
Isolation and Identification of Lactococcus lactis and Weissella cibaria Strains from Fermented Beetroot and an Investigation of Their Properties as Potential Starter Cultures and Probiotics

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Microorganisms
2.2. Identification Using the MALDI Technique
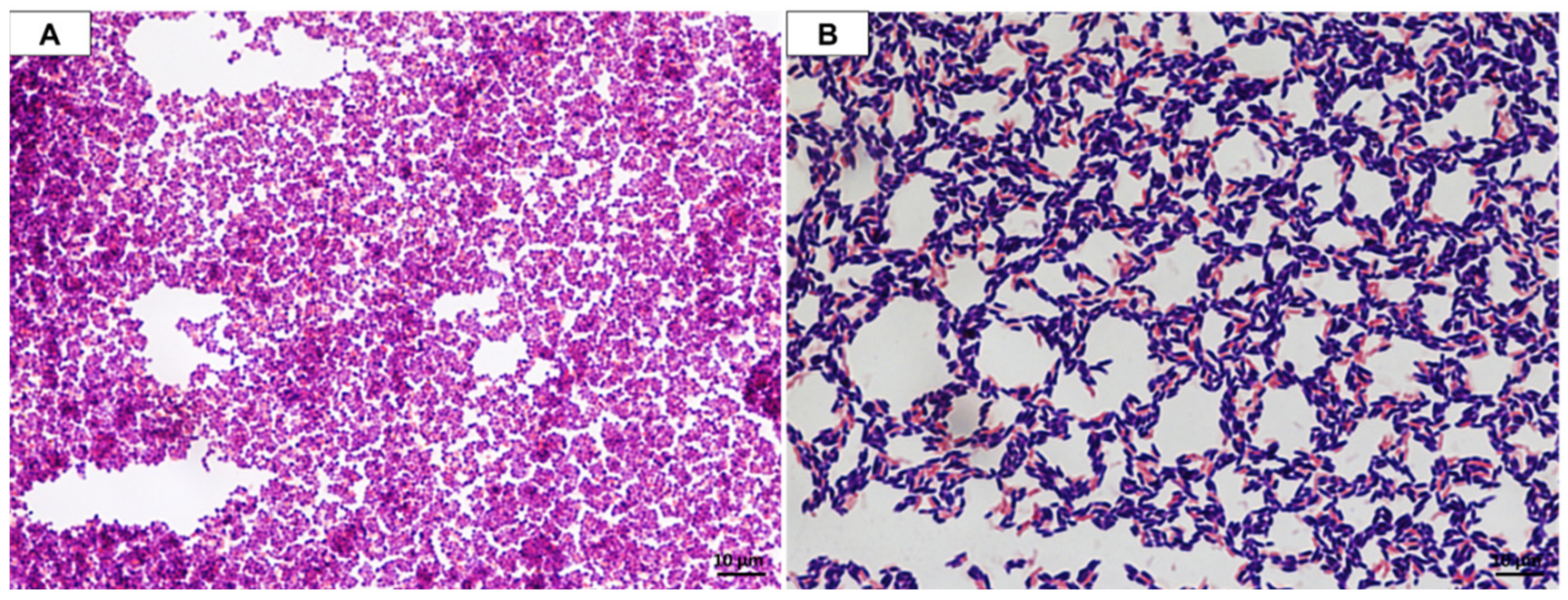
2.3. Microscopic Observation
2.4. Confirmation of Identification by 16S rRNA Gene Sequencing
2.5. Growth Assessment of Selected Isolates
2.6. Determination of Proteolytic Properties
2.7. Volatile Organic Compounds Analysis
2.8. Analysis of Fats by Mass Spectrometry
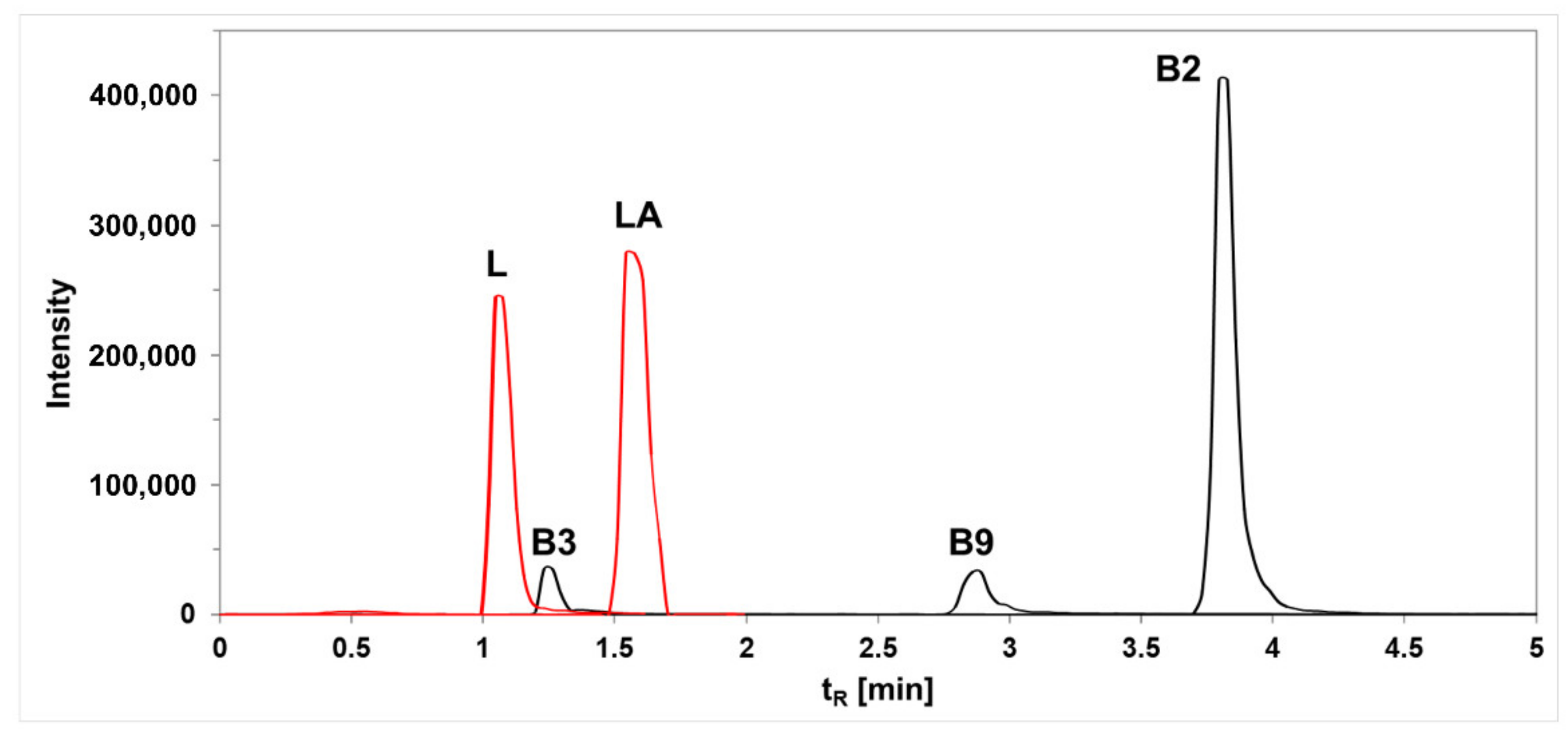
2.9. Analysis of B Vitamins and Lactic Acid
2.10. Inhibition of Pathogens
3. Results and Discussion
3.1. Isolation of Lactic Acid Bacteria—Identification and Phenotypic Characterization
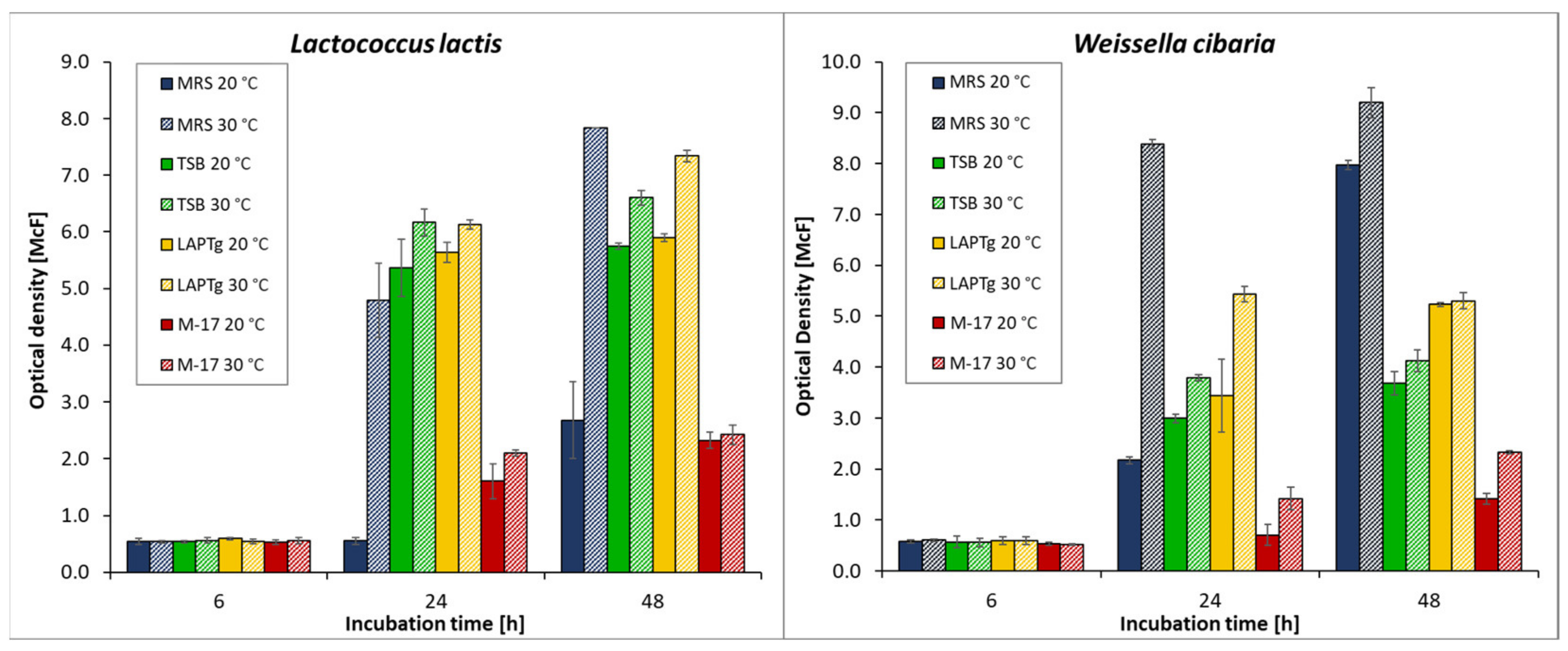
3.2. Growth Assessment of Selected Isolates
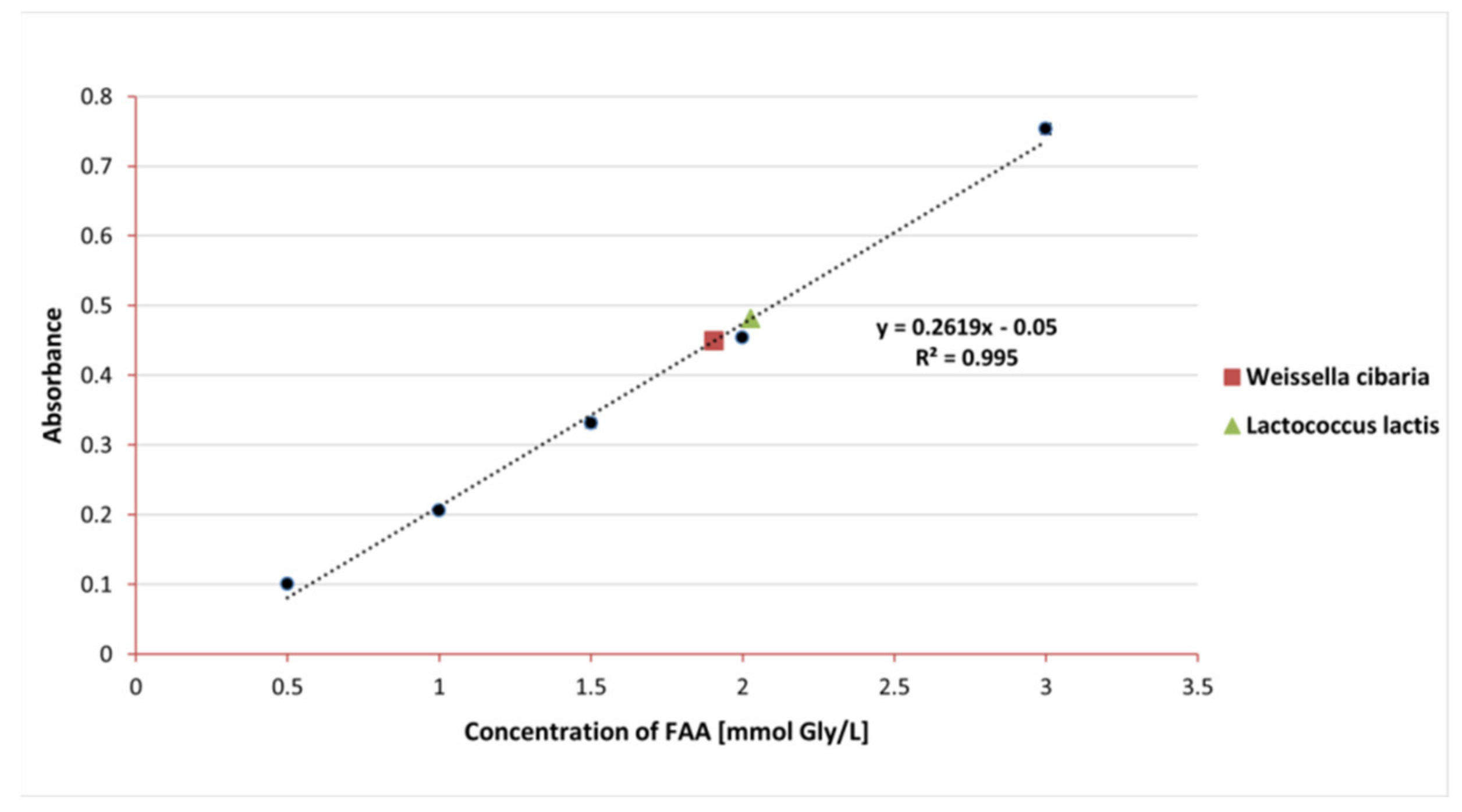
3.3. Proteolytic Properties
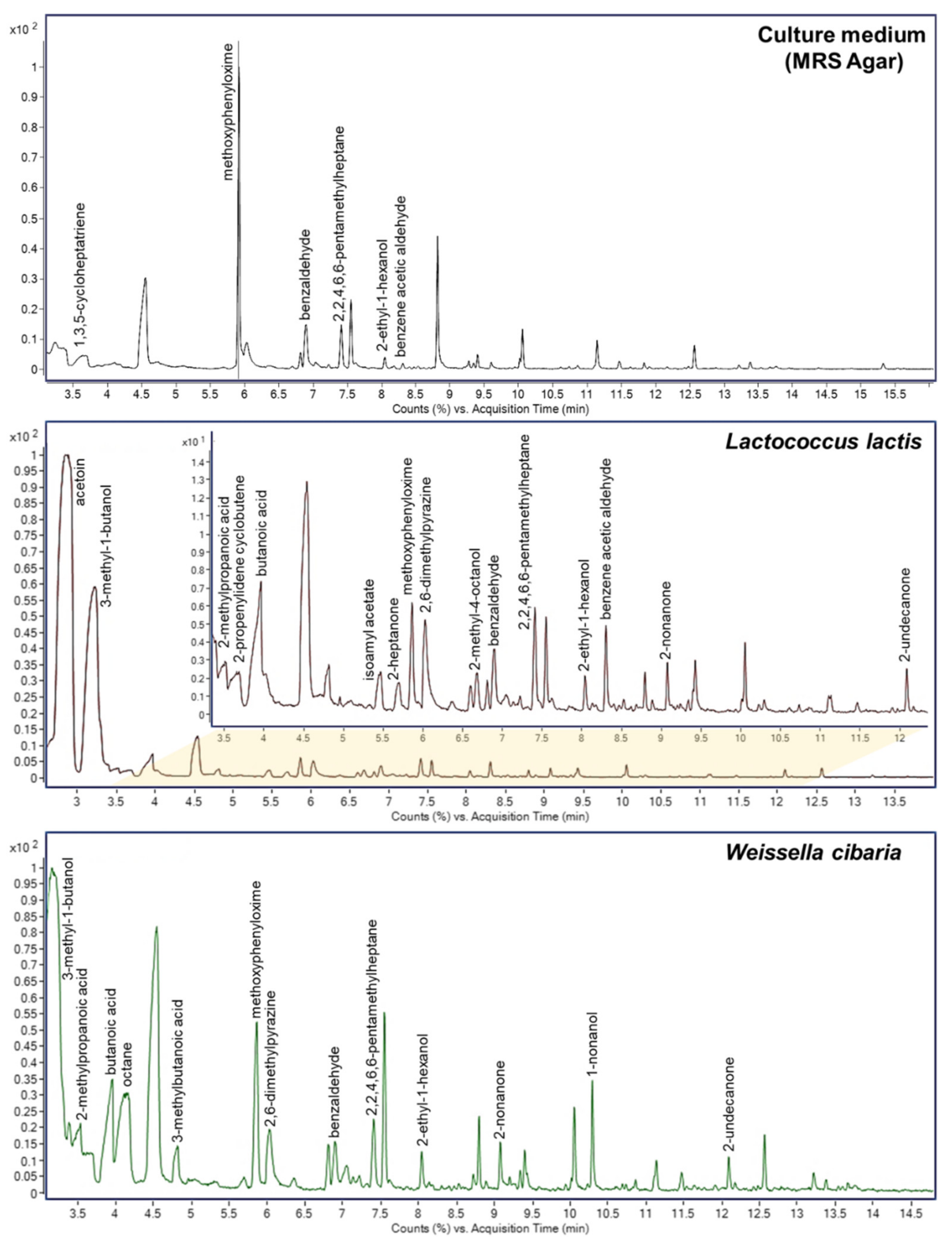
3.4. Volatile Compounds
3.5. Analysis of Fats
3.6. Synthesis of Vitamins and Lactic Acid
3.7. Study of Antagonistic Abilities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO/WHO. Guidelines for the Evaluation of Probiotics in Food; FAO: Rome, Italy, 2002; pp. 1–11. [Google Scholar]
- Santacroce, L.; Charitos, I.A.; Bottalico, L. A successful history: Probiotics and their potential as antimicrobials. Expert Rev. Anti-Infect. Ther. 2019, 17, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Fenster, K.; Freeburg, B.; Hollard, C.; Wong, C.; Laursen, R.R.; Ouwehand, A.C. The production and delivery of probiotics: A review of a practical approach. Microorganisms 2019, 7, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worku, H.; Amenu, K.; Fesseha, H. The Role of Starter Culture in Producing Probiotic Yoghurt: Significance for Human Health—A Review. Int. J. Recent Biotechnol. 2020, 8, 20–34. [Google Scholar]
- Kumar, B.V.; Vijayendra, S.V.N.; Reddy, O.V.S. Trends in dairy and non-dairy probiotic products—A review. J. Food Sci. Technol. 2015, 52, 6112–6124. [Google Scholar] [CrossRef] [Green Version]
- Khaneghah, A.M.; Abhari, K.; Eş, I.; Soares, M.B.; Oliveira, R.B.A.; Hosseini, H.; Rezaei, M.; Balthazar, C.F.; Silva, R.; Cruz, A.G.; et al. Interactions between probiotics and pathogenic microorganisms in hosts and foods: A review. Trends Food Sci. Technol. 2020, 95, 205–218. [Google Scholar] [CrossRef]
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; el Enshasy, H. Lactic acid bacteria: From starter cultures to producers of chemicals. FEMS Microbiol. Lett. 2018, 365, fny213. [Google Scholar] [CrossRef] [Green Version]
- Abedi, E.; Hashemi, S.M.B. Lactic acid production—Producing microorganisms and substrates sources-state of art. Heliyon 2020, 6, e04974. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, L.; Wen, R.; Chen, Q.; Kong, B. Role of lactic acid bacteria in flavor development in traditional Chinese fermented foods: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 2741–2755. [Google Scholar] [CrossRef]
- Walczak-Skierska, J.; Złoch, M.; Pauter, K.; Pomastowski, P.; Buszewski, B. Lipidomic analysis of lactic acid bacteria strains by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. J. Dairy Sci. 2020, 103, 11062–11078. [Google Scholar] [CrossRef]
- Nagpal, R.; Kumar, A.; Kumar, M.; Behare, P.V.; Jain, S.; Yadav, H. Probiotics, their health benefits and applications for developing healthier foods: A review. FEMS Microbiol. Lett. 2012, 334, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kerry, R.G.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.S.; Das, G. Benefaction of probiotics for human health: A review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, R.S.D.; Vieira-Coelho, M.A. Probiotics and prebiotics: Focus on psychiatric disorders- A systematic review. Nutr. Rev. 2020, 78, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Curtis, N. The influence of the intestinal microbiome on vaccine responses. Vaccine 2018, 36, 4433–4439. [Google Scholar] [CrossRef] [PubMed]
- Shori, A.B. The potential applications of probiotics on dairy and non-dairy foods focusing on viability during storage. Biocatal. Agric. Biotechnol. 2015, 4, 423–431. [Google Scholar] [CrossRef]
- Bruker Daltonics. MALDI Biotyper 3.1 User Manual; Bruker Daltonics: Billerica, MA, USA, 2012; Volume 1, pp. 1–212. [Google Scholar]
- Pomastowski, P.; Złoch, M.; Rodzik, A.; Ligor, M.; Kostrzewa, M.; Buszewski, B. Analysis of bacteria associated with honeys of different geographical and botanical origin using two different identification approaches: MALDI-TOF MS and 16S rDNA PCR technique. PLoS ONE 2019, 14, e0217078. [Google Scholar] [CrossRef] [Green Version]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric Assay Using o-Phthaldialdehyde for Determination of Proteolysis in Milk and Isolated Milk Proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Arendowski, A.; Sagandykova, G.; Mametov, R.; Rafińska, K.; Pryshchepa, O.; Pomastowski, P. Nanostructured Layer of Silver for Detection of Small Biomolecules in Surface-Assisted Laser Desorption Ionization Mass Spectrometry. Materials 2022, 15, 4076. [Google Scholar] [CrossRef]
- These, H. Spoilage of Vegetables and Fruits. In Food Microbiology: Principles into Practice; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 337–363. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Medina, E.; Sánchez, B.; Benítez-Cabello, A.; Arroyo-López, F.N. Role of lactic acid bacteria in fermented vegetables. Grasas Aceites 2020, 71, e358. [Google Scholar] [CrossRef]
- Yu, H.S.; Lee, N.K.; Choi, A.J.; Choe, J.S.; Bae, C.H.; Paik, H.D. Anti-inflammatory potential of probiotic strain Weissella cibaria JW15 isolated from kimchi through regulation of NF-κB and MAPKs pathways in LPS-induced RAW 264.7 cells. J. Microbiol. Biotechnol. 2019, 29, 1022–1032. [Google Scholar] [CrossRef]
- Buszewski, B.; Rogowska, A.; Pomastowski, P.; Złoch, M.; Railean-Plugaru, V. Identification of microorganisms by modern analytical techniques. J. AOAC Int. 2017, 100, 1607–1623. [Google Scholar] [CrossRef]
- Model, M.A. Methods for cell volume measurement. Cytom. Part A 2018, 93, 281–296. [Google Scholar] [CrossRef] [Green Version]
- Sáez, G.D.; Flomenbaum, L.; Zárate, G. Lactic acid bacteria from argentinean femented foods: Isolation and characterization for their potential use as starters for fermentation of vegetables. Food Technol. Biotechnol. 2018, 56, 398–410. [Google Scholar] [CrossRef]
- Joković, N.; Rajković, J.; Veljović, K.; Tolina, M. Screening of lactic acid bacteria isolated from Serbian kajmak for use in starter cultures. Biol. Nyssana 2014, 5, 37–46. [Google Scholar]
- González, L.; Sacristán, N.; Arenas, R.; Fresno, J.M.; Tornadijo, M.E. Enzymatic activity of lactic acid bacteria (with antimicrobial properties) isolated from a traditional Spanish cheese. Food Microbiol. 2010, 27, 592–597. [Google Scholar] [CrossRef]
- Herreros, M.A.; Fresno, J.M.; Prieto, M.J.G.; Tornadijo, M.E. Technological characterization of lactic acid bacteria isolated from Armada cheese (a Spanish goats’ milk cheese). Int. Dairy J. 2003, 13, 469–479. [Google Scholar] [CrossRef]
- McSweeney, P.L.H.; Fox, P.F. Advanced Dairy Chemistry; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Liu, J.M.; Chen, L.; Dorau, R.; Lillevang, S.K.; Jensen, P.R.; Solem, C. From Waste to Taste—Efficient Production of the Butter Aroma Compound Acetoin from Low-Value Dairy Side Streams Using a Natural (Nonengineered) Lactococcus lactis Dairy Isolate. J. Agric. Food Chem. 2020, 68, 5891–5899. [Google Scholar] [CrossRef]
- Capozzi, V.; Lonzarich, V.; Khomenko, I.; Cappellin, L.; Navarini, L.; Biasioli, F. Unveiling the molecular basis of mascarpone cheese aroma: VOCs analysis by SPME-GC/MS and PTR-ToF-MS. Molecules 2020, 25, 1242. [Google Scholar] [CrossRef] [Green Version]
- Mallia, S.; Escher, F.; Schlichtherle-Cerny, H. Aroma-active compounds of butter: A review. Eur. Food Res. Technol. 2008, 226, 315–325. [Google Scholar] [CrossRef]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
- Fahy, E.; Sud, M.; Cotter, D.; Subramaniam, S. LIPID MAPS online tools for lipid research. Nucleic Acids Res. 2007, 35, W606–W612. [Google Scholar] [CrossRef] [Green Version]
- Caspi, R.; Billington, R.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Midford, P.E.; Ong, W.K.; Paley, S.; Subhraveti, P.; Karp, P.D. The MetaCyc database of metabolic pathways and enzymes-a 2019 update. Nucleic Acids Res. 2020, 48, D445–D453. [Google Scholar] [CrossRef] [Green Version]
- Arendowski, A.; Nizioł, J.; Ossoliński, K.; Ossolińska, A.; Ossoliński, T.; Dobrowolski, Z.; Ruman, T. Laser desorption/ionization MS imaging of cancer kidney tissue on silver nanoparticle-enhanced target. Bioanalysis 2018, 10, 83–94. [Google Scholar] [CrossRef]
- Nizioł, J.; Rode, W.; Zieliński, Z.; Ruman, T. Matrix-free laser desorption-ionization with silver nanoparticle-enhanced steel targets. Int. J. Mass Spectrom. 2013, 335, 22–32. [Google Scholar] [CrossRef]
- Cioffi, N.; Colaianni, L.; Pilolli, R.; Calvano, C.D.; Palmisano, F.; Zambonin, P.G. Silver nanofractals: Electrochemical synthesis, XPS characterization and application in LDI-MS. Anal. Bioanal. Chem. 2009, 394, 1375–1383. [Google Scholar] [CrossRef]
- Silina, Y.E.; Meier, F.; Nebolsin, V.A.; Koch, M.; Volmer, D.A. Novel galvanic nanostructures of Ag and Pd for efficient laser desorption/ionization of low molecular weight compounds. J. Am. Soc. Mass Spectrom. 2014, 25, 841–851. [Google Scholar] [CrossRef]
- Sekuła, J.; Nizioł, J.; Rode, W.; Ruman, T. Silver nanostructures in laser desorption/ionization mass spectrometry and mass spectrometry imaging. Analyst 2015, 140, 6195–6209. [Google Scholar] [CrossRef]
- Ulmer, C.Z.; Jones, C.M.; Yost, R.A.; Garrett, T.J.; Bowden, J.A. Optimization of Folch, Bligh-Dyer, and Matyash sample-to-extraction solvent ratios for human plasma-based lipidomics studies. Anal. Chim. Acta 2018, 1037, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Mierau, I.; Venema, G.; Kok, J.; Kunji, E.R.S. Casein and peptide degradation in lactic acid bacteria. Biotechnol. Genet. Eng. Rev. 1997, 14, 279–302. [Google Scholar] [CrossRef] [PubMed]
- Raveschot, C.; Cudennec, B.; Coutte, F.; Flahaut, C.; Fremont, M.; Drider, D.; Dhulster, P. Production of bioactive peptides by Lactobacillus species: From gene to application. Front. Microbiol. 2018, 9, 2354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliazucchi, D.; Martini, S.; Solieri, L. Bioprospecting for bioactive peptide production by lactic acid bacteria isolated from fermented dairy food. Fermentation 2019, 5, 96. [Google Scholar] [CrossRef] [Green Version]
- Doi, Y. Glycerol metabolism and its regulation in lactic acid bacteria. Appl. Microbiol. Biotechnol. 2019, 103, 5079–5093. [Google Scholar] [CrossRef]
- Gao, Y.; Li, D. Screening of lactic acid bacteria with cholesterol-lowering and triglyceride-lowering activity in vitro and evaluation of probiotic function. Ann. Microbiol. 2018, 68, 537–545. [Google Scholar] [CrossRef]
- Alvarez, H.M.; Steinbüchel, A. Triacylglycerols in prokaryotic microorganisms. Appl. Microbiol. Biotechnol. 2002, 60, 367–376. [Google Scholar] [CrossRef]
- López-Lara, I.M.; Geiger, O. Bacterial lipid diversity. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 1287–1299. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2015, 40, 133–159. [Google Scholar] [CrossRef] [Green Version]
- Kondakova, T.; D’Heygère, F.; Feuilloley, M.J.; Orange, N.; Heipieper, H.J.; Poc, C.D. Glycerophospholipid synthesis and functions in Pseudomonas. Chem. Phys. Lipids 2015, 190, 27–42. [Google Scholar] [CrossRef]
- Lew, L.-C.; Liong, M.-T. Bioactives from probiotics for dermal health: Functions and benefits. J. Appl. Microbiol. 2013, 114, 1241–1253. [Google Scholar] [CrossRef]
- Wei, J.H.; Yin, X.; Welander, P.V. Sterol Synthesis in Diverse Bacteria. Front. Microbiol. 2016, 7, 990. [Google Scholar] [CrossRef] [Green Version]
- Adsul, M.; Khire, J.; Bastawde, K.; Gokhale, D. Production of Lactic Acid from Cellobiose and Cellotriose by Lactobacillus delbrueckii Mutant Uc-3. Appl. Environ. Microbiol. 2007, 73, 5055–5057. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, J.G.; Laiño, J.E.; del Valle, M.J.; Vannini, V.; van Sinderen, D.; Taranto, M.P.; de Valdez, G.F.; de Giori, G.S.; Sesma, F. B-Group vitamin production by lactic acid bacteria—Current knowledge and potential applications. J. Appl. Microbiol. 2011, 111, 1297–1309. [Google Scholar] [CrossRef]
- Yépez, A.; Russo, P.; Spano, G.; Khomenko, I.; Biasioli, F.; Capozzi, V.; Aznar, R. In situ riboflavin fortification of different kefir-like cereal-based beverages using selected Andean LAB strains. Food Microbiol. 2019, 77, 61–68. [Google Scholar] [CrossRef]
- Ewe, J.; Liong, M. Viability and growth characteristics of Lactobacillus in soymilk supplemented with B-vitamins. Int. J. Food Sci. Nutr. 2010, 61, 87–107. [Google Scholar] [CrossRef]
- Thakur, K.; Lule, V.K.; Rajni, C.S.; Kumar, N.; Mandal, S.; Anand, S.; Kumari, V.; Tomar, S.K. Riboflavin Producing Probiotic Lactobacilli as a Biotechnological Strategy to Obtain Riboflavin-enriched Fermented Foods. J. Pure Appl. Microbiol. 2016, 10, 161–166. [Google Scholar]
- Jayashree, S.; Jayaraman, K.; Kalaichelvan, G. Isolation, Screening and Characterization of Riboflavin. Recent Res. Sci. Technol. 2010, 2, 83–88. [Google Scholar]
- Levit, R.; de Giori, G.S.; de LeBlanc, A.M.; LeBlanc, J.G. Recent update on lactic acid bacteria producing riboflavin and folates: Application for food fortification and treatment of intestinal inflammation. J. Appl. Microbiol. 2021, 130, 1412–1424. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Burgess, C.; Sesma, F.; de Giori, G.S.; van Sinderen, D. Lactococcus lactis is capable of improving the riboflavin status in deficient rats. Br. J. Nutr. 2005, 94, 262–267. [Google Scholar] [CrossRef] [Green Version]
- Burgess, C.; O’Connell-Motherway, M.; Sybesma, W.; Hugenholtz, J.; van Sinderen, D. Riboflavin Production in Lactococcus lactis: Potential for In Situ Production of Vitamin-Enriched Foods. Appl. Environ. Microbiol. 2004, 70, 5769–5777. [Google Scholar] [CrossRef] [Green Version]
- Zeisel, H.S.; Stover, P. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline1; National Agricultural Library: Baltimore, MD, USA, 1998; pp. 35–50. [Google Scholar]
- Jung, M.Y.; Lee, C.; Seo, M.J.; Roh, S.W.; Lee, S.H. Characterization of a potential probiotic bacterium Lactococcus raffinolactis WiKim0068 isolated from fermented vegetable using genomic and in vitro analyses. BMC Microbiol. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- Kang, M.-S.; Yeu, J.-E.; Hong, S.-P. Safety Evaluation of Oral Care Probiotics Weissella cibaria CMU and CMS1 by Phenotypic and Genotypic Analysis. Int. J. Mol. Sci. 2019, 20, 2693. [Google Scholar] [CrossRef] [Green Version]
- Divya, J.B.; Varsha, K.K.; Nampoothiri, K.M. Newly isolated lactic acid bacteria with probiotic features for potential application in food industry. Appl. Biochem. Biotechnol. 2012, 167, 1314–1324. [Google Scholar] [CrossRef]
- Månberger, A.; Verbrugghe, P.; Guðmundsdóttir, E.E.; Santesson, S.; Nilsson, A.; Hreggviðsson, G.Ó.; Linares-Pastén, J.A.; Karlsson, E.N. Taxogenomic assessment and genomic characterisation of Weissella cibaria strain 92 able to metabolise oligosaccharides derived from dietary fibres. Sci. Rep. 2020, 10, 5853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Bungenstock, L.; Abdulmawjood, A.; Reich, F. Evaluation of antibacterial properties of lactic acid bacteria from traditionally and industrially produced fermented sausages from Germany. PLoS ONE 2020, 15, e0230345. [Google Scholar] [CrossRef] [PubMed]
- Alegría, Á.; Delgado, S.; Roces, C.; López, B.; Mayo, B. Bacteriocins produced by wild Lactococcus lactis strains isolated from traditional, starter-free cheeses made of raw milk. Int. J. Food Microbiol. 2010, 143, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, A.; Suzuki, M. Antimicrobial activity of Lactococcus lactis subsp. lactis isolated from a stranded cuvier’s beaked whale (Ziphius cavirostris) against gram-positive and -negative bacteria. Microorganisms 2021, 9, 243. [Google Scholar] [CrossRef]
- Srionnual, S.; Yanagida, F.; Lin, L.H.; Hsiao, K.N.; Chen, Y.S. Weissellicin 110, a newly discovered bacteriocin from Weissella cibaria 110, isolated from plaa-som, a fermented fish product from Thailand. Appl. Environ. Microbiol. 2007, 73, 2247–2250. [Google Scholar] [CrossRef] [Green Version]
- Lakra, A.K.; Domdi, L.; Hanjon, G.; Tilwani, Y.M.; Arul, V. Some probiotic potential of Weissella confusa MD1 and Weissella cibaria MD2 isolated from fermented batter. LWT 2020, 125, 109261. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time (Min) | Ionization | Molecular Ion | Fragment Ion | Q1 (V) | CE | Q3 (V) | |
|---|---|---|---|---|---|---|---|
| B2 | 3.811 | Positive | 337 | 243 | −20 | −25 | −20 |
| B3 | 1.247 | Positive | 124 | 79 | −10 | −24 | −15 |
| B9 | 2.971 | Positive | 442 | 176 | −16 | −39 | −19 |
| L | 1.060 | Negative | 341 | 161.15 | 13 | 8 | 15 |
| LA | 1.594 | Negative | 89.25 | 43.05 | 19 | 13 | 14 |
| Linearity (µg/mL) | R2 | Standard Curve Equation | LOD (µg/mL) | LOQ (µg/mL) | |
|---|---|---|---|---|---|
| B2 | 0.005–10 | 0.9941 | y = 1,137,260x − 89,427 | 0.001 | 0.0033 |
| B3 | 0.25–10 | 0.9985 | y = 8819.9x − 1000.03 | 0.050 | 0.1650 |
| B9 | 0.025–10 | 0.9980 | y = 104,830x + 11,539 | 0.005 | 0.0165 |
| L | 0.01–400 | 0.9963 | y = 19,947x + 122,904 | 0.005 | 0.0165 |
| LA | 0.01–2000 | 0.9972 | y = 18,770x + 651,622 | 0.005 | 0.0165 |
| Identification by MALDI | 16S rRNA Identification | Number in PCM ** | |||
|---|---|---|---|---|---|
| Identification Result | MSP Log Score * | Identification Result | % Similarity | Strand Length (bp) | |
| Lactococcus lactis ssp lactis 3C1_QSA IBS | 2.02 | Lactococcus lactis strain NBRC 100933 [NR_113960.1] Lactococcus lactis strain NCDO 604 [NR_040955.1] | 99.86 99.86 | 1412 | L. lactis Beet 1 [OM957496] |
| Weissella cibaria DSM 15878T DSM | 2.13 | Weissella cibaria strain II-I-59 [NR_036924.1] Weissella confusa strain JCM 1093 [NR_040816.1] | 99.58 99.17 | 1442 | W. cibaria Beet 2 [OM957500] |
 | |||||
|---|---|---|---|---|---|
| Compound * | Adduct Formula | m/z Exp. | m/z Calc. | Δ (ppm) | |
| 1. | [FA oxo(15:0)] 4-oxo-pentadecanoic acid | [C15H28O3 + 107Ag]+ | 363.108 | 363.108 | 0.0 |
| 2. | [FA hydroxy(17:2)]12-hydroxy-8E,10Eheptadecadienoic acid | [C17H30O3 + 107Ag]+ | 389.123 | 389.124 | −2.6 |
| 3. | Dehydroabietadienal | [C20H28O + 107Ag]+ | 391.120 | 391.119 | 2.6 |
| [C20H28O + 109Ag]+ | 393.119 | 393.118 | 2.5 | ||
| 4. | [GP (6:0/6:0)] 1,2-dihexanoyl-sn-glycero-3-phosphate | [C15H29O8P + K]+ | 407.123 | 407.123 | 0.0 |
| 5. | Ala-Leu-Ala-Gly | [C14H26N4O5 + 107Ag]+ | 437.096 | 437.095 | 2.3 |
| 6. | Met-Phe-Cys | [C17H25N3O4S2 + K]+ | 438.093 | 438.092 | 2.3 |
| 7. | Cys-Trp-Gly-Gly | [C18H23N5O5S + Na]+ | 444.130 | 444.131 | −2.3 |
| 8. | Asn-Thr-Cys-Ser | [C14H25N5O8S + Na]+ | 446.132 | 446.132 | 0.0 |
| 9. | Ala-Ala-Ala-Asp | [C13H22N4O7 + 107Ag]+ | 453.052 | 453.053 | −2.2 |
| 10. | Ala-Asn-Gly-Ser | [C12H21N5O7 + 107Ag]+ | 454.050 | 454.049 | 2.2 |
| 11. | [GP (6:0/6:0)] 1,2-dihexanoyl-sn-glycero-3-phosphate | [C15H29O8P + 107Ag]+ | 475.066 | 475.065 | 2.1 |
| 12. | Val-Asp-His | [C15H23N5O6 + 107Ag]+ | 476.069 | 476.069 | 0.0 |
| 13. | [GP (6:0/6:0)] 1,2-dihexanoyl-sn-glycero-3-phosphate | [C15H29O8P + 109Ag]+ | 477.065 | 477.064 | 2.1 |
| 14. | PA(17:2(9Z,12Z)/0:0) | [C20H37O7P + 109Ag]+ | 529.130 | 529.132 | −3.8 |
| 15. | Ala-Trp-Val-Tyr | [C28H35N5O6 + H]+ | 538.267 | 538.266 | 1.9 |
| 16. | C12-ACP (Dodecanoyl-ACP) | [C23H44N2O8PS + H]+ | 540.263 | 540.263 | 0.0 |
| 17. | LysoPE(20:3(8Z,11Z,14Z)/0:0) | [C25H46NO7P + K]+ | 542.259 | 542.264 | −9.2 |
| 18. | Asp-Leu-Met-His | [C21H34N6O7S + K]+ | 553.184 | 553.184 | 0.0 |
| 19. | Alpha-Trisaccharide | [C20H37NO14 + K]+ | 554.187 | 554.185 | 3.6 |
| 20. | Ala-Ala-Tyr-His | [C21H28N6O6 + 109Ag]+ | 569.110 | 569.111 | −1.8 |
| 21. | Glu-Phe-Ala-Pro | [C22H30N4O7 + 109Ag]+ | 571.116 | 571.116 | 0.0 |
| 22. | Cys-Pro-Pro-Tyr | [C22H30N4O6S + 107Ag]+ | 585.094 | 585.093 | 1.7 |
| 23. | Ala-Cys-Tyr-His | [C21H28N6O6S + 107Ag]+ | 599.084 | 599.084 | 0.0 |
| [C21H28N6O6S + 109Ag]+ | 601.085 | 601.083 | 3.3 | ||
| 24. | Cellotriose | [C18H32O16 + 109Ag]+ | 613.075 | 613.073 | 3.3 |
| 25. | Octacosanyl hexadecanoate | [C44H88O2 + H]+ | 649.684 | 649.686 | −3.1 |
| 26. | FAD stem group | [C15H26N6O13P2 + 109Ag]+ | 669.009 | 669.008 | 1.5 |
| 27. | GalCer(d18:2/20:0) | [C44H83NO8 + H]+ | 754.621 | 754.619 | 2.7 |
| 28. | TG(13:0/13:0/17:2(9Z,12Z)) | [C46H84O6 + Na]+ | 755.614 | 755.616 | −2.6 |
| 29. | PC(18:2(9Z,12Z)/P-18:1(11Z)) | [C44H82NO7P + H]+ | 768.592 | 768.590 | 2.6 |
| 30. | [PC (16:2/18:1)] 1-hexadecyl-2-(9Z-octadecenyl)sn- glycero-3-phosphocholine | [C42H86NO6P + K]+ | 770.587 | 770.582 | 6.5 |
| 31. | PA(19:0/22:2(13Z,16Z)) | [C44H83O8P + H]+ | 771.584 | 771.590 | −7.8 |
| 32. | PC(15:0/20:2(11Z,14Z)) | [C43H82NO8P + H]+ | 772.582 | 772.585 | −3.9 |
| 33. | PG(P-20:0/17:2(9Z,12Z)) | [C43H81O9P + H]+ | 773.572 | 773.569 | 3.9 |
| 34. | PC(14:1(9Z)/18:3(6Z,9Z,12Z)) | [C40H72NO8P + 109Ag]+ | 834.407 | 834.404 | 3.6 |
| 35. | [PE (6:0/8:0)] 1-(6-[5]-ladderane-hexanyl)-2(8-[3]- ladderane-octanyl)-sn-glycerophosphoethanolamine | [C43H72NO6P + 107Ag]+ | 836.407 | 836.414 | −8.4 |
| 36. | [PC (18:1/22:6)] 1-(11Z-octadecenoyl)-2-(4Z,7Z,10Z,13Z,16Z,19Z-docosahexaenoyl)-sn-glycero-3-phosphocholine | [C48H82NO8P + K]+ | 870.543 | 870.541 | 2.3 |
| 37. | PI(16:1(9Z)/19:1(9Z)) | [C44H81O13P + Na]+ | 871.535 | 871.531 | 4.6 |
| 38. | PS(O-20:0/16:0) | [C42H84NO9P + 109Ag]+ | 886.494 | 886.493 | 1.1 |
| 39. | PI(15:0/20:2(11Z,14Z)) | [C44H81O13P + K]+ | 887.497 | 887.505 | −9.0 |
| 40. | PE(P-18:0/22:4(7Z,10Z,13Z,16Z)) | [C45H82NO7P + 109Ag]+ | 888.495 | 888.487 | 9.0 |
| 41. | PS(O-16:0/21:0) | [C43H86NO9P + 107Ag]+ | 898.505 | 898.509 | −4.5 |
| 42. | PI(16:0/20:3(8Z,11Z,14Z)) | [C45H81O13P + K]+ | 899.504 | 899.505 | −1.1 |
| 43. | (3′-sulfo)Galbeta-Cer(d18:1/2-OH-16:0) | [C40H77NO12S + 107Ag]+ | 902.422 | 902.421 | 1.1 |
| 44. | PG(16:1(9Z)/22:4(7Z,10Z,13Z,16Z)) | [C44H77O10P + 107Ag]+ | 903.428 | 903.430 | −2.2 |
| 45. | PE(22:1(13Z)/24:1(15Z)) | [C51H98NO8P + Na]+ | 906.688 | 906.692 | −4.4 |
| 46. | PS(15:0/22:1(11Z)) | [C43H82NO10P + 107Ag]+ | 910.468 | 910.472 | −4.4 |
| 47. | PC(18:3(6Z,9Z,12Z)/20:4(5Z,8Z,11Z,14Z)) | [C46H78NO8P + 109Ag]+ | 912.451 | 912.451 | 0.0 |
| 48. | PG(P-18:0/22:6(4Z,7Z,10Z,13Z,16Z,19Z)) | [C46H79O9P + 107Ag]+ | 913.450 | 913.451 | −1.1 |
| 49. | (3′-sulfo)GalBeta-Cer(d18:1/18:0) | [C42H81NO11S + 107Ag]+ | 914.455 | 914.458 | −3.3 |
| 50. | PI(P-16:0/17:0) | [C42H81O12P + 107Ag]+ | 915.455 | 915.451 | 4.4 |
| 51. | (3′-sulfo)GalBeta-Cer(d18:1/18:0) | [C42H81NO11S + 109Ag]+ | 916.463 | 916.457 | 6.5 |
| 52. | PI(O-16:0/18:3(9Z,12Z,15Z)) | [C43H79O12P + 109Ag]+ | 927.437 | 927.435 | 2.2 |
| 53. | PS(17:2(9Z,12Z)/22:4(7Z,10Z,13Z,16Z)) | [C45H76NO10P + 107Ag]+ | 928.421 | 928.425 | −4.3 |
| 54. | PS(18:1(9Z)/22:6(4Z,7Z,10Z,13Z,16Z,19Z)) | [C46H76NO10P + 107Ag]+ | 940.430 | 940.425 | 5.3 |
| 55. | PI(16:1(9Z)/18:1(9Z)) | [C43H79O13P + 107Ag]+ | 941.432 | 941.430 | 2.1 |
| 56. | [PE (22:6/22:6)] 1,2-di-(4Z,7Z,10Z,13Z,16Z,19Z-docosahexaenoyl)-sn-glycero-3-phosphoethanolamine | [C49H74NO8P + 107Ag]+ | 942.413 | 942.420 | −7.4 |
| 57. | GalCer(d18:0/26:1) | [C50H97NO8 + 109Ag]+ | 948.626 | 948.626 | 0.0 |
| 58. | [GL (18:0/20:0/20:0)] 1-octadecanoyl-2,3-dieicosanoyl-sn-glycerol | [C61H118O6 + K]+ | 985.857 | 985.856 | 1.01 |
| 59. | [PC (24:0/26:0)] 1-tetracosanoyl-2-hexacosanoyl-sn- glycero-3-phosphocholine | [C58H116NO8P + H]+ | 986.859 | 986.851 | 8.1 |
| 60. | PE(22:2(13Z,16Z)/24:1(15Z)) | [C51H96NO8P + 109Ag]+ | 990.593 | 990.592 | 1.0 |
| 61. | [GP (18:0/18:0/2:0/2:0)] 1,2-di-(9Z-octadecenoyl)-sn- glycero-3-cytidine-5′-diphosphate | [C48H85N3O15P2 + H]+ | 1006.555 | 1006.552 | 3.0 |
| 62. | Siroheme amide | [C42H47FeN5O15 + 107Ag]+ | 1024.151 | 1024.146 | 4.9 |
| 63. | 2-Oxo-delta3-4,5,5-trimethylcyclopentenylacetyl-CoA | [C31H48N7O18P3S + 109Ag]+ | 1040.106 | 1040.103 | 2.9 |
| 64. | CoA(22:6(4Z,7Z,10Z,13Z,16Z,19Z)) | [C43H66N7O17P3S + 107Ag]+ | 1184.255 | 1184.249 | 5.1 |
| [C43H66N7O17P3S + 109Ag]+ | 1186.246 | 1186.249 | −2.5 | ||
 | |||||
|---|---|---|---|---|---|
| Compound * | Adduct Formula | m/z Exp. | m/z Calc. | Δ (ppm) | |
| 1. | [FA hydroxy(15:0)] 2-hydroxy-pentadecanoic acid | [C15H30O3 + 107Ag]+ | 365.12 | 365.12 | 0.0 |
| 2. | N-palmitoyl proline | [C21H39NO3 + Na]+ | 376.28 | 376.28 | 0.0 |
| 3. | Galactan | [C14H26O11 + Na]+ | 393.14 | 393.14 | 0.0 |
| 4. | [PC (6:0)] 1-hexanoyl-sn-glycero3-phosphocholine | [C14H30NO7P + K]+ | 394.14 | 394.14 | 0.0 |
| 5. | 2-hydroxy-9Z,12Z,15Z-octadecatrienoic acid | [C18H30O3 + 109Ag]+ | 403.12 | 403.12 | 0.0 |
| 6. | Met-Tyr | [C14H20N2O4S + 107Ag]+ | 419.02 | 419.02 | 0.0 |
| 7. | [PC (3:0)] 1-(2E-propionyl)sn-glycero-3- phosphocholine | [C11H22NO7P + 109Ag]+ | 420.02 | 420.02 | 0.0 |
| 8. | S-(2-Chloroacetyl)glutathione | [C12H18N3O7ClS + K]+ | 422.02 | 422.02 | 0.0 |
| 9. | Val-Gly-Arg | [C13H26N6O4 + 107Ag]+ | 437.11 | 437.11 | 0.0 |
| 10. | S-Adenosylmethionine | [C15H23N6O5S + K]+ | 438.11 | 438.11 | 0.0 |
| 11. | Ala-Cys-Gly-Arg | [C14H27N7O5S + K]+ | 444.14 | 444.14 | 0.0 |
| 12. | Phe-Trp-Gly | [C22H24N4O4 + K]+ | 447.14 | 447.14 | 0.0 |
| 13. | Asp-Gly-Arg | [C12H22N6O6 + 107Ag]+ | 453.07 | 453.06 | 22.1 |
| 14. | Ala-Ala-Ala-Asn | [C13H23N5O6 + 109Ag]+ | 454.07 | 454.07 | 0.0 |
| 15. | Dopaxanthin | [C18H18N2O8 + 107Ag]+ | 497.01 | 497.01 | 0.0 |
| [C18H18N2O8 + 109Ag]+ | 499.01 | 499.01 | 0.0 | ||
| 16. | CPA(18:2(9Z,12Z)/0:0) | [C21H37O6P + 109Ag]+ | 525.14 | 525.14 | 0.0 |
| 17. | CPA(18:1(11Z)/0:0) | [C21H39O6P + 109Ag]+ | 527.15 | 527.15 | 0.0 |
| 18. | Ile-Lys-Tyr | [C21H34N4O5 + 107Ag]+ | 529.16 | 529.16 | 0.0 |
| 19 | PS(17:2(9Z,12Z)/0:0) | [C23H42NO9P + Na]+ | 530.25 | 530.25 | 0.0 |
| 20. | PA(17:0/0:0) | [C20H41O7P + 107Ag]+ | 531.16 | 531.16 | 0.0 |
| 21. | Arg-Ile-Ile-Pro | [C23H43N7O5 + K]+ | 536.29 | 536.30 | −18.6 |
| 22. | PG(12:0/0:0) | [C18H37O9P + 109Ag]+ | 537.12 | 537.12 | 0.0 |
| 23. | LysoPC(18:4(6Z,9Z,12Z,15Z)) | [C26H46NO7P + Na]+ | 538.29 | 538.29 | 0.0 |
| 24. | Ala-Glu-Ile-Thr | [C18H32N4O8 + 107Ag]+ | 539.13 | 539.13 | 0.0 |
| 25. | Asn-Leu-Phe-Phe | [C28H37N5O6 + H]+ | 540.28 | 540.28 | 0.0 |
| 26. | Ala-Phe-Val-Pro | [C22H32N4O5 + 109Ag]+ | 541.14 | 541.14 | 0.0 |
| 27. | His-Lys-Val-His | [C23H37N9O5 + Na]+ | 542.28 | 542.28 | 0.0 |
| 28. | Glu-Glu-Met-Pro | [C20H32N4O9S + K]+ | 543.15 | 543.15 | 0.0 |
| 29. | PC(O-10:1(9E)/2:0) | [C20H40NO7P + 107Ag]+ | 544.16 | 544.16 | 0.0 |
| 30. | LysoPA(0:0/18:1(9Z)) | [C21H41O7P + 109Ag]+ | 545.16 | 545.16 | 0.0 |
| 31. | Asn-Trp-Asp-Pro | [C24H30N6O8 + Na]+ | 553.20 | 553.20 | 0.0 |
| 32. | Lys-Trp-Gly-Gly | [C21H30N6O5 + 109Ag]+ | 555.13 | 555.13 | 0.0 |
| 33. | Ala-Cys-Tyr-Tyr | [C24H30N4O7S + K]+ | 557.14 | 557.15 | −17.9 |
| 34. | Arg-Asn-Asp-Gly | [C16H28N8O8 + 107Ag]+ | 567.11 | 567.11 | 0.0 |
| 35. | Phe-Thr-Pro-Pro | [C23H32N4O6 + 109Ag]+ | 569.14 | 569.14 | 0.0 |
| 36. | Thr-Trp-Arg | [C21H31N7O5 + 109Ag]+ | 570.14 | 570.14 | 0.0 |
| 37. | Ile-Met-Thr-Thr | [C19H36N4O7S + 107Ag]+ | 571.13 | 571.14 | −17.5 |
| 38. | Cys-Ser-Tyr-Tyr | [C24H30N4O8S + K]+ | 573.14 | 573.14 | 0.0 |
| 39. | Phe-Gly-Ser-Tyr | [C23H28N4O7 + 109Ag]+ | 581.10 | 581.10 | 0.0 |
| 40. | Asn-Leu-Asn-Asp | [C18H30N6O9 + 109Ag]+ | 583.11 | 583.11 | 0.0 |
| 41. | Ala-Phe-Asn-Gln | [C21H30N6O7 + 107Ag]+ | 585.12 | 585.12 | 0.0 |
| [C21H30N6O7 + 109Ag]+ | 587.12 | 587.12 | 0.0 | ||
| 42. | His-Met-Phe-Gly | [C22H30N6O5S + 107Ag]+ | 597.10 | 597.10 | 0.0 |
| 43. | Arg-Cys-Gln-Ser | [C17H32N8O7S + 107Ag]+ | 599.12 | 599.12 | 0.0 |
| [C17H32N8O7S + 109Ag]+ | 601.12 | 601.12 | 0.0 | ||
| 44. | Cys-Met-Phe-Thr | [C21H32N4O6S2 + 109Ag]+ | 609.08 | 609.08 | 0.0 |
| 45. | Asp-Met-Thr-His | [C19H30N6O8S + 109Ag]+ | 611.09 | 611.09 | 0.0 |
| 46. | Ala-Trp-Cys-Gln | [C22H30N6O6S + 107Ag]+ | 613.10 | 613.10 | 0.0 |
| [C22H30N6O6S + 109Ag]+ | 615.10 | 615.10 | 0.0 | ||
| 47. | Asp-Met-Asp-His | [C19H28N6O9S + 107Ag]+ | 623.07 | 623.07 | 0.0 |
| 48. | Cys-Met-Thr-Tyr | [C21H32N4O7S2 + 109Ag]+ | 625.08 | 625.08 | 0.0 |
| 49. | Poly-g-D-glutamate | [C20H30N4O12 + 109Ag]+ | 627.09 | 627.09 | 0.0 |
| 50. | Asn-Met-Met-Gln | [C19H34N6O7S2 + 107Ag]+ | 629.10 | 629.10 | 0.0 |
| 51. | P-(bromoacetamido)phenyl uridylpyrophosphate | [C17H20BrN3O13P2 + Na]+ | 637.96 | 637.95 | 15.7 |
| 52. | Asn-Trp-Asp-Cys | [C22H28N6O8S + 107Ag]+ | 643.08 | 643.07 | 15.6 |
| 53. | TG(12:0/12:0/20:0) | [C47H90O6 + H]+ | 751.68 | 751.68 | 0.0 |
| 54. | Stigmast-5,22E-dien-3beta-yl (13Z-docosenoate) | [C51H88O2 + Na]+ | 755.67 | 755.67 | 0.0 |
| [C51H88O2 + K]+ | 771.64 | 771.64 | 0.0 | ||
| 55. | PE(15:0/22:0) | [C42H84NO8P + H]+ | 762.60 | 762.60 | 0.0 |
| 56. | GlcCer(d18:2/21:0) | [C45H85NO8 + H]+ | 768.64 | 768.63 | 13.0 |
| 57. | PS(17:1(9Z)/20:4(5Z,8Z,11Z,14Z)) | [C43H74NO10P + K]+ | 834.47 | 834.47 | 0.0 |
| 58. | PG(18:1(9Z)/20:4(5Z,8Z,11Z,14Z)) | [C44H77O10P + K]+ | 835.48 | 835.49 | −12.0 |
| 59. | N-(2-hydroxy-eicosanoyl)-1-beta-glucosyl-4E,6E-pentadecasphingadienine | [C41H77NO9 + 109Ag]+ | 836.47 | 836.46 | 12.0 |
| 60. | PG(20:5(5Z,8Z,11Z,14Z,17Z)/20:5(5Z,8Z,11Z,14Z,17Z)) | [C46H71O10P + Na]+ | 837.47 | 837.47 | 0.0 |
| 61. | TG(16:1(9Z)/18:3(6Z,9Z,12Z)/18:3(6Z,9Z,12Z)) | [C55H92O6 + H]+ | 849.70 | 849.70 | 0.0 |
| 62. | [PC (3:0/3:0/3:0)] 1-(2E,6E,10E-phytatrienyl)-2-(2E,6E10E-phytatrienyl)-sn-glycero-3-phosphocholine | [C48H88NO6P + 107Ag]+ | 912.54 | 912.54 | 0.0 |
| 63. | PG(O-20:0/19:1(9Z)) | [C45H89O9P + 109Ag]+ | 913.53 | 913.53 | 0.0 |
| 64. | PG(22:1(11Z)/22:6(4Z,7Z,10Z,13Z,16Z,19Z)) | [C50H85O10P + K]+ | 915.56 | 915.55 | 10.9 |
| 65. | PC(22:4(7Z,10Z,13Z,16Z)/P-18:1(11Z)) | [C48H86NO7P + 107Ag]+ | 926.51 | 926.52 | −10.8 |
| 66. | PS(17:0/22:1(11Z)) | [C45H86NO10P + 109Ag]+ | 940.51 | 940.50 | 10.6 |
| 67. | PIP(16:0/18:0) | [C43H84O16P2 + Na]+ | 941.51 | 941.51 | 0.0 |
| 68. | LacCer(d18:0/14:0) | [C44H85NO13 + 107Ag]+ | 942.51 | 942.51 | 0.0 |
| 69. | PI(17:2(9Z,12Z)/18:4(6Z,9Z,12Z,15Z)) | [C44H73O13P + 107Ag]+ | 947.39 | 947.38 | 10.6 |
| [C44H73O13P + 109Ag]+ | 949.39 | 949.38 | 10.5 | ||
| 70. | PS(P-20:0/22:6(4Z,7Z,10Z,13Z,16Z,19Z)) | [C48H82NO9P + 107Ag]+ | 954.49 | 954.48 | 10.5 |
| [C48H82NO9P + 109Ag]+ | 956.48 | 956.48 | 0.0 | ||
| 71. | PI(O-16:0/20:2(11Z,14Z)) | [C45H85O12P + 107Ag]+ | 955.49 | 955.48 | 10.5 |
| 72. | PI(15:0/20:1(11Z)) | [C44H83O13P + 107Ag]+ | 957.47 | 957.46 | 10.4 |
| 73. | PI(17:1(9Z)/20:3(8Z,11Z,14Z)) | [C46H81O13P + 109Ag]+ | 981.44 | 981.45 | −10.2 |
| 74. | TG(19:1(9Z)/21:0/21:0) | [C64H122O6 + H]+ | 987.94 | 987.93 | 10.1 |
| 75. | 5-formyl-tetrahydrosarcinapterin | [C36H52N7O20P + 107Ag]+ | 1040.21 | 1040.21 | 0.0 |
| 76. | Lipid IVA | [C68H126N2O23P2 + H]+ | 1401.84 | 1401.83 | 7.1 |
| 77. | Tetrahexosylceramide (d18:1/26:1(17Z)) | [C70H128N2O23 + K]+ | 1403.85 | 1403.85 | 0.0 |
| Concentration (µg/mL) | |||
|---|---|---|---|
| Sample | Niacin—B3 | Folic Acid—B9 | Riboflavin—B2 |
| TEMPERATURE 20 °C | |||
| Niacin Assay Medium | 0.178 ± 0.009 | nd | 4.104 ± 0.126 |
| L. lactis | 3.582 ± 0.132 | nd | 1.409 ± 0.045 |
| W. cibaria | nd | LOD | −2.276 ± 0.118 |
| MRS Broth Medium | nd | nd | nd |
| L. lactis | 0.718 ± 0.000 | nd | 0.147 ± 0.006 |
| W. cibaria | 1.465 ± 0.188 | nd | 0.109 ± 0.002 |
| TEMPERATURE 30 °C | |||
| Niacin Assay Medium | 0.174 ± 0.001 | nd | 4.592 ± 0.095 |
| L. lactis | 1.273 ± 0.177 | LOD | 0.754 ± 0.065 |
| W. cibaria | 0.081 ± 0.005 LOQ | LOD | −0.307 ± 0.495 |
| MRS Broth Medium | 1.350 ± 0.073 | nd | 4.011 ± 0.067 |
| L. lactis | −0.874 ± 0.072 | nd | −3.871 ± 0.001 |
| W. cibaria | −0.872 ± 0.058 | nd | −1.314 ± 0.115 |
| Sample | Lactose (µg/mL) | Lactic Acid (µg/mL) |
|---|---|---|
| TEMPERATURE 20 °C | ||
| LAPTg Medium | 38.61 ± 0.58 | 34.01 ± 1.88 |
| L. lactis | 2.61 ± 0.81 | 1014.99 ± 47.59 |
| W. cibaria | 14.49 ± 1.14 | 310.99 ± 7.48 |
| TEMPERATURE 30 °C | ||
| LAPTg Medium | 49.91 ± 1.00 | 16.43 ± 0.22 |
| L. lactis | 12.38 ± 0.95 | 638.21 ± 8.03 |
| W. cibaria | 0.91 ± 0.50 | 277.36 ± 5.82 |
| Pathogenic Strain | Number of Grown Pathogen Colonies (Percentage of Inhibition) | ||
|---|---|---|---|
| Control | L. lactis | W. cibaria | |
| Escherichia coli | 340 ± 63 | 174 | 156 |
| 49% | 54% | ||
| Enterobacter cloacae | 62 ± 13 | 22 | 5 |
| 65% | 92% | ||
| Pseudomonas aeruginosa | 204 ± 12 | 0 | 0 |
| 100% | 100% | ||
| Listeria innocua | 956 ± 35 | 0 | 0 |
| 100% | 100% | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maślak, E.; Złoch, M.; Arendowski, A.; Sugajski, M.; Janczura, I.; Rudnicka, J.; Walczak-Skierska, J.; Buszewska-Forajta, M.; Rafińska, K.; Pomastowski, P.; et al. Isolation and Identification of Lactococcus lactis and Weissella cibaria Strains from Fermented Beetroot and an Investigation of Their Properties as Potential Starter Cultures and Probiotics. Foods 2022, 11, 2257. https://doi.org/10.3390/foods11152257
Maślak E, Złoch M, Arendowski A, Sugajski M, Janczura I, Rudnicka J, Walczak-Skierska J, Buszewska-Forajta M, Rafińska K, Pomastowski P, et al. Isolation and Identification of Lactococcus lactis and Weissella cibaria Strains from Fermented Beetroot and an Investigation of Their Properties as Potential Starter Cultures and Probiotics. Foods. 2022; 11(15):2257. https://doi.org/10.3390/foods11152257
Chicago/Turabian StyleMaślak, Ewelina, Michał Złoch, Adrian Arendowski, Mateusz Sugajski, Izabela Janczura, Joanna Rudnicka, Justyna Walczak-Skierska, Magdalena Buszewska-Forajta, Katarzyna Rafińska, Paweł Pomastowski, and et al. 2022. "Isolation and Identification of Lactococcus lactis and Weissella cibaria Strains from Fermented Beetroot and an Investigation of Their Properties as Potential Starter Cultures and Probiotics" Foods 11, no. 15: 2257. https://doi.org/10.3390/foods11152257