14.12 ± 0.34 a | 11.35 ± 0.38 b | 1.13 ± 0.03 b |
| LP | 96.57 ± 3.57 c | 11.31 ± 0.30 c | 9.09 ± 0.23 d | 0.98 ± 0.03 c |
| MP | 101.90 ± 2.24 b | 12.32 ± 0.14 b | 9.60 ± 0.12 c | 1.14 ± 0.04 b |
| HP | 107.27 ± 1.40 a | 13.60 ± 0.22 a | 12.50 ± 0.17 a | 1.63 ± 0.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Guan, X.; Liu, X.; Zhuang, W.; Xiao, Y.; Zheng, Y.; Wang, Q. Polysaccharides from Bamboo Shoot (Leleba oldhami Nakal) Byproducts Alleviate Antibiotic-Associated Diarrhea in Mice through Their Interactions with Gut Microbiota. Foods 2022, 11, 2647. https://doi.org/10.3390/foods11172647
Chen C, Guan X, Liu X, Zhuang W, Xiao Y, Zheng Y, Wang Q. Polysaccharides from Bamboo Shoot (Leleba oldhami Nakal) Byproducts Alleviate Antibiotic-Associated Diarrhea in Mice through Their Interactions with Gut Microbiota. Foods. 2022; 11(17):2647. https://doi.org/10.3390/foods11172647
Chicago/Turabian StyleChen, Canhui, Xuefang Guan, Xiaoyan Liu, Weijing Zhuang, Yiqian Xiao, Yafeng Zheng, and Qi Wang. 2022. "Polysaccharides from Bamboo Shoot (Leleba oldhami Nakal) Byproducts Alleviate Antibiotic-Associated Diarrhea in Mice through Their Interactions with Gut Microbiota" Foods 11, no. 17: 2647. https://doi.org/10.3390/foods11172647