
Lacticaseibacillus paracasei as a Modulator of Fatty Acid Compositions and Vitamin D3 in Cream

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Obtaining Lacticaseibacillus paracasei Biomass
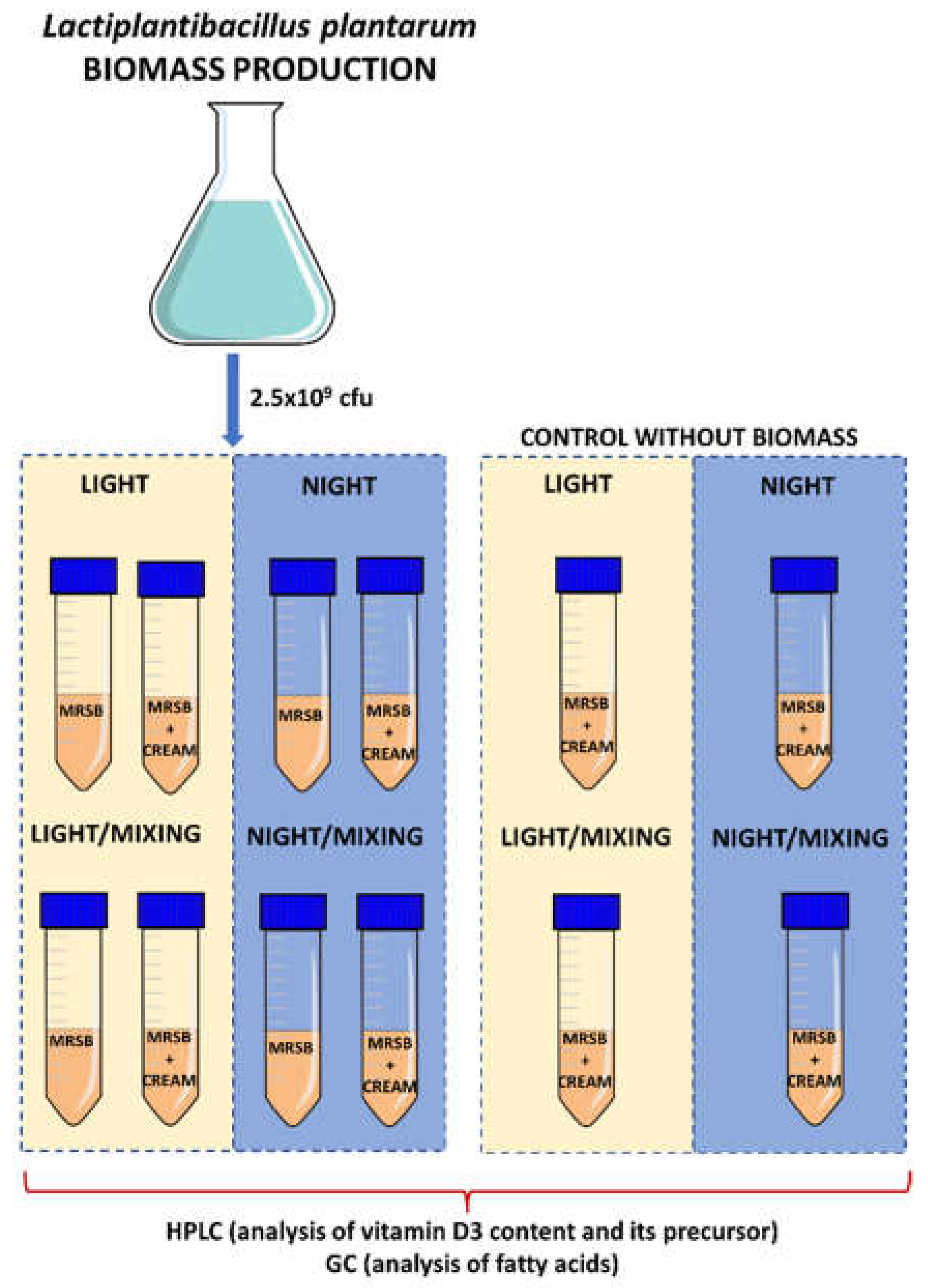
2.3. Experimental Variants
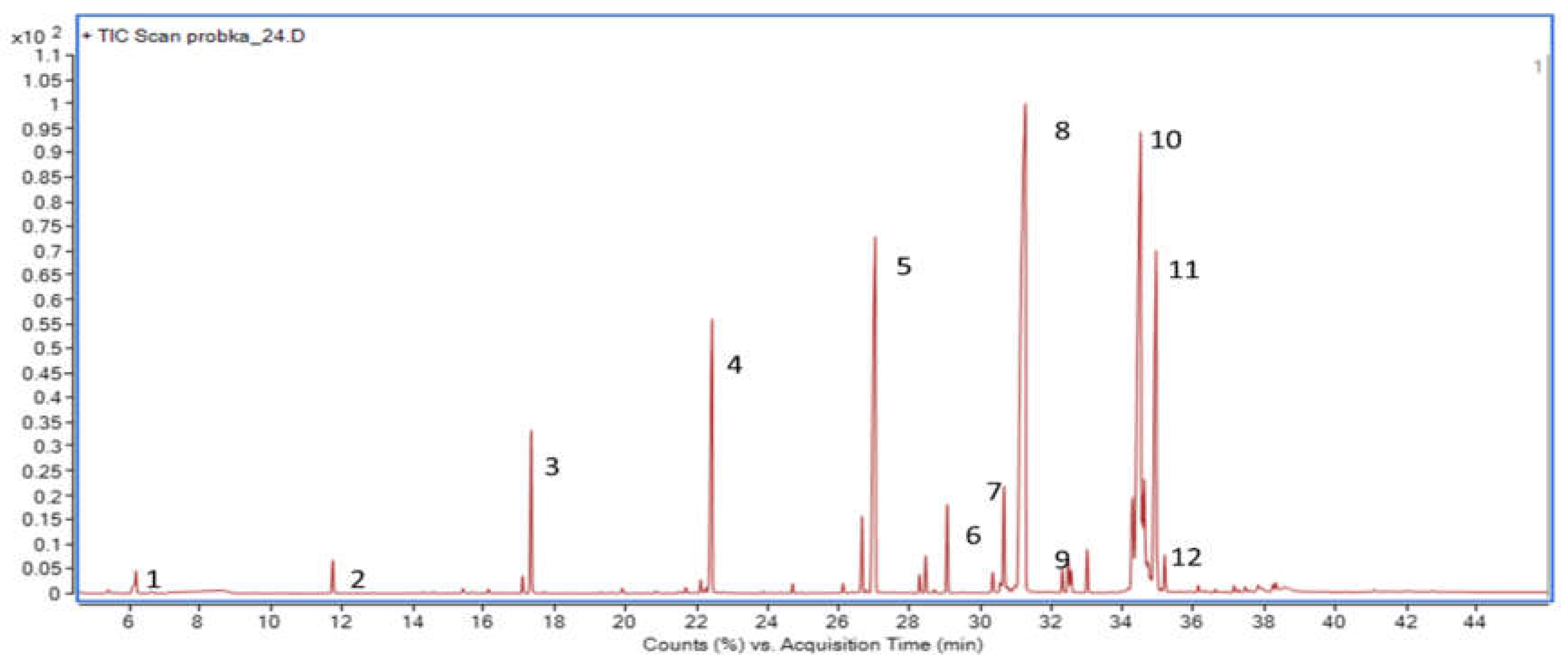
2.4. Sample Preparation and Analysis Procedure for Determination of Fatty Acid Profiles by GC–MS
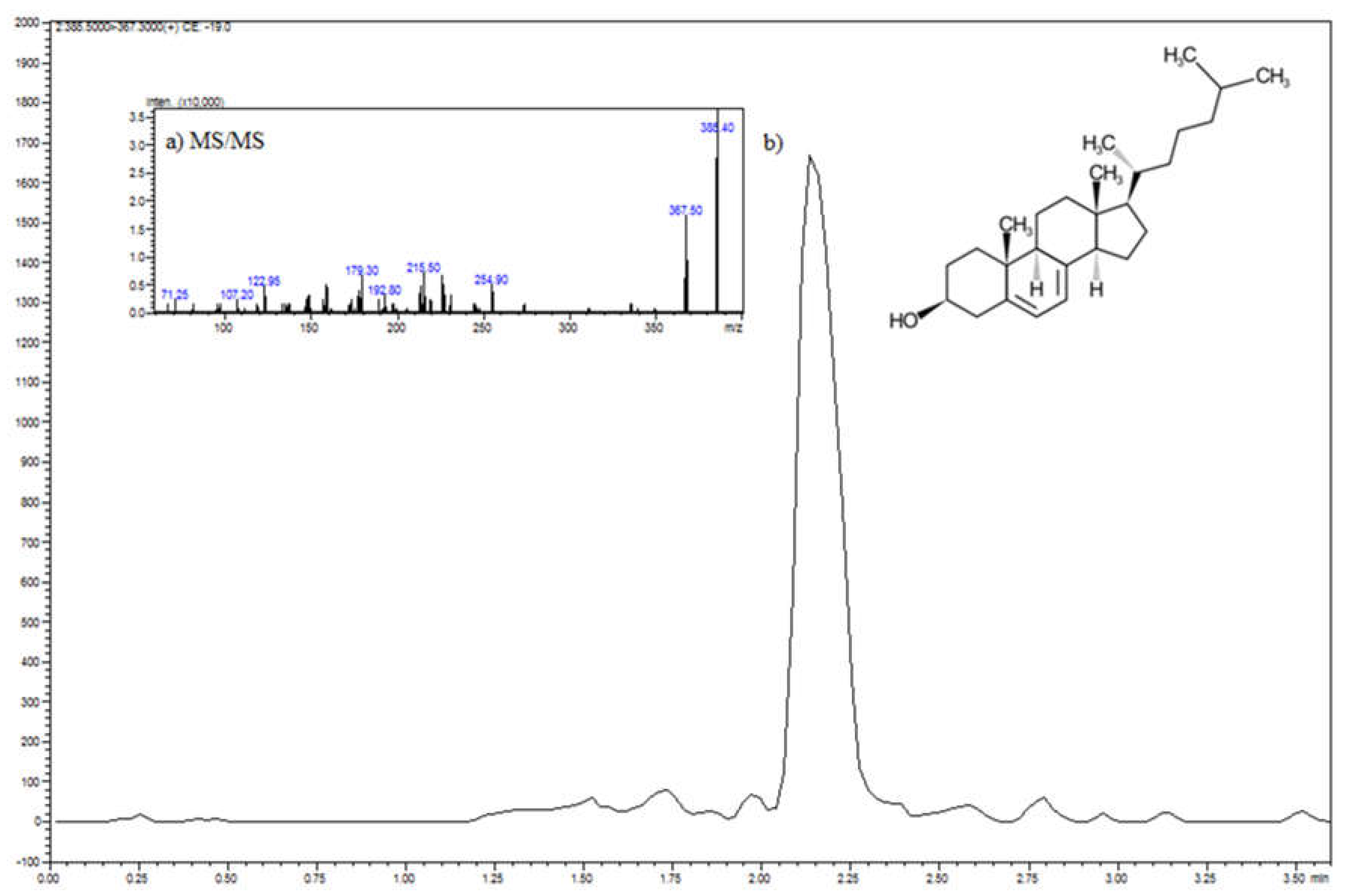
2.5. Sample Preparation Procedure and Determination of Vitamin D3 and 7-Dehydrocholesterol Level by HPLC
3. Results and Discussion
3.1. Impact of the L. paracasei Biomass on the Cream Fatty Acid Composition
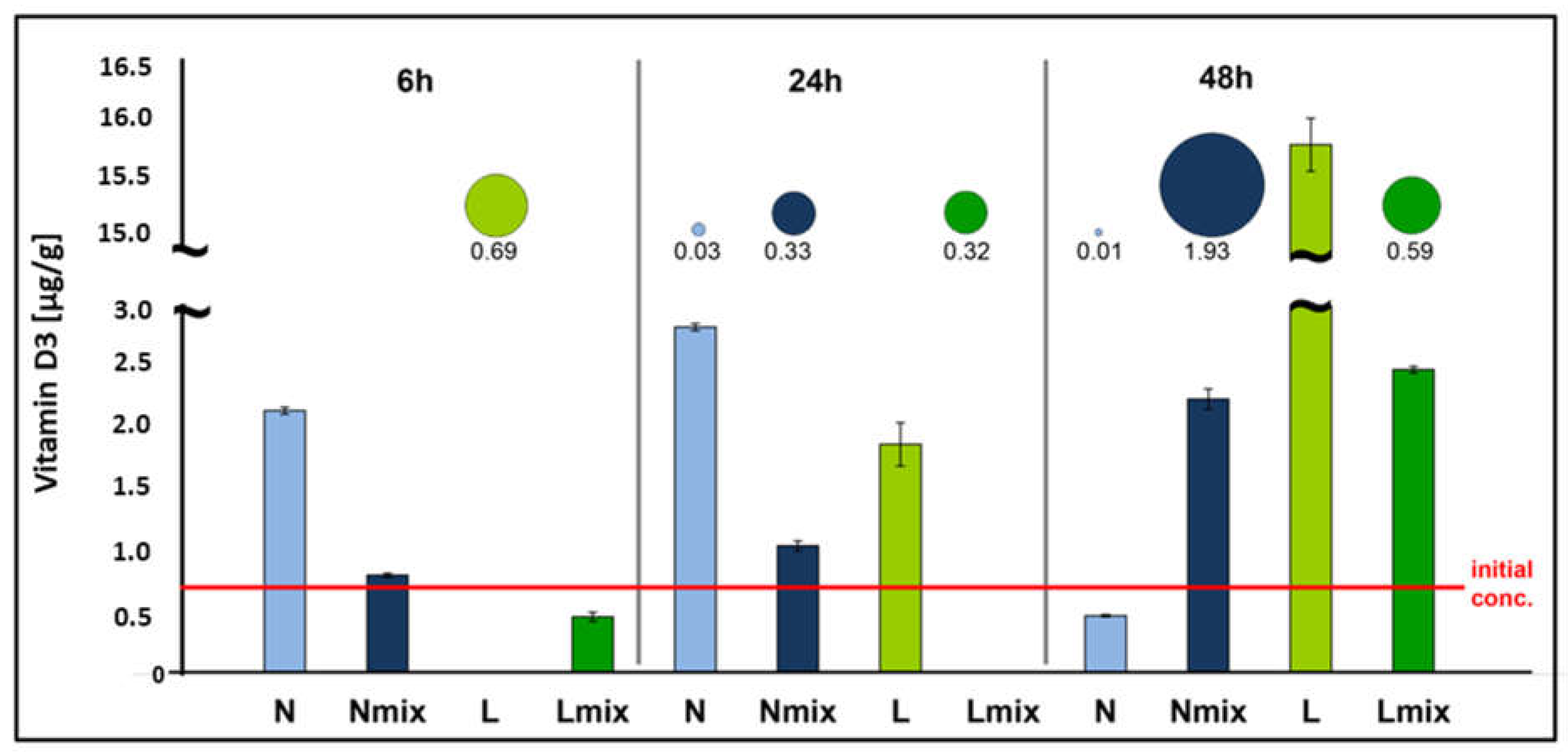
3.2. Impact of the L. paracasei Biomass on Vitamin D3 and Its Precursor Content in Cream
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yilmaz-Ersan, L.; Ozcan, T.; Akpinar-Bayizit, A.; Turan, M.A.; Taskin, M.B. Probiotic cream: Viability of probiotic bacteria and chemical characterization. J. Food Process. Preserv. 2017, 41, e12797. [Google Scholar] [CrossRef]
- Ferreira, L.; Borges, A.; Gomes, D.; Dias, S.; Pereira, C.; Henriques, M. Adding value and innovation in dairy SMEs: From butter to probiotic butter and buttermilk. J. Food Process. Preserv. 2020, e14867. [Google Scholar] [CrossRef]
- World Health Organization. Food and Agriculture Organization of the United Nations Probiotics in food health and nutritional properties and guidelines for evaluation. FAO Food Nutr. Pap. 2006, 85, 1–50. [Google Scholar]
- Terpou, A.; Papadaki, A.; Lappa, I.K.; Kachrimanidou, V.; Bosnea, L.A.; Kopsahelis, N. Probiotics in food systems: Significance and emerging strategies towards improved viability and delivery of enhanced beneficial value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef] [Green Version]
- Evivie, S.E.; Huo, G.-C.; Oamen Igene, J.; Bian, X. Some current applications, limitations and future perspectives of lactic acid bacteria as probiotics. Food Nutr. Res. 2017, 61, 1318034. [Google Scholar] [CrossRef] [Green Version]
- Karaca, Y.; Gün, İ.; Seydim, A.C.; Guzel-Seydim, Z.B. Proizvodnja i kvaliteta maslaca inokuliranog kefirnom kulturom. Mljekarstvo 2018, 68, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Erkaya, T.; Ürkek, B.; Doğru, Ü.; Çetin, B.; Şengül, M. Probiotic butter: Stability, free fatty acid composition and some quality parameters during refrigerated storage. Int. Dairy J. 2015, 49, 102–110. [Google Scholar] [CrossRef]
- Zahedirad, M.; Asadzadeh, S.; Nikooyeh, B.; Neyestani, T.R.; Khorshidian, N.; Yousefi, M.; Mortazavian, A.M. Fortification aspects of vitamin D in dairy products: A review study. Int. Dairy J. 2019, 94, 53–64. [Google Scholar] [CrossRef]
- Holick, M.F. High prevalence of vitamin D inadequacy and implications for health. Mayo Clin. Proc. 2006, 81, 353–373. [Google Scholar] [CrossRef] [Green Version]
- Golfomitsou, I.; Mitsou, E.; Xenakis, A.; Papadimitriou, V. Development of food grade O/W nanoemulsions as carriers of vitamin D for the fortification of emulsion based food matrices: A structural and activity study. J. Mol. Liq. 2018, 268, 734–742. [Google Scholar] [CrossRef]
- Aloğlu, H.; Öner, Z. Assimilation of cholesterol in broth, cream, and butter by probiotic bacteria. Eur. J. Lipid Sci. Technol. 2006, 108, 709–713. [Google Scholar] [CrossRef]
- Yilmaz-Ersan, L. Fatty acid composition of cream fermented by probiotic bacteria. Mljekarstvo Dairy 2013, 63, 132–139. [Google Scholar]
- Ekinci, F.Y.; Okur, O.D.; Ertekin, B.; Guzel-Seydim, Z. Effects of probiotic bacteria and oils on fatty acid profiles of cultured cream. Eur. J. Lipid Sci. Technol. 2008, 110, 216–224. [Google Scholar] [CrossRef]
- Ewe, J.A.; Loo, S.Y. Effect of cream fermentation on microbiological, physicochemical and rheological properties of L. helveticus-butter. Food Chem. 2016, 201, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Caplice, E.; Fitzgerald, G.F. Food fermentations: Role of microorganisms in food production and preservation. Int. J. Food Microbiol. 1999, 50, 131–149. [Google Scholar] [CrossRef]
- Gokhale, S.; Bhaduri, A. Provitamin D3 modulation through prebiotics supplementation: Simulation based assessment. Sci. Rep. 2019, 9, 19267. [Google Scholar] [CrossRef] [PubMed]
- Rudnicka, J. Nowe spojrzenie na proces technologiczny produktów mleczarskich: Otrzymywanie i charakterystyka masła. Przem. Chem. 2021, 1, 35–46. [Google Scholar] [CrossRef]
- Rodríguez-Alcalá, L.M.; Villar-Tajadura, A.; Juarez, M.; Fontecha, J. Commercial conjugated linoleic acid (CLA) fortified dairy products. In Handbook of Food Fortification and Health, 1st ed.; Preedy, V.R., Srirajaskanthan, R., Patel, V.B., Eds.; Humana Press: New York, NY, USA, 2013; Volume 1, pp. 173–184. [Google Scholar]
- Florence, A.C.R.; Da Silva, R.C.; Do Espírito Santo, A.P.; Gioielli, L.A.; Tamime, A.Y.; De Oliveira, M.N. Increased CLA content in organic milk fermented by bifidobacteria or yoghurt cultures. Dairy Sci. Technol. 2009, 89, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Guzel-Seydim, Z.B.; Seydim, A.C.; Greene, A.K.; Taş, T. Determination of antimutagenic properties of acetone extracted fermented milks and changes in their total fatty acid profiles including conjugated linoleic acids. Int. J. Dairy Technol. 2006, 59, 209–215. [Google Scholar] [CrossRef]
- Hayatsu, H.; Inoue, K.; Ohta, H.; Namba, T.; Togawa, K.; Hayatsu, T.; Makita, M.; Wataya, Y. Inhibition of the mutagenicity of cooked-beef basic fraction by its acidic fraction. Mutat. Res. 1981, 91, 437–442. [Google Scholar] [CrossRef]
- Gonzalez, S.; Duncan, S.E.; O’Keefe, S.F.; Sumner, S.S.; Herbein, J.H. Oxidation and textural characteristics of butter and ice cream with modified fatty acid profiles. J. Dairy Sci. 2003, 86, 70–77. [Google Scholar] [CrossRef] [Green Version]
- McDonald, I.W.; Scott, T.W. Foods of ruminant origin with elevated content of polyunsaturated fatty acids. World Rev. Nutr. Diet. 1977, 26, 144–207. [Google Scholar]
- Lin, M.P.; Sims, C.A.; Staples, C.R.; O’Keefe, S.F. Flavor quality and texture of modified fatty acid high monoene, low saturate butter. Food Res. Int. 1996, 29, 367–371. [Google Scholar] [CrossRef]
- Lin, X.; Xia, Y.; Wang, G.; Yang, Y.; Xiong, Z.; Lv, F.; Zhou, W.; Ai, L. Lactic acid bacteria with antioxidant activities alleviating oxidized oil induced hepatic injury in mice. Front. Microbiol. 2018, 9, 2684. [Google Scholar] [CrossRef]
- Holick, M.F. Sunlight and vitamin D for bone health and prevention of autoimmune diseases, cancers, and cardiovascular disease. Am. J. Clin. Nutr. 2004, 80, 1678S–1688S. [Google Scholar] [CrossRef] [Green Version]
- Hansson, P.; Holven, K.B.; Øyri, L.K.; Brekke, H.K.; Biong, A.S.; Gjevestad, G.O.; Raza, G.S.; Herzig, K.H.; Thoresen, M.; Ulven, S.M. Meals with similar fat content from different dairy products induce different postprandial triglyceride responses in healthy adults: A randomized controlled cross-over trial. J. Nutr. 2019, 149, 422–431. [Google Scholar] [CrossRef] [Green Version]
- Kiani, A.; Fathi, M.; Ghasemi, S.M. Production of novel vitamin D3 loaded lipid nanocapsules for milk fortification. Int. J. Food Prop. 2017, 20, 2466–2476. [Google Scholar] [CrossRef] [Green Version]
- Kiełczewska, K.; Jankowska, A.; Dąbrowska, A.; Wachowska, M.; Ziajka, J. The effect of high pressure treatment on the dispersion of fat globules and the fatty acid profile of caprine milk. Int. Dairy J. 2020, 102, 104607. [Google Scholar] [CrossRef]
- Westermann, S.; Brüggemann, D.A.; Olsen, K.; Skibsted, L.H. Light-induced formation of free radicals in cream cheese. Food Chem. 2009, 116, 974–981. [Google Scholar] [CrossRef]
- Sakharov, D.V.; Elstak, E.D.R.; Chernyak, B.; Wirtz, K.W.A. Prolonged lipid oxidation after photodynamic treatment. Study with oxidation-sensitive probe C11-BODIPY581/591. FEBS Lett. 2005, 579, 1255–1260. [Google Scholar] [CrossRef] [Green Version]
- Boswell, C.D.; Nienow, A.W.; Hewitt, C.J. Studies on the effect of mechanical agitation on the performance of brewing fermentations: Fermentation rate, yeast physiology, and development of flavor compounds. J. Am. Soc. Brew. Chem. 2002, 60, 101–106. [Google Scholar] [CrossRef]
- Nienow, A.W.; Mcleod, G.; Hewitt, C.J. Studies supporting the use of mechanical mixing in large scale beer fermentations. Biotechnol. Lett. 2010, 32, 623–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
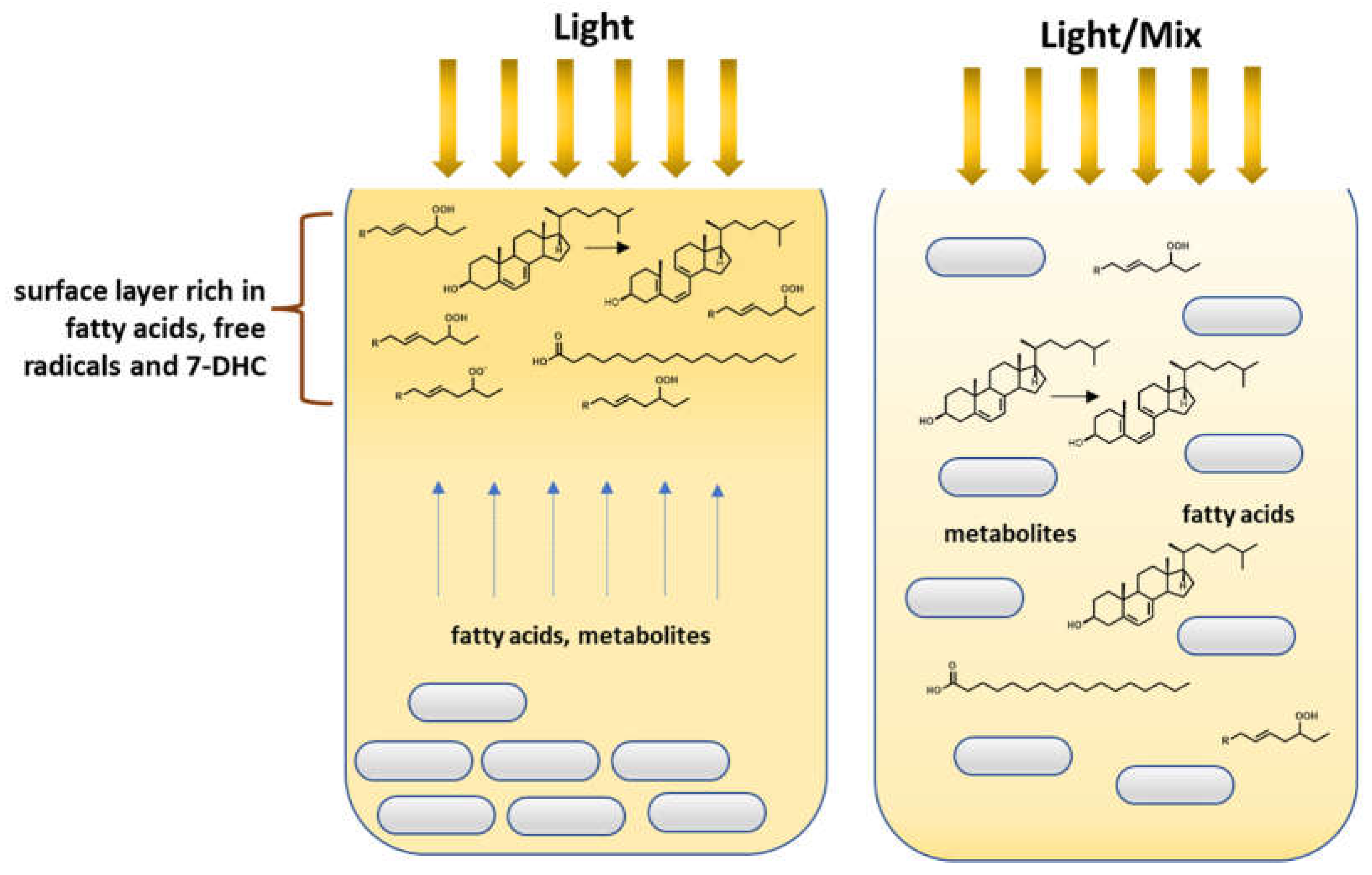{kind=link}
| Fatty Acids [%] | ||||||
|---|---|---|---|---|---|---|
| Variant | SFA | Regulation vs. Control | MUFA | Regulation vs. Control | PUFA | Regulation vs. Control |
| Cream | 66.84 | 30.53 | 2.63 | |||
| MRSB | 95.56 | 4.44 | 0.00 | |||
| LP in MRSB, night | 77.87 |  | 4.04 | | 18.08 |  |
| LP in MRSB, night/mix | 96.20 | | 3.80 | | 0.00 | |
| LP in MRSB, light | 69.32 | | 27.10 | | 3.58 | |
| LP in MRSB, light/mix | 93.42 | | 6.10 | | 0.47 | |
| Cream:MRSB—3:1, night | 68.74 | 27.82 | 3.44 | |||
| Cream:MRSB—3:1, night/mix | 62.38 | 35.98 | 1.63 | |||
| Cream:MRSB—3:1, light | 80.38 | 19.20 | 0.43 | |||
| Cream:MRSB—3:1, light/mix | 78.02 | 19.83 | 2.15 | |||
| LP in Cream:MRSB—3:1, night | 65.92 | | 30.17 | | 3.92 | |
| LP in Cream:MRSB—3:1, night/mix | 94.10 | | 5.90 | | 0.00 | |
| LP in Cream:MRSB—3:1, light | 77.03 | | 22.97 | | 0.00 | |
| LP in Cream:MRSB—3:1, light/mix | 70.92 | | 25.98 | | 3.10 | |
| Conditions with the highest impact on the FA content | ||||||
| SFA ↓ | MUFA ↑ | PUFA ↑ | ||||
| LP in MRSB | light | light | night | |||
| LP in Cream:MRSB | light/mix | light/mix | light/mix | |||
decrease in fatty acid content compared to control samples (MRSB and Cream:MRSB—3:1); increase in fatty acid content compared to control samples (MRSB and Cream:MRSB—3:1).| Variant | MUFA [%] | PUFA [%] | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C12:1 | RSD | C16:1 | RSD | C17:1 | RSD | C18:1 | RSD | C20:1 | RSD | C18:2 | RSD | C20:2 | RSD | C20:3 | RSD | C20:4 | RSD | ||
| Cream | 0.74 ± 0.015 | 2.02 | 4.41 ± 0.0009 | 0.02 | 1.15 ± 0.019 | 1.65 | 24.24 ± 0.36 | 1.49 | 0.25 ± 0.008 | 3.2 | 1.69 ± 0.055 | 3.25 | 0.94 ± 0.016 | 1.7 | - | - | |||
| MRSB | - | - | - | 4.44 ± 0.25 | 5.6 | - | - | - | - | - | |||||||||
| LP in MRSB | night | - | 0.31 ± 0.01 | 3.23 | 0.79 ± 0.019 ** | 2.4 | 2.95 ± 0.17 | 5.76 | - | 18.08 ± 0.66 ** | 3.65 | - | - | - | |||||
| night/mix | - | - | - | 3.8 ± 0.21 | 5.52 | - | - | - | - | - | |||||||||
| light | - | 7.91 ± 0.19 ** | 2.4 | - | 19.2 ± 0.28 ** | 1.45 | - | - | - | - | - | ||||||||
| light/mix | - | 0.26 ± 0.02 | 7.69 | - | 5.84 ± 0.12 | 2.05 | - | 0.47 ± 0.009 | 1.92 | - | - | - | |||||||
| Cream | night | - | 1.99 ± 0.018 | 0.9 | 0.39 ± 0.011 | 2.82 | 25.15 ± 0.18 | 0.72 | 0.3 ± 0.006 | 2.00 | 3.16 ± 0.022 | 0.70 | 0.76 ± 0.018 | 2.37 | - | - | |||
| night/mix | 0.36 ± 0.013 | 3.61 | 4.85 ± 0.023 | 0.47 | 1.02 ± 0.017 | 1.66 | 29.76 ± 0.28 | 0.94 | 0.67 ± 0.009 | 1.34 | - | 1.25 ± 0.019 | 1.52 | 0.21 ± 0.007 | 3.33 | 0.18 ± 0.011 | 6.11 | ||
| light | 0.39 ± 0.009 | 2.31 | 1.61 ± 0.015 | 0.93 | 0.27 ± 0.005 | 1.85 | 16.93 ± 0.014 | 0.08 | - | 0.43 ± 0.008 | 1.86 | - | - | - | |||||
| light/mix | 0.72 ± 0.012 | 1.66 | 8.78 ± 0.17 | 1.94 | 1.46 ± 0.019 | 1.30 | 8.87 ± 0.16 | 1.80 | 1.02 ± 0.016 | 1.57 | - | 2.15 ± 0.025 | 1.16 | - | - | ||||
| LP in Cream | night | 0.35 ± 0.01 ** | 2.86 | 2.57 ± 0.008 ** | 0.3 | 0.49 ± 0.006 ** | 1.22 | 26.75 ± 0.15 ** | 0.56 | - ** | 3.16 ± 0.21 | 6.65 | 0.76 ± 0.013 - ** | 17.11 | - | ||||
| night/mix | - ** | - ** | - ** | 5.90 ± 0.09 ** | 1.53 | - ** | - | - | - ** | - ** | |||||||||
| light | - ** | 1.88 ± 0.19 ** | 1.00 | 0.37 ± 0.008 ** | 2.16 | 20.72 ± 0.15 ** | 0.72 | - | - ** | - | - | - | |||||||
| light/mix | - ** | 2.41 ± 0.25 ** | 10.37 | 0.42 ± 0.009 ** | 2.14 | 23.15 ± 0.21 ** | 0.91 | - ** | 2.39 ± 0.10 ** | 4.18 | 0.71 ± 0.016 ** | 2.25 | - | - | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Złoch, M.; Rafińska, K.; Sugajski, M.; Buszewska-Forajta, M.; Walczak-Skierska, J.; Railean, V.; Pomastowski, P.; Białczak, D.; Buszewski, B. Lacticaseibacillus paracasei as a Modulator of Fatty Acid Compositions and Vitamin D3 in Cream. Foods 2022, 11, 1659. https://doi.org/10.3390/foods11111659
Złoch M, Rafińska K, Sugajski M, Buszewska-Forajta M, Walczak-Skierska J, Railean V, Pomastowski P, Białczak D, Buszewski B. Lacticaseibacillus paracasei as a Modulator of Fatty Acid Compositions and Vitamin D3 in Cream. Foods. 2022; 11(11):1659. https://doi.org/10.3390/foods11111659
Chicago/Turabian StyleZłoch, Michał, Katarzyna Rafińska, Mateusz Sugajski, Magdalena Buszewska-Forajta, Justyna Walczak-Skierska, Viorica Railean, Paweł Pomastowski, Dorota Białczak, and Bogusław Buszewski. 2022. "Lacticaseibacillus paracasei as a Modulator of Fatty Acid Compositions and Vitamin D3 in Cream" Foods 11, no. 11: 1659. https://doi.org/10.3390/foods11111659