
Understanding the Effect of Ozone on Listeria monocytogenes and Resident Microbiota of Gorgonzola Cheese Surface: A Culturomic Approach

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gorgonzola Rind Samples
2.2. Inoculum Preparation
2.3. Inoculation of Rind Samples and Ozone Treatments
2.4. Microbiological Analysis and Determination of pH/aw
2.5. MALDI-TOF MS Identification and Culturomic Analysis
2.6. Statistical Analyses and Graphing
3. Results
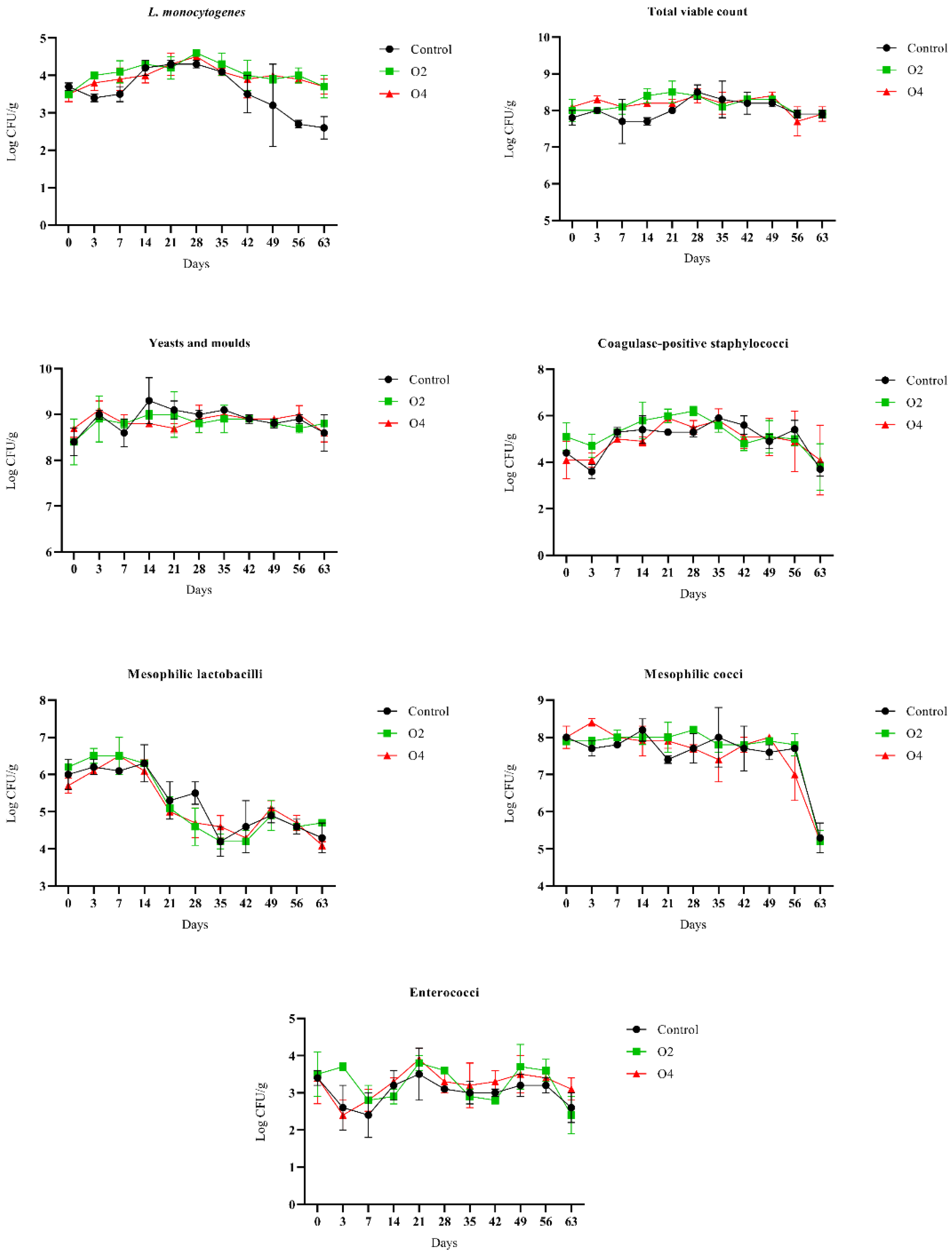
3.1. Behaviour of L. monocytogenes and Resident Populations during Storage
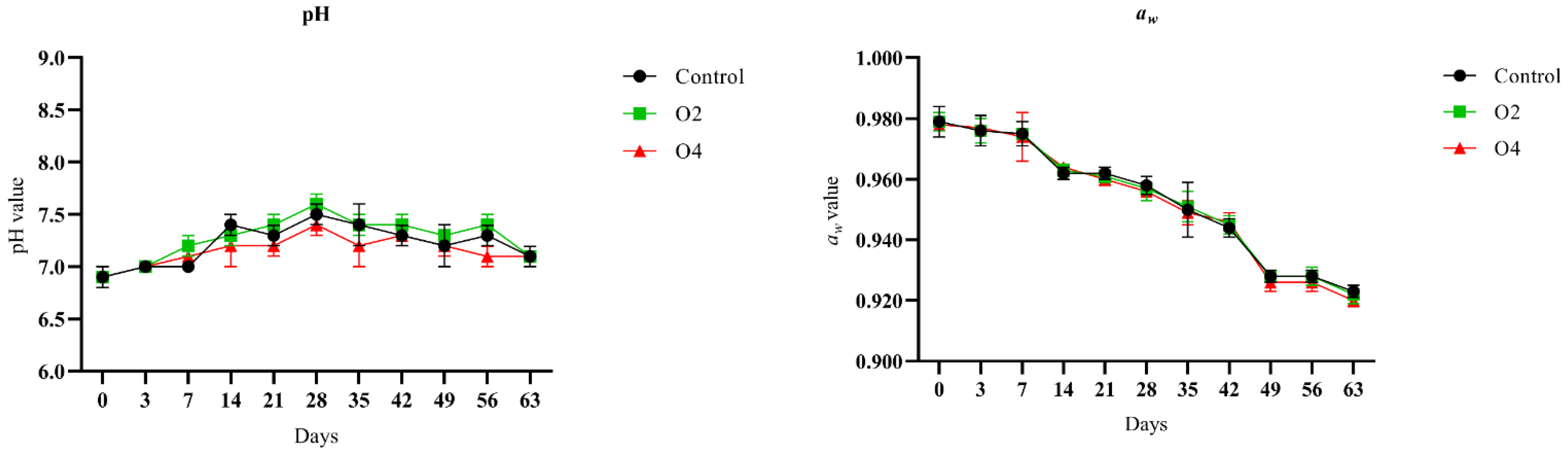
3.2. Changes in pH and aw during Storage
3.3. Culturomic Analysis and MALDI-TOF MS Identification
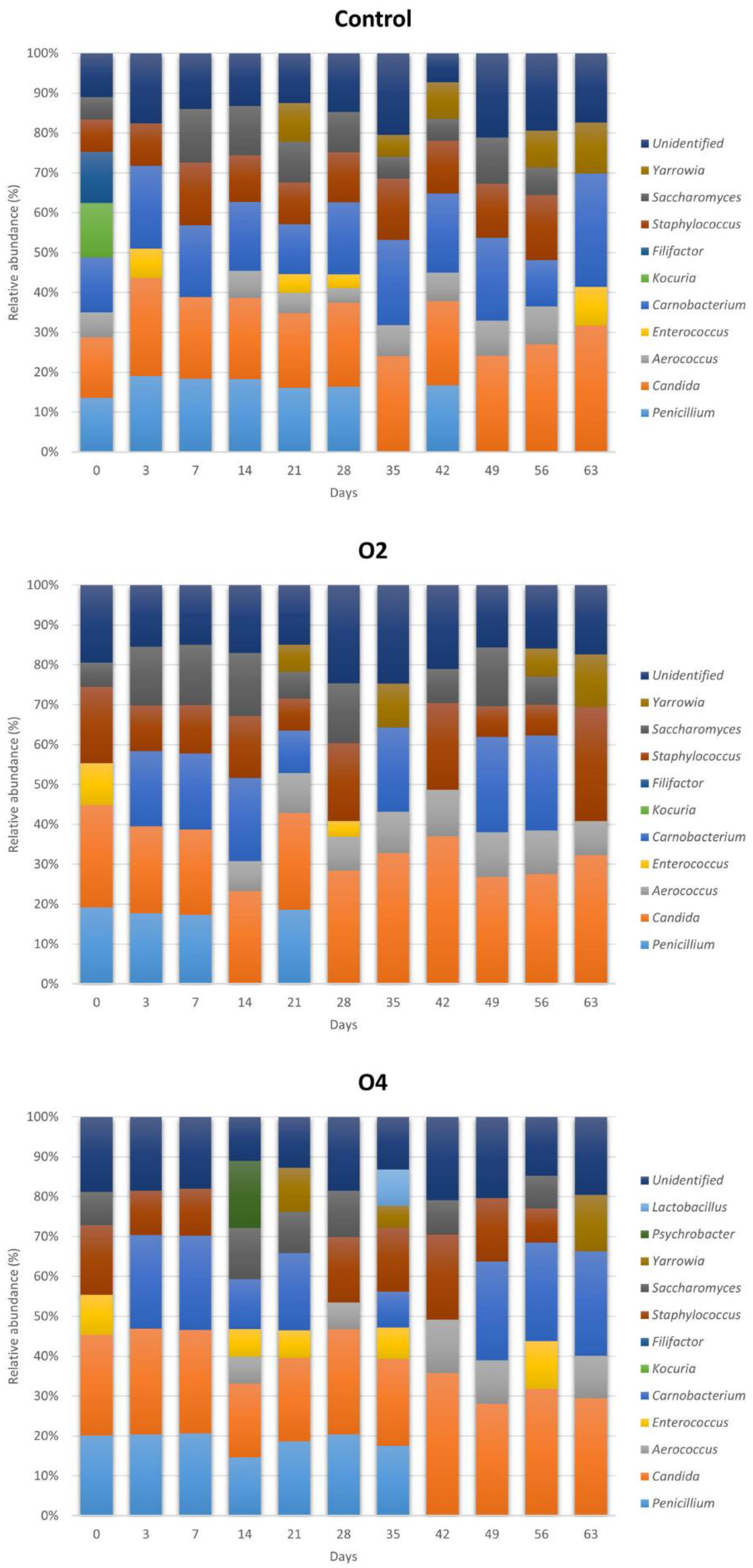
3.3.1. Relative Abundances of Microbial Genera
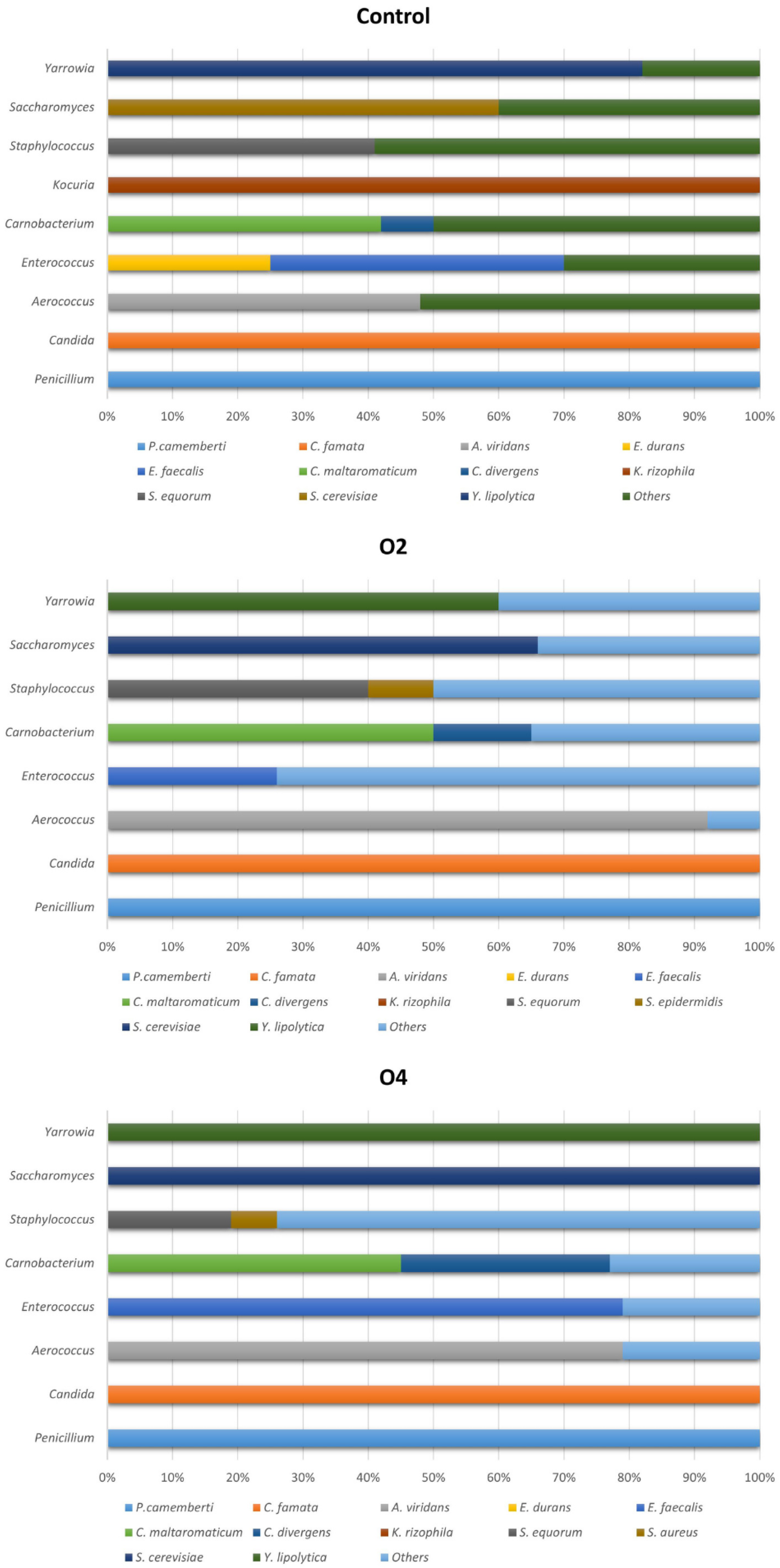
3.3.2. Percentage of Colonies Identified at Species Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority (EFSA) and European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- European Commission (EC). Commission Regulation (EC) No. 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. Eur. Union 2005, L338, 1–26. [Google Scholar]
- Di Ciccio, P.; Rubiola, S.; Panebianco, F.; Lomonaco, S.; Allard, M.; Bianchi, D.M.; Civera, T.; Chiesa, F. Biofilm Formation and Genomic Features of Listeria Monocytogenes Strains Isolated from Meat and Dairy Industries Located in Piedmont (Italy). Int. J. Food Microbiol. 2022, 378, 109784. [Google Scholar] [CrossRef]
- Martinez-Rios, V.; Dalgaard, P. Prevalence of Listeria Monocytogenes in European Cheeses: A Systematic Review and Meta-Analysis. Food Control 2018, 84, 205–214. [Google Scholar] [CrossRef]
- Di Ciccio, P.; Rubiola, S.; Grassi, M.A.; Civera, T.; Abbate, F.; Chiesa, F. Fate of Listeria Monocytogenes in the Presence of Resident Cheese Microbiota on Common Packaging Materials. Front. Microbiol. 2020, 11, 1526. [Google Scholar] [CrossRef]
- Foxx, P.F.; Guineee, T.P.; Cogann, T.M.; Mcsweeney, P.L.H. Fundamentals of Cheese Science, 2nd ed.; Springer: New York, NY, USA, 2017. [Google Scholar]
- Morandi, S.; Silvetti, T.; Vezzini, V.; Morozzo, E.; Brasca, M. How We Can Improve the Antimicrobial Performances of Lactic Acid Bacteria? A New Strategy to Control Listeria Monocytogenes in Gorgonzola Cheese. Food Microbiol. 2020, 90, 103488. [Google Scholar] [CrossRef]
- Lomonaco, S.; Decastelli, L.; Nucera, D.; Gallina, S.; Manila Bianchi, D.; Civera, T. Listeria Monocytogenes in Gorgonzola: Subtypes, Diversity and Persistence over Time. Int. J. Food Microbiol. 2009, 128, 516–520. [Google Scholar] [CrossRef]
- Carminati, D.; Perrone, A.; Giraffa, G.; Neviani, E.; Mucchetti, G. Characterization of Listeria Monocytogenes Strains Isolated from Gorgonzola Cheese Rinds. Food Microbiol. 2004, 21, 801–807. [Google Scholar] [CrossRef]
- Manfreda, G.; De Cesare, A.; Stella, S.; Cozzi, M.; Cantoni, C. Occurrence and Ribotypes of Listeria Monocytogenes in Gorgonzola Cheeses. Int. J. Food Microbiol. 2005, 102, 287–293. [Google Scholar] [CrossRef]
- Bernini, V.; Dalzini, E.; Lazzi, C.; Bottari, B.; Gatti, M.; Neviani, E. Cutting Procedures Might Be Responsible for Listeria Monocytogenes Contamination of Foods: The Case of Gorgonzola Cheese. Food Control 2016, 61, 54–61. [Google Scholar] [CrossRef]
- Morandi, S.; Silvetti, T.; Battelli, G.; Brasca, M. Can Lactic Acid Bacteria Be an Efficient Tool for Controlling Listeria Monocytogenes Contamination on Cheese Surface? The Case of Gorgonzola Cheese. Food Control 2019, 96, 499–507. [Google Scholar] [CrossRef]
- Bernini, V.; Dalzini, E.; Lazzi, C.; Bottari, B.; Bisotti, S.; Fontana, M.; Neviani, E. A Multi-Sampling Approach to Evaluate an Infrared Surface Treatment for Reducing Listeria Monocytogenes Contamination on Whole Gorgonzola Cheese Rinds. Food Control 2015, 55, 75–81. [Google Scholar] [CrossRef]
- Carminati, D.; Gatti, M.; Bonvini, B.; Neviani, E.; Mucchetti, G. High-Pressure Processing of Gorgonzola Cheese: Influence on Listeria Monocytogenes Inactivation and on Sensory Characteristics. J. Food Prot. 2004, 67, 1671–1675. [Google Scholar] [CrossRef]
- Mucchetti, G.; Bonvini, B.; Francolino, S.; Neviani, E.; Carminati, D. Effect of Washing with a High Pressure Water Spray on Removal of Listeria Innocua from Gorgonzola Cheese Rind. Food Control 2008, 19, 521–525. [Google Scholar] [CrossRef]
- Masotti, F.; Vallone, L.; Ranzini, S.; Silvetti, T.; Morandi, S.; Brasca, M. Effectiveness of Air Disinfection by Ozonation or Hydrogen Peroxide Aerosolization in Dairy Environments. Food Control 2019, 97, 32–38. [Google Scholar] [CrossRef]
- Sivaranjani, S.; Prasath, V.A.; Pandiselvam, R.; Kothakota, A.; Mousavi Khaneghah, A. Recent Advances in Applications of Ozone in the Cereal Industry. LWT 2021, 146, 111412. [Google Scholar] [CrossRef]
- Botta, C.; Ferrocino, I.; Pessione, A.; Cocolin, L.; Rantsiou, K. Spatiotemporal Distribution of the Environmental Microbiota in Food Processing Plants as Impacted by Cleaning and Sanitizing Procedures: The Case of Slaughterhouses and Gaseous Ozone. Appl. Environ. Microbiol. 2020, 86, e01861-20. [Google Scholar] [CrossRef]
- Panebianco, F.; Rubiola, S.; Di Ciccio, P.A. The Use of Ozone as an Eco-Friendly Strategy against Microbial Biofilm in Dairy Manufacturing Plants: A Review. Microorganisms 2022, 10, 162. [Google Scholar] [CrossRef]
- Panebianco, F.; Rubiola, S.; Chiesa, F.; Civera, T.; Di Ciccio, P.A. Effect of Gaseous Ozone on Listeria Monocytogenes Planktonic Cells and Biofilm: An in Vitro Study. Foods 2021, 10, 1484. [Google Scholar] [CrossRef]
- Marino, M.; Maifreni, M.; Baggio, A.; Innocente, N. Inactivation of Foodborne Bacteria Biofilms by Aqueous and Gaseous Ozone. Front. Microbiol. 2018, 9, 02024. [Google Scholar] [CrossRef]
- Bigi, F.; Haghighi, H.; Quartieri, A.; de Leo, R.; Pulvirenti, A. Impact of Low-Dose Gaseous Ozone Treatment to Reduce the Growth of in Vitro Broth Cultures of Foodborne Pathogenic/Spoilage Bacteria in a Food Storage Cold Chamber. J. Food Saf. 2021, 41, e12892. [Google Scholar] [CrossRef]
- De Candia, S.; Morea, M.; Baruzzi, F. Eradication of High Viable Loads of Listeria Monocytogenes Contaminating Food-Contact Surfaces. Front. Microbiol. 2015, 6, 733. [Google Scholar] [CrossRef]
- Harada, A.M.M.; Nascimento, M.S. Efficacy of Dry Sanitizing Methods on Listeria Monocytogenes Biofilms. Food Control 2021, 124, 107897. [Google Scholar] [CrossRef]
- Giménez, B.; Graiver, N.; Giannuzzi, L.; Zaritzky, N. Treatment of Beef with Gaseous Ozone: Physicochemical Aspects and Antimicrobial Effects on Heterotrophic Microflora and Listeria Monocytogenes. Food Control 2021, 121, 107602. [Google Scholar] [CrossRef]
- Baggio, A.; Marino, M.; Innocente, N.; Celotto, M.; Maifreni, M. Antimicrobial Effect of Oxidative Technologies in Food Processing: An Overview. Eur. Food Res. Technol. 2020, 246, 669–692. [Google Scholar] [CrossRef]
- Panebianco, F.; Rubiola, S.; Chiesa, F.; Civera, T.; Di Ciccio, P.A. Effect of Gaseous Ozone Treatment on Biofilm of Dairy-Isolated Pseudomonas Spp. Strains. Ital. J. Food Saf. 2022, 11, 10350. [Google Scholar] [CrossRef]
- Lagier, J.C.; Dubourg, G.; Million, M.; Cadoret, F.; Bilen, M.; Fenollar, F.; Levasseur, A.; Rolain, J.M.; Fournier, P.E.; Raoult, D. Culturing the Human Microbiota and Culturomics. Nat. Rev. Microbiol. 2018, 16, 540–550. [Google Scholar] [CrossRef]
- Ziino, G.; Marotta, S.M.; Giarratana, F.; Giuffrida, A.; Panebianco, F. Reliability Evaluation of MALDI-TOF MS Associated with SARAMIS Software in Rapid Identification of Thermophilic Campylobacter Isolated from Food. Food Anal. Methods 2019, 12, 1128–1132. [Google Scholar] [CrossRef]
- Huber, I.; Pavlovic, M.; Maggipinto, M.; Konrad, R.; Busch, U. Interlaboratory Proficiency Test Using MALDI-TOF MS for Identification of Food-Associated Bacteria. Food Anal. Methods 2018, 11, 1068–1075. [Google Scholar] [CrossRef]
- Ryu, S.; Park, W.S.; Yun, B.; Shin, M.; Go, G.W.; Kim, J.N.; Oh, S.; Kim, Y. Diversity and Characteristics of Raw Milk Microbiota from Korean Dairy Farms Using Metagenomic and Culturomic Analysis. Food Control 2021, 127, 108160. [Google Scholar] [CrossRef]
- Höll, L.; Behr, J.; Vogel, R.F. Identification and Growth Dynamics of Meat Spoilage Microorganisms in Modified Atmosphere Packaged Poultry Meat by MALDI-TOF MS. Food Microbiol. 2016, 60, 84–91. [Google Scholar] [CrossRef]
- Peruzy, M.F.; Murru, N.; Yu, Z.; Kerkhof, P.J.; Neola, B.; Joossens, M.; Proroga, Y.T.R.; Houf, K. Assessment of Microbial Communities on Freshly Killed Wild Boar Meat by MALDI-TOF MS and 16S RRNA Amplicon Sequencing. Int. J. Food Microbiol. 2019, 301, 51–60. [Google Scholar] [CrossRef]
- Altakhis, M.; Pillidge, C.J.; Osborn, A.M.; Torley, P.J.; Kaur, M. Assessment of the Potential Use of MALDI-TOF MS for the Identification of Bacteria Associated with Chilled Vacuum-Packaged Lamb Meat. Meat Sci. 2021, 177, 108508. [Google Scholar] [CrossRef]
- Lappa, I.K.; Gantzias, C.; Manolopoulou, E.; De Brandt, E.; Aerts, M.; Vandamme, P.; Tsakalidou, E.; Georgalaki, M. MALDI-TOF MS Insight into the Biodiversity of Staka, the Artisanal Cretan Soured Cream. Int. Dairy J. 2021, 116, 104969. [Google Scholar] [CrossRef]
- Cocolin, L.; Nucera, D.; Alessandria, V.; Rantsiou, K.; Dolci, P.; Grassi, M.A.; Lomonaco, S.; Civera, T. Microbial Ecology of Gorgonzola Rinds and Occurrence of Different Biotypes of Listeria Monocytogenes. Int. J. Food Microbiol. 2009, 133, 200–205. [Google Scholar] [CrossRef]
- Fontana, C.; Cappa, F.; Rebecchi, A.; Cocconcelli, P.S. Surface Microbiota Analysis of Taleggio, Gorgonzola, Casera, Scimudin and Formaggio Di Fossa Italian Cheeses. Int. J. Food Microbiol. 2010, 138, 205–211. [Google Scholar] [CrossRef]
- Gobbetti, M.; Burzigotti, R.; Smacchi, E.; Corsetti, A.; De Angelis, M. Microbiology and biochemistry of Gorgonzola cheese during ripening. Int. Dairy J. 1997, 7, 519–529. [Google Scholar] [CrossRef]
- Morandi, S.; Brasca, M.; Battelli, G. Impiego di ozono per il controllo di Listeria monocytogenes in diverse tipologie di formaggio. Sci. E Tec. Latt. Casearia 2009, 60, 211–215. [Google Scholar]
- Robbins, J.B.; Fisher, C.W.; Moltz, A.G.; Martin, S.E. Elimination of Listeria monocytogenes biofilms by ozone, chlorine, and hydrogen peroxide. J. Food Prot. 2005, 68, 494–498. [Google Scholar] [CrossRef]
- Wani, S.; Maker, J.K.; Thompson, J.R.; Barnes, J.; Singleton, I. Effect of Ozone Treatment on Inactivation of Escherichia Coli and Listeria Sp. on Spinach. Agriculture 2015, 5, 155–169. [Google Scholar] [CrossRef]
- Pascual, A.; Llorca, I.; Canut, A. Use of Ozone in Food Industries for Reducing the Environmental Impact of Cleaning and Disinfection Activities. Trends Food Sci. Technol. 2007, 18, S29–S35. [Google Scholar] [CrossRef]
- Sørensen, J.S.; Bøknæs, N.; Mejlholm, O.; Dalgaard, P. Superchilling in Combination with Modified Atmosphere Packaging Resulted in Long Shelf-Life and Limited Microbial Growth in Atlantic Cod (Gadus Morhua L.) from Capture-Based-Aquaculture in Greenland. Food Microbiol. 2020, 88, 103405. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.S.; Chang, Y.R.; Tsai, Y.T.; Yang, Y.T.; Weng, S.H.; Tseng, L.F.; Chou, H.C.; Hu, A.T.; Liao, E.C.; Chen, H.Y.; et al. The Distribution of Cultivable Oral Anaerobic Microbiota Identified by MALDI-TOF MS in Healthy Subjects and in Patients with Periodontal Disease. J. Pharm. Biomed. Anal. 2021, 192, 113647. [Google Scholar] [CrossRef] [PubMed]
- Muthukumar, A.; Muthuchamy, M. Optimization of Ozone in Gaseous Phase to Inactivate Listeria Monocytogenes on Raw Chicken Samples. Food Res. Int. 2013, 54, 1128–1130. [Google Scholar] [CrossRef]
- Campagnollo, F.B.; Margalho, L.P.; Kamimura, B.A.; Feliciano, M.D.; Freire, L.; Lopes, L.S.; Alvarenga, V.O.; Cadavez, V.A.P.; Gonzales-Barron, U.; Schaffner, D.W.; et al. Selection of Indigenous Lactic Acid Bacteria Presenting Anti-Listerial Activity, and Their Role in Reducing the Maturation Period and Assuring the Safety of Traditional Brazilian Cheeses. Food Microbiol. 2018, 73, 288–297. [Google Scholar] [CrossRef]
- Østergaard, N.B.; Eklöw, A.; Dalgaard, P. Modelling the Effect of Lactic Acid Bacteria from Starter- and Aroma Culture on Growth of Listeria Monocytogenes in Cottage Cheese. Int. J. Food Microbiol. 2014, 188, 15–25. [Google Scholar] [CrossRef]
- Panebianco, F.; Giarratana, F.; Caridi, A.; Sidari, R.; De Bruno, A.; Giuffrida, A. Lactic Acid Bacteria Isolated from Traditional Italian Dairy Products: Activity against Listeria Monocytogenes and Modelling of Microbial Competition in Soft Cheese. LWT 2021, 137, 110446. [Google Scholar] [CrossRef]
- Salomskiene, J.; Jonkuviene, D.; Macioniene, I.; Abraitiene, A.; Zeime, J.; Repeckiene, J.; Vaiciulyte-Funk, L. Differences in the Occurence and Efficiency of Antimicrobial Compounds Produced by Lactic Acid Bacteria. Eur. Food Res. Technol. 2019, 245, 569–579. [Google Scholar] [CrossRef]
- Cadavez, V.A.P.; Campagnollo, F.B.; Silva, R.A.; Duffner, C.M.; Schaffner, D.W.; Sant’Ana, A.S.; Gonzales-Barron, U. A Comparison of Dynamic Tertiary and Competition Models for Describing the Fate of Listeria Monocytogenes in Minas Fresh Cheese during Refrigerated Storage. Food Microbiol. 2019, 79, 48–60. [Google Scholar] [CrossRef]
- Costa, J.C.C.P.; Bover-Cid, S.; Bolívar, A.; Zurera, G.; Pérez-Rodríguez, F. Modelling the Interaction of the Sakacin-Producing Lactobacillus Sakei CTC494 and Listeria Monocytogenes in Filleted Gilthead Sea Bream (Sparus Aurata) under Modified Atmosphere Packaging at Isothermal and Non-Isothermal Conditions. Int. J. Food Microbiol. 2019, 297, 72–84. [Google Scholar] [CrossRef]
- Ross, T.; Dalgaard, P.; Tienungoon, S. Predictive Modelling of the Growth and Survival of Listeria in Fishery Products. Int. J. Food Microbiol. 2000, 62, 231–245. [Google Scholar] [CrossRef]
- Giménez, B.; Dalgaard, P. Modelling and Predicting the Simultaneous Growth of Listeria Monocytogenes and Spoilage Micro-Organisms in Cold-Smoked Salmon. J. Appl. Microbiol. 2004, 96, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Naito, S.; Takahara, H. Ozone Contribution in Food Industry in Japan. Ozone Sci. Eng. 2006, 28, 425–429. [Google Scholar] [CrossRef]
- Manousaridis, G.; Nerantzaki, A.; Paleologos, E.K.; Tsiotsias, A.; Savvaidis, I.N.; Kontominas, M.G. Effect of Ozone on Microbial, Chemical and Sensory Attributes of Shucked Mussels. Food Microbiol. 2005, 22, 1–9. [Google Scholar] [CrossRef]
- Segat, A.; Biasutti, M.; Iacumin, L.; Comi, G.; Baruzzi, F.; Carboni, C.; Innocente, N. Use of Ozone in Production Chain of High Moisture Mozzarella Cheese. LWT Food Sci. Technol. 2014, 55, 513–520. [Google Scholar] [CrossRef]
- Werlang, G.O.; Kich, J.D.; Lopes, G.V.; Coldebella, A.; Feddern, V.; Cardoso, M. Effect of Gaseous Ozone Application during Chilling on Microbial and Quality Attributes of Pig Carcasses. Food Sci. Technol. Int. 2022, 28, 366–376. [Google Scholar] [CrossRef]
- Correa, F.T.; de Souza, A.C.; de Souza Júnior, E.A.; Isidoro, S.R.; Piccoli, R.H.; Dias, D.R.; de Abreu, L.R. Effect of Brazilian Green Propolis on Microorganism Contaminants of Surface of Gorgonzola-Type Cheese. J. Food Sci. Technol. 2019, 56, 1978–1987. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Internal ID | NCBI ID a | Isolation Source | Year | Lineage | Serogroup | Sequence Type | Clonal Complex |
|---|---|---|---|---|---|---|---|
| G40 | CFSAN044840 | Gorgonzola cheese | 2005 | II | IIa | 325 | 31 |
| G52 | CFSAN044807 | Gorgonzola cheese | 2004 | II | Iia | 325 | 31 |
| G70 | CFSAN044814 | Gorgonzola cheese | 2004 | II | Iia | 325 | 31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panebianco, F.; Rubiola, S.; Buttieri, C.; Di Ciccio, P.A.; Chiesa, F.; Civera, T. Understanding the Effect of Ozone on Listeria monocytogenes and Resident Microbiota of Gorgonzola Cheese Surface: A Culturomic Approach. Foods 2022, 11, 2640. https://doi.org/10.3390/foods11172640
Panebianco F, Rubiola S, Buttieri C, Di Ciccio PA, Chiesa F, Civera T. Understanding the Effect of Ozone on Listeria monocytogenes and Resident Microbiota of Gorgonzola Cheese Surface: A Culturomic Approach. Foods. 2022; 11(17):2640. https://doi.org/10.3390/foods11172640
Chicago/Turabian StylePanebianco, Felice, Selene Rubiola, Chiara Buttieri, Pierluigi Aldo Di Ciccio, Francesco Chiesa, and Tiziana Civera. 2022. "Understanding the Effect of Ozone on Listeria monocytogenes and Resident Microbiota of Gorgonzola Cheese Surface: A Culturomic Approach" Foods 11, no. 17: 2640. https://doi.org/10.3390/foods11172640