
Determination of 14 Isoflavone Isomers in Natto by UPLC-ESI-MS/MS and Antioxidation and Antiglycation Profiles

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. The Fermentation of Soybean and Isoflavone Preparation
2.3. Characterization of Soybean Isoflavone from Natto
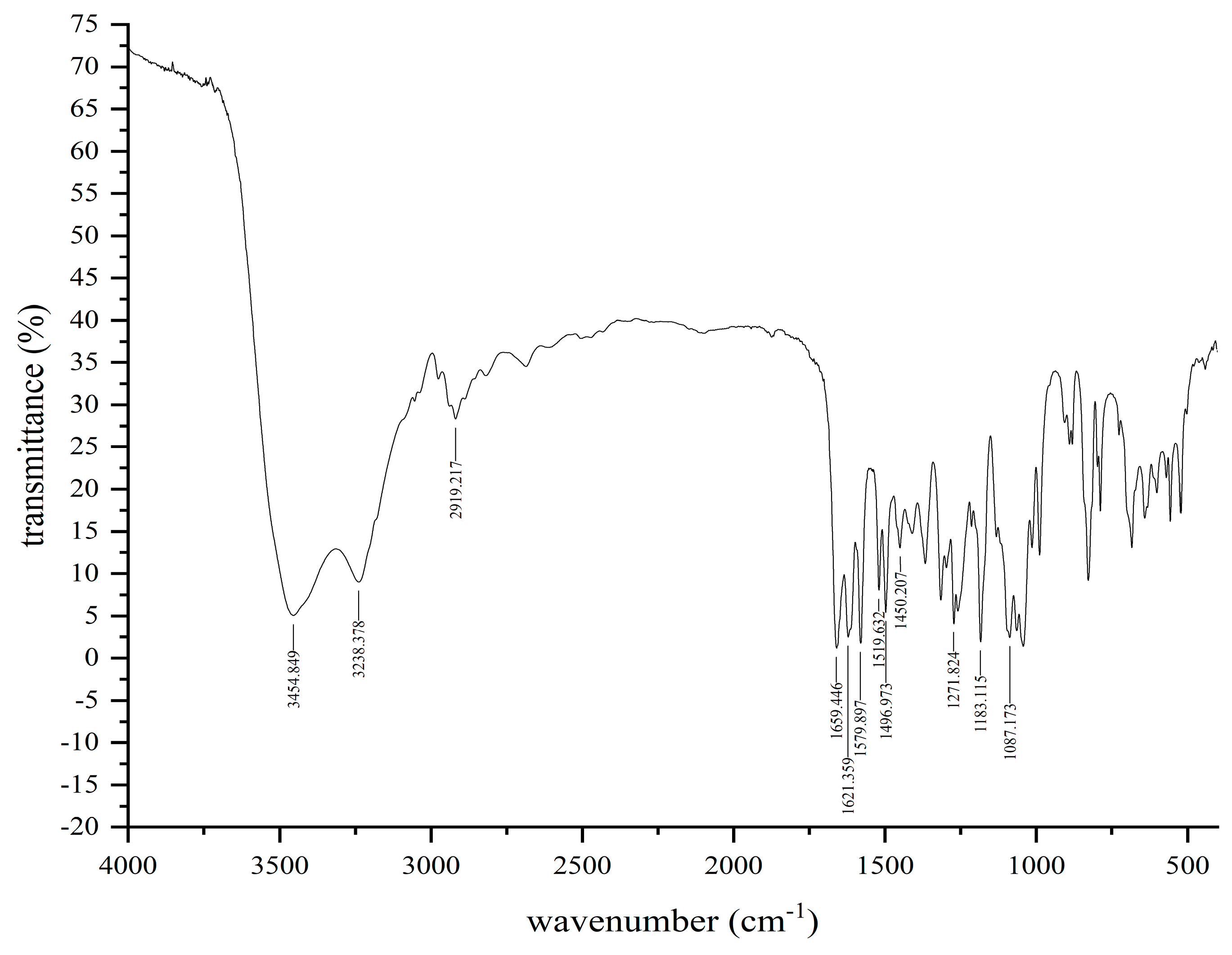
2.3.1. FT-IR (Fourier Transform Infrared Spectoscopy) Analysis
2.3.2. HPLC and UV Analysis
2.4. UPLC-ESI-MS/MS Analysis of Soybean Isoflavone from Natto
2.5. Antioxidant Capacity of Isoflavones from Natto
2.5.1. DPPH Radical Scavenging Capacity
2.5.2. ABTS Radical Scavenging Capacity
2.5.3. O2− Radical Scavenging Capacity
2.5.4. Ferric Reducing Antioxidant Power (FRAP) Capacity
2.6. In Vitro Inhibition of Advanced Glycation End Products by Soybean Isoflavones from Natto
2.6.1. Fructose-BSA System
2.6.2. α-lactose-Lys System
2.6.3. BSA-MGO System
2.7. Mechanism of Genistein Inhibiting AGEs in BSA-MGO System
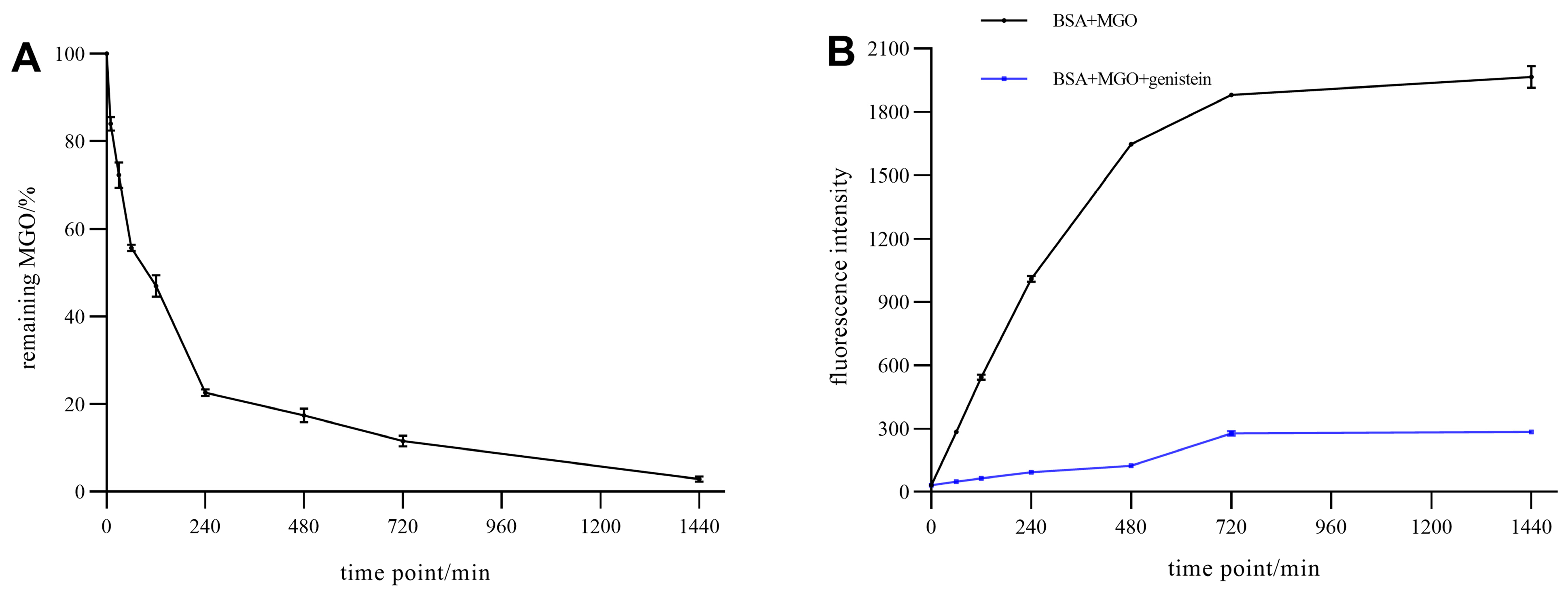
2.7.1. Kinetic Study of the Trapping of MGO by Genistein
2.7.2. Kinetic Study of the Inhibitory Effects on the Formation of AGEs by Genistein
2.7.3. UPLC-Q-TO-ESI-MS/MS Analysis of Genistein-MGO Adducts
2.8. Statistical Analysis
3. Results
3.1. Nutrient Components Analysis of Natto and Isoflavones Preparation
3.2. Spectral Characterization of Soybean Isoflavone in Natto
3.2.1. HPLC and UV Analysis
3.2.2. FT-IR Analysis
3.2.3. UPLC-ESI-MS/MS Analysis
3.3. Antioxidant Capacity of Soybean Isoflavones from Natto
3.4. Anti-Glycation Activity of Soybean Isoflavones from Natto
3.5. Mechanism of Genistein Inhibiting AGEs in BSA-MGO System
3.5.1. Kinetic Study of the Inhibitory Effects on the Formation of AGEs by Genistein
3.5.2. UPLC-Q-TOF-ESI-MS/MS Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cao, Z.-H.; Green-Johnson, J.M.; Buckley, N.D.; Lin, Q.-Y. Bioactivity of soy-based fermented foods: A review. Biotechnol. Adv. 2019, 37, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented Foods: Definitions and Characteristics, Impact on the Gut Microbiota and Effects on Gastrointestinal Health and Disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef] [Green Version]
- Zaheer, K.; Humayoun Akhtar, M. An updated review of dietary isoflavones: Nutrition, processing, bioavailability and impacts on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1280–1293. [Google Scholar] [CrossRef]
- Brouns, F. Soya isoflavones: A new and promising ingredient for the health foods sector. Food Res. Int. 2002, 35, 187–193. [Google Scholar] [CrossRef]
- Lee, M.; Hong, G.-E.; Zhang, H.; Yang, C.-Y.; Han, K.-H.; Mandal, P.K.; Lee, C.-H. Production of the isoflavone aglycone and antioxidant activities in black soymilk using fermentation with Streptococcus thermophilus S10. Food Sci. Biotechnol. 2015, 24, 537–544. [Google Scholar] [CrossRef]
- Miadokova, E. Isoflavonoids—An overview of their biological activities and potential health benefits. Interdiscip. Toxicol. 2009, 2, 211–218. [Google Scholar] [CrossRef]
- Popa, D.-S.; Rusu, M.E. Isoflavones: Vegetable Sources, Biological Activity, and Analytical Methods for Their Assessment. In Superfood and Functional Food—The Development of Superfoods and Their Roles as Medicine; IntechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Jung, Y.S.; Rha, C.S.; Baik, M.Y.; Baek, N.I.; Kim, D.O. A brief history and spectroscopic analysis of soy isoflavones. Food Sci. Biotechnol. 2020, 29, 1605–1617. [Google Scholar] [CrossRef]
- Wang, H.J.; Chang, L.; Lin, Y.S. Changes in Functionality of Germinated and Non-Germinated Brown Rice Fermented by Bacillus natto. Foods 2021, 10, 2779. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Jung, Y.S.; Jang, D.; Cho, C.H.; Lee, S.H.; Han, N.S.; Kim, D.O. Antioxidant capacity of 12 major soybean isoflavones and their bioavailability under simulated digestion and in human intestinal Caco-2 cells. Food Chem. 2022, 374, 131493. [Google Scholar] [CrossRef]
- Kojima, A.; Ikehara, S.; Kamiya, K.; Kajita, E.; Sato, Y.; Kouda, K.; Tamaki, J.; Kagamimori, S.; Iki, M. Natto Intake is Inversely Associated with Osteoporotic Fracture Risk in Postmenopausal Japanese Women. J. Nutr. 2020, 150, 599–605. [Google Scholar] [CrossRef]
- Soukup, S.T.; Stoll, D.A.; Danylec, N.; Schoepf, A.; Kulling, S.E.; Huch, M. Metabolism of Daidzein and Genistein by Gut Bacteria of the Class Coriobacteriia. Foods 2021, 10, 2741. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Uesugi, T.; Hirai, K.; Nukaya, H.; Tsuji, K.; Ishida, H. New 6-O-acyl isoflavone glycosides from soybeans fermented with Bacillus subtilis (natto). I. 6-O-succinylated isoflavone glycosides and their preventive effects on bone loss in ovariectomized rats fed a calcium-deficient diet. Biol. Pharm. Bull. 1999, 22, 1193–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soman, S.; Rauf, A.A.; Indira, M.; Rajamanickam, C. Antioxidant and antiglycative potential of ethyl acetate fraction of Psidium guajava leaf extract in streptozotocin-induced diabetic rats. Plant Foods Hum. Nutr. 2010, 65, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Uribarri, J.; Woodruff, S.; Goodman, S.; Cai, W.; Chen, X.; Pyzik, R.; Yong, A.; Striker, G.E.; Vlassara, H. Advanced glycation end products in foods and a practical guide to their reduction in the diet. J. Am. Diet. Assoc. 2010, 110, 911–916 e912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulsen, M.W.; Hedegaard, R.V.; Andersen, J.M.; de Courten, B.; Bugel, S.; Nielsen, J.; Skibsted, L.H.; Dragsted, L.O. Advanced glycation endproducts in food and their effects on health. Food Chem. Toxicol. 2013, 60, 10–37. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Shao, X.; Chen, H.; Ho, C.T.; Sang, S. Genistein inhibits advanced glycation end product formation by trapping methylglyoxal. Chem. Res. Toxicol. 2011, 24, 579–586. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Bartosz, G. Prevention of protein glycation by natural compounds. Molecules 2015, 20, 3309–3334. [Google Scholar] [CrossRef]
- Wang, C.; Lu, Y.; Huang, Q.; Zheng, T.; Sang, S.; Lv, L. Levels and formation of alpha-dicarbonyl compounds in beverages and the preventive effects of flavonoids. J. Food Sci. Technol. 2017, 54, 2030–2040. [Google Scholar] [CrossRef]
- Shen, Y.; Xu, Z.; Sheng, Z. Ability of resveratrol to inhibit advanced glycation end product formation and carbohydrate-hydrolyzing enzyme activity, and to conjugate methylglyoxal. Food Chem. 2017, 216, 153–160. [Google Scholar] [CrossRef]
- Li, C.; Xu, T.; Liu, X.W.; Wang, X.; Xia, T. The expression of β-glucosidase during natto fermentation increased the active isoflavone content. Food Biosci. 2021, 43, 101286. [Google Scholar] [CrossRef]
- Yang, Y.; Lan, G.; Tian, X.; He, L.; Li, C.; Zeng, X.; Wang, X. Effect of Fermentation Parameters on Natto and Its Thrombolytic Property. Foods 2021, 10, 2547. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Mu, T.; Sun, H. Preparative purification of polyphenols from sweet potato (Ipomoea batatas L.) leaves by AB-8 macroporous resins. Food Chem. 2015, 172, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Yanaka, K.; Takebayashi, J.; Matsumoto, T.; Ishimi, Y. Determination of 15 isoflavone isomers in soy foods and supplements by high-performance liquid chromatography. J. Agric. Food Chem. 2012, 60, 4012–4016. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-W.; Ho, C.-T. Antioxidant Properties of Polyphenols Extracted from Green and Black Teas. J. Food Lipids 1995, 2, 35–46. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yagiz, Y.; Buran, T.J.; Nunes, C.D.N.; Gu, L. Phytochemicals from berries and grapes inhibited the formation of advanced glycation end-products by scavenging reactive carbonyls. Food Res. Int. 2011, 44, 2666–2673. [Google Scholar] [CrossRef]
- Kong, Y.; Li, X.; Zheng, T.; Lv, L. Glycation of beta-lactoglobulin and antiglycation by genistein in different reactive carbonyl model systems. Food Chem. 2015, 183, 36–42. [Google Scholar] [CrossRef]
- Singh, H.; Singh, S.; Srivastava, A.; Tandon, P.; Bharti, P.; Kumar, S.; Maurya, R. Conformational analysis and vibrational study of daidzein by using FT-IR and FT-Raman spectroscopies and DFT calculations. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 120, 405–415. [Google Scholar] [CrossRef]
- Huang, Z.; Zhao, M.; Cui, C.; Huang, M.; Lin, L.; Feng, Y. A new sight on soy isoflavones during the whole soy sauce fermentation process by UPLC-MS/MS. LWT 2021, 152, 112249. [Google Scholar] [CrossRef]
- Koh, E.; Mitchell, A.E. Characterization of Urinary Isoflavone Metabolites Excreted after the Consumption of Soy Flour or Soybean Paste Using Lc-(Esi)Ms/Ms. J. Food Biochem. 2011, 35, 1474–1485. [Google Scholar] [CrossRef]
- Kim, D.O.; Lee, C.Y. Comprehensive study on vitamin C equivalent antioxidant capacity (VCEAC) of various polyphenolics in scavenging a free radical and its structural relationship. Crit. Rev. Food Sci. Nutr. 2004, 44, 253–273. [Google Scholar] [CrossRef] [PubMed]
- Lee, C. Relative antioxidant activity of soybean isoflavones and their glycosides. Food Chem. 2005, 90, 735–741. [Google Scholar] [CrossRef]
- Chen, P.; Sun, J.; Liang, Z.; Xu, H.; Du, P.; Li, A.; Meng, Y.; Reshetnik, E.I.; Liu, L.; Li, C. The bioavailability of soy isoflavones in vitro and their effects on gut microbiota in the simulator of the human intestinal microbial ecosystem. Food Res. Int. 2022, 152, 110868. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liu, S.; Liu, Z.; Song, F.; Liu, S. Superoxide generated by pyrogallol reduces highly water-soluble tetrazolium salt to produce a soluble formazan: A simple assay for measuring superoxide anion radical scavenging activities of biological and abiological samples. Anal. Chim. Acta 2013, 793, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Genova, V.M.; Fernandes, A.C.F.; Hiramatsu, É.; Queirós, L.D.; Macedo, J.A.; Macedo, G.A. Biotransformed Antioxidant isoflavone extracts present high-capacity to attenuate the in vitro formation of advanced glycation end products. Food Biotechnol. 2021, 35, 50–66. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, P.; Sang, S. Dietary Genistein Inhibits Methylglyoxal-Induced Advanced Glycation End Product Formation in Mice Fed a High-Fat Diet. J. Nutr. 2019, 149, 776–787. [Google Scholar] [CrossRef]
- McLellan, A.C.; Phillips, S.A.; Thornalley, P.J. The assay of methylglyoxal in biological systems byderivatization with 1,2-diamino-4,5-dimethoxybenzene. Anal. Biochem. 1992, 206, 17–23. [Google Scholar] [CrossRef]
- Wang, P.; Chen, H.; Sang, S. Trapping Methylglyoxal by Genistein and Its Metabolites in Mice. Chem. Res. Toxicol. 2016, 29, 406–414. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein (%) | Fat (%) | Carbohydrate (%) | Ash (%) | Total Isoflavones (mg/kg) | ABTS 1 | Superoxide Anion 2 | |
|---|---|---|---|---|---|---|---|
| Before fermentation | 40.80 ± 0.13 b | 15.75 ± 0.21 b | 7.43 ± 0.32 b | 4.46 ± 0.39 a | 1487.23 ± 34.22 b | 17.15 ± 0.24% b | n.d. 3 |
| After fermentation | 43.25 ± 0.25 a | 20.85 ± 0.07 a | 27.33 ± 1.65 a | 4.40 ± 0.4 a | 2402.65 ± 55.87 a | 44.27 ± 1.27% a | 25 4 |
| Peaks | TR a (min) | Compounds | Formulas | Formula Weight | Precursor Ions [M + H]+ | Exact Mass [M + H]+ | Mass Error b | λmax (nm) | Fragment Ions (m/z) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 11.13 | daidzein-4′-glucoside | C21H20O9 | 416 | 417.11783 | 417.11037 | 1.78 | 250.0, 305.0 | 417.11673/255.06509 |
| 2 | 12.72 | daidzein-7-glucoside | C21H20O9 | 416 | 417.11795 | 417.11037 | 1.79 | 250.0, 304.0 | 417.11658/255.06525 |
| 3 | 13.54 | glycitin | C22H22O10 | 446 | 447.12851 | 447.12130 | 1.61 | 259.2, 319.8 | 447.12766/285.07574 |
| 4 | 15.36 | genistein-4′-glucoside | C21H20O10 | 432 | 433.11310 | 433.10565 | 1.72 | 262.0, 327.5 | 433.11438/271.06021 |
| 5 | 18.42 | genistein-7-glucoside | C21H20O10 | 432 | 433.11298 | 433.10565 | 1.69 | 262.0, 326.0 | 433.11438/271.06021 |
| 6 | 23.10 | 6″-O-succinyldaidzin | C25H24O12 | 516 | 517.13416 | 517.12678 | 1.43 | 249.7 | 517.13409/255.06514 |
| 7 | 25.33 | 6″-O-acetyldaidzin | C23H22O10 | 458 | 459.12869 | 459.12130 | 1.61 | 249.7 | 459.12933/255.06514 |
| 8 | 27.53 | 6″-O-acetylglycitin | C24H24O11 | 488 | 489.13922 | 489.13186 | 1.50 | 260.3, 317.4 | 489.13754/285.07571 |
| 9 | 31.51 | 6″-O-succinyl-4′-genistin | C25H24O13 | 532 | 533.12921 | 533.12169 | 1.41 | 262.0, 326.0 | 533.12982/271.06030 |
| 10 | 33.86 | daidzein | C15H10O4 | 254 | 255.06509 | 255.05791 | 2.82 | 249.7, 301.9 | 255.06511/- |
| 11 | 36.35 | 6″-O-succinyl-7-genistin | C25H24O13 | 532 | 533.12933 | 533.12169 | 1.43 | 262.0, 325.0 | 533.12946/271.06021 |
| 12 | 37.55 | glycitein | C16H12O5 | 284 | 285.07587 | 285.06847 | 2.59 | 255.2, 318.4 | 285.07581/- |
| 13 | 42.88 | 6″-O-acetylgenistin | C23H22O11 | 474 | 475.12372 | 475.11621 | 1.58 | 260.3, 328.1 | 475.12369/271.06012 |
| 14 | 59.93 | genistein | C15H10O5 | 270 | 271.06006 | 271.05282 | 2.67 | 260.3 | 271.060188/- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, A.; Wang, J.; Xie, B.; Hu, K.; Chen, M.; Sun, Z. Determination of 14 Isoflavone Isomers in Natto by UPLC-ESI-MS/MS and Antioxidation and Antiglycation Profiles. Foods 2022, 11, 2229. https://doi.org/10.3390/foods11152229
Xiang A, Wang J, Xie B, Hu K, Chen M, Sun Z. Determination of 14 Isoflavone Isomers in Natto by UPLC-ESI-MS/MS and Antioxidation and Antiglycation Profiles. Foods. 2022; 11(15):2229. https://doi.org/10.3390/foods11152229
Chicago/Turabian StyleXiang, Aoli, Jingyi Wang, Bijun Xie, Kai Hu, Mengting Chen, and Zhida Sun. 2022. "Determination of 14 Isoflavone Isomers in Natto by UPLC-ESI-MS/MS and Antioxidation and Antiglycation Profiles" Foods 11, no. 15: 2229. https://doi.org/10.3390/foods11152229