
A Review on the Structure and Anti-Diabetic (Type 2) Functions of β-Glucans


Abstract
:1. Introduction
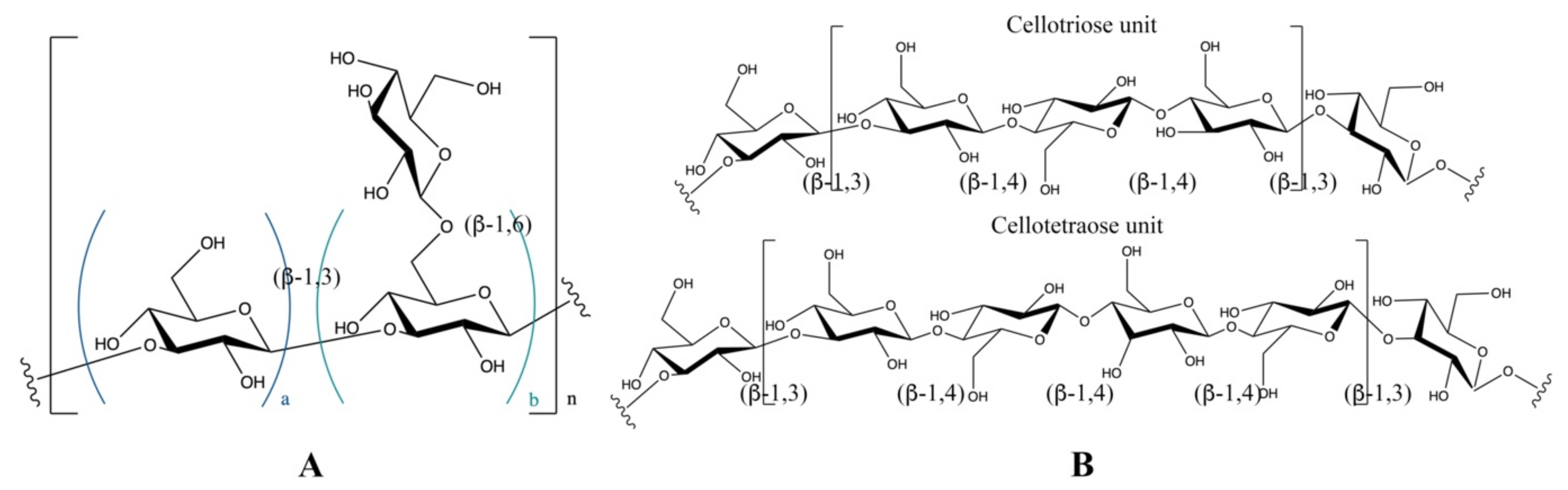
2. Structural Features of β-Glucans
3. Conformational Features of β-Glucans
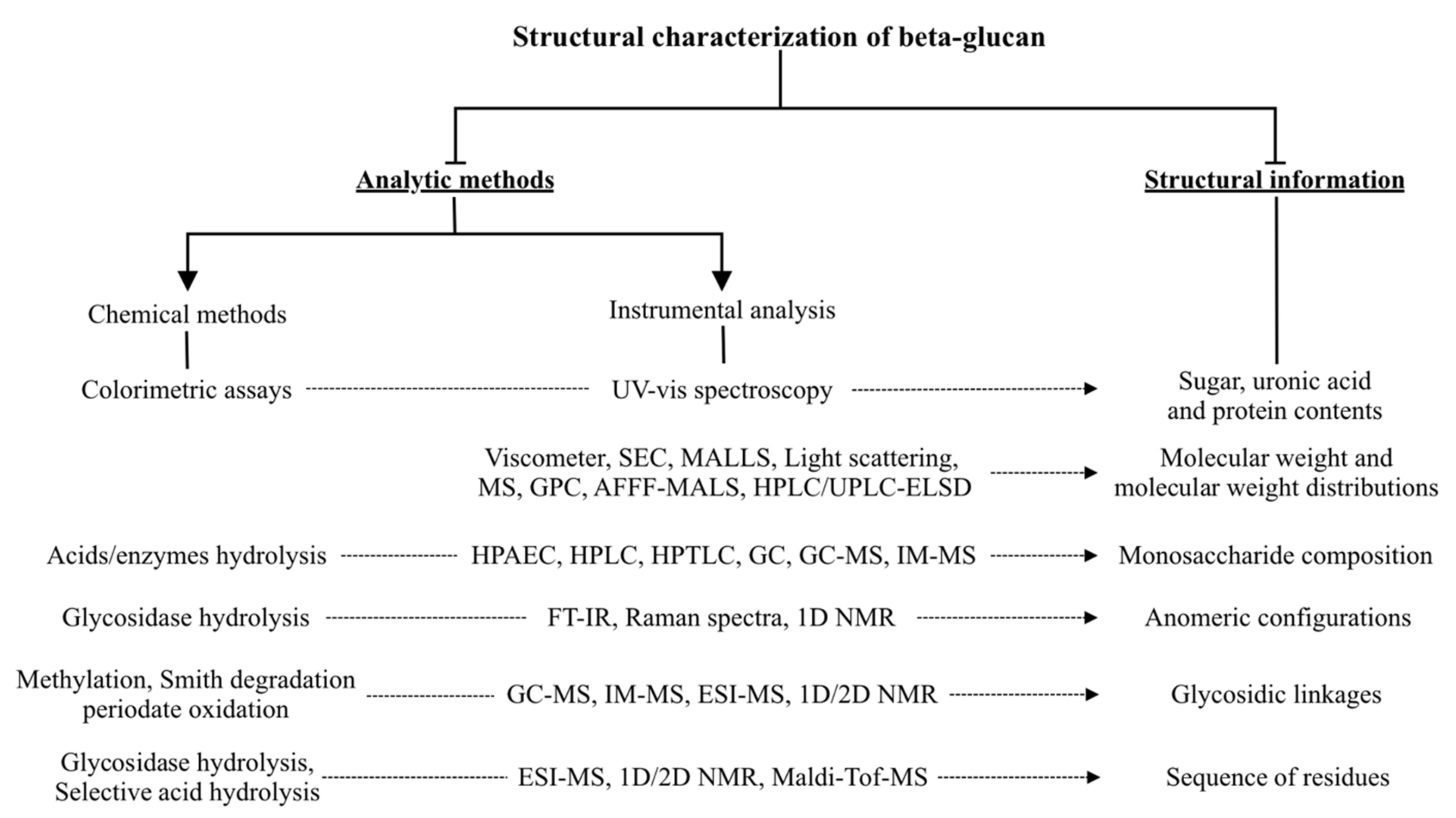
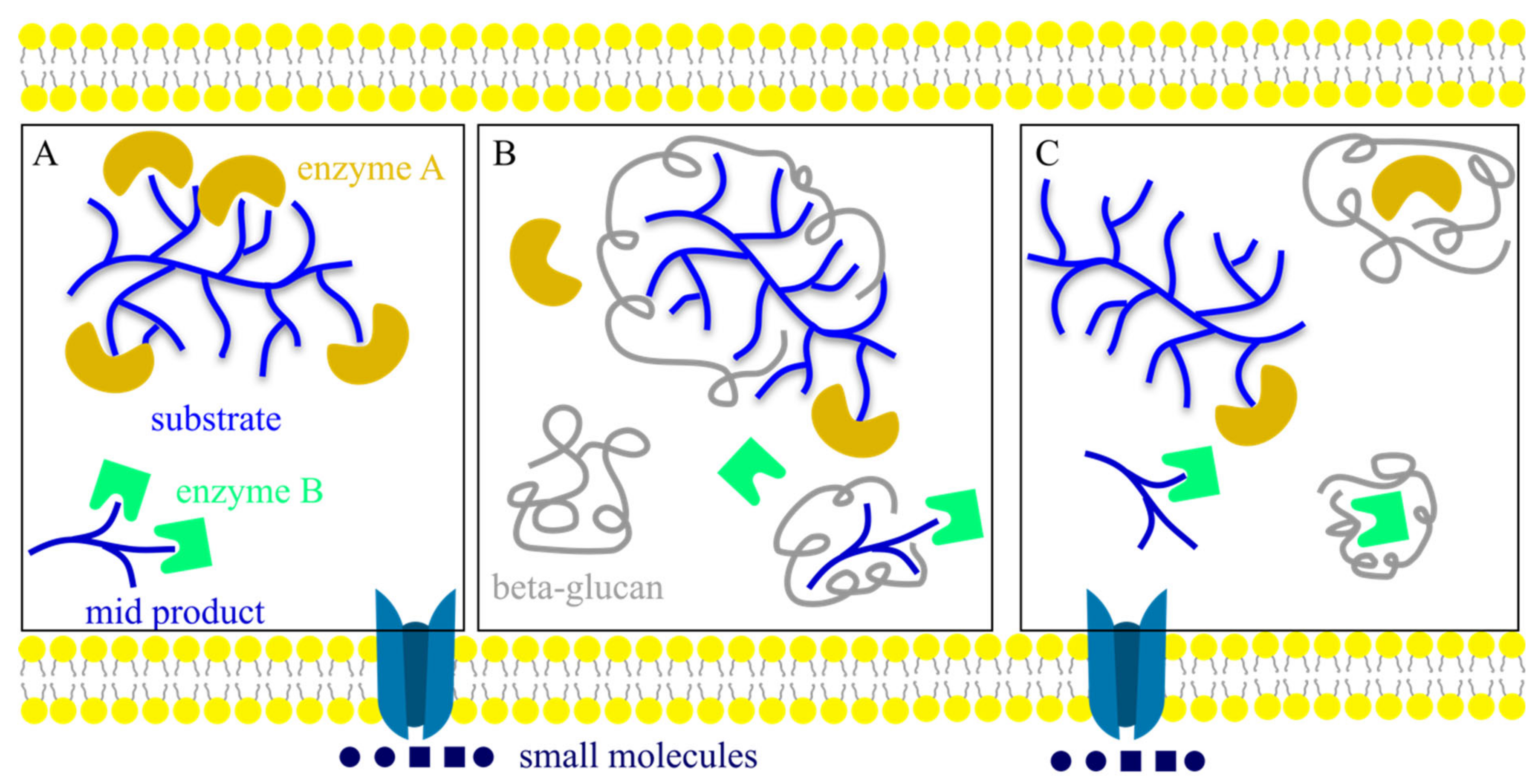
4. Characterization Methods for β-Glucan Structure and Conformation Analysis
5. Amelioration of Type 2 Diabetes and Associated Mechanisms
5.1. Pharmacotherapy for T2D and Anti-Diabetic Mechanisms
5.2. Glucans Used in Controlling T2D and Underlying Mechanisms
5.2.1. Retardation of Macronutrient Absorption
5.2.2. Inhibition of Digestive Enzyme
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Association, A.D. Diagnosis and classification of diabetes mellitus. Diabetes Care 2014, 37, S81–S90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, N.; Shaw, J.; Karuranga, S.; Huang, Y.; da Rocha Fernandes, J.; Ohlrogge, A.; Malanda, B. IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract. 2018, 138, 271–281. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation. IDF Diabetes Atlas, 9th ed.; International Diabetes Federation: Brussels, Belgium, 2019. [Google Scholar]
- Hu, Y.; Ding, B.; Shen, Y.; Yan, R.-N.; Li, F.-F.; Sun, R.; Jing, T.; Lee, K.-O.; Ma, J.-H. Rapid changes in serum testosterone in men with newly diagnosed type 2 diabetes with intensive insulin and metformin. Diabetes Care 2021, 44, 1059–1061. [Google Scholar] [CrossRef] [PubMed]
- Ling, S.; Zaccardi, F.; Lawson, C.; Seidu, S.I.; Davies, M.J.; Khunti, K. Glucose control, sulfonylureas, and insulin treatment in elderly people with type 2 diabetes and risk of severe hypoglycemia and death: An observational study. Diabetes Care 2021, 44, 915–924. [Google Scholar] [CrossRef]
- Han, S.; Hagan, D.L.; Taylor, J.R.; Xin, L.; Meng, W.; Biller, S.A.; Wetterau, J.R.; Washburn, W.N.; Whaley, J.M. Dapagliflozin, a selective SGLT2 inhibitor, improves glucose homeostasis in normal and diabetic rats. Diabetes 2008, 57, 1723–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.; Huh, J.H.; Lee, M.; Lee, Y.H.; Lee, B.W. Glycaemic control with add-on thiazolidinedione or a sodium-glucose co-transporter-2 inhibitor in patients with type 2 diabetes after the failure of an oral triple antidiabetic regimen: A 24-week, randomized controlled trial. Diabetes Obes. Metab. 2021, 23, 609–618. [Google Scholar] [CrossRef]
- Group, U.P.D.S. Effect of intensive blood-glucose control with metformin on complications in overweight patients with type 2 diabetes (UKPDS 34). Lancet 1998, 352, 854–865. [Google Scholar]
- Bergenstal, R.; Lewin, A.; Bailey, T.; Chang, D.; Gylvin, T.; Roberts, V.; Group, N.M.-v.-E.S. Efficacy and safety of biphasic insulin aspart 70/30 versus exenatide in subjects with type 2 diabetes failing to achieve glycemic control with metformin and a sulfonylurea. Curr. Med. Res. Opin. 2009, 25, 65–75. [Google Scholar] [CrossRef]
- Mafra, D.; Borges, N.A.; Lindholm, B.; Shiels, P.G.; Evenepoel, P.; Stenvinkel, P. Food as medicine: Targeting the uraemic phenotype in chronic kidney disease. Nat. Rev. Nephrol. 2020, 17, 153–171. [Google Scholar] [CrossRef]
- Cui, L.; Chen, L.; Yang, G.; Li, Y.; Qiao, Z.; Liu, Y.; Meng, Y.; Zhou, Y.; Sun, L. Structural characterization and immunomodulatory activity of a heterogalactan from Panax ginseng flowers. Food Res. Int. 2021, 140, 109859. [Google Scholar] [CrossRef]
- Wan, Y.; Shi, H.; Xu, R.; Yin, J.; Nie, S.; Xiong, T.; Xie, M. Origin of hypoglycemic benefits of probiotic-fermented carrot pulp. J. Agric. Food Chem. 2019, 67, 895–904. [Google Scholar] [CrossRef]
- Song, Q.; Wang, Y.; Huang, L.; Shen, M.; Yu, Y.; Yu, Q.; Chen, Y.; Xie, J. Review of the relationships among polysaccharides, gut microbiota, and human health. Food Res. Int. 2020, 140, 109858. [Google Scholar] [CrossRef] [PubMed]
- Chihara, G.; Hamuro, J.; Maeda, Y.Y.; Arai, Y.; Fukuoka, F. Fractionation and purification of the polysaccharides with marked antitumor activity, especially lentinan, from Lentinus edodes (Berk.) Sing.(an edible mushroom). Cancer Res. 1970, 30, 2776–2781. [Google Scholar]
- Chihara, G.; Maeda, Y.; Hamuro, J.; Sasaki, T.; Fukuoka, F. Inhibition of mouse sarcoma 180 by polysaccharides from Lentinus edodes (Berk.) sing. Nature 1969, 222, 687–688. [Google Scholar] [CrossRef]
- El Enshasy, H.A.; Hatti-Kaul, R. Mushroom immunomodulators: Unique molecules with unlimited applications. Trends Biotechnol. 2013, 31, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Xie, J.; Xue, B.; Li, X.; Gan, J.; Zhu, T.; Sun, T. Effect of acid and oxidative degradation on the structural, rheological, and physiological properties of oat β-glucan. Food Hydrocoll. 2021, 112, 106284. [Google Scholar] [CrossRef]
- Sun, T.; Li, J.; Qin, Y.; Xie, J.; Xue, B.; Li, X.; Gan, J.; Bian, X.; Shao, Z. Rheological and functional properties of oat β-glucan with different molecular weight. J. Mol. Struct. 2020, 1209, 127944. [Google Scholar] [CrossRef]
- Xiao, J.B.; Jiang, H. A review on the structure-function relationship aspect of polysaccharides from tea materials. Crit. Rev. Food Sci. Nutr. 2015, 55, 930–938. [Google Scholar] [CrossRef]
- Meng, Y.; Lyu, F.; Xu, X.; Zhang, L. Recent advances in chain conformation and bioactivities of triple-helix polysaccharides. Biomacromolecules 2020, 21, 1653–1677. [Google Scholar] [CrossRef]
- Mejia, S.M.V.; de Francisco, A.; Bohrer, B.M. A comprehensive review on cereal beta-glucan: Extraction, characterization, causes of degradation, and food application. Crit. Rev. Food Sci. Nutr. 2020, 61, 3693–3704. [Google Scholar] [CrossRef]
- Wang, J.T.; Xu, X.J.; Zheng, H.; Li, J.L.; Deng, C.; Xu, Z.H.; Chen, J.H. Structural characterization, chain conformation, and morphology of a beta-(1 – >3)-D-glucan isolated from the fruiting body of dictyophora indusiata. J. Agric. Food Chem. 2009, 57, 5918–5924. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Hu, Z.; Fu, H.; Hu, M.; Xu, X.; Chen, J. Chemical analysis and antioxidant activity in vitro of a beta-D-glucan isolated from Dictyophora indusiata. Int. J. Biol. Macromol. 2012, 51, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Yang, B.; Tang, J.; Ma, Z.; Gao, Q.; Zhao, M. Structural analysis of water-soluble polysaccharides in the fruiting body of Dictyophora indusiata and their in vivo antioxidant activities. Carbohydr. Polym. 2012, 87, 343–347. [Google Scholar] [CrossRef]
- Zhang, R.; Edgar, K.J. Properties, chemistry, and applications of the bioactive polysaccharide curdlan. Biomacromolecules 2014, 15, 1079–1096. [Google Scholar] [CrossRef]
- Harada, T.; Misaki, A.; Saito, H. Curdlan—A Bacterial gel-forming beta-1→3-glucan. Arch. Biochem. Biophys. 1968, 124, 292. [Google Scholar] [CrossRef]
- Ahmad, A.; Anjum, F.M.; Zahoor, T.; Nawaz, H.; Ahmed, Z. Extraction and characterization of β-d-glucan from oat for industrial utilization. Int. J. Biol. Macromol. 2010, 46, 304–309. [Google Scholar] [CrossRef]
- Gamel, T.H.; Abdel-Aal, E.-S.M.; Ames, N.P.; Duss, R.; Tosh, S.M. Enzymatic extraction of beta-glucan from oat bran cereals and oat crackers and optimization of viscosity measurement. J. Cereal Sci. 2014, 59, 33–40. [Google Scholar] [CrossRef]
- Okobira, T.; Miyoshi, K.; Uezu, K.; Sakurai, K.; Shinkai, S. Molecular dynamics studies of side chain effect on the β-1, 3-D-glucan triple helix in aqueous solution. Biomacromolecules 2008, 9, 783–788. [Google Scholar] [CrossRef]
- Zhong, K.; Tong, L.; Liu, L.; Zhou, X.; Liu, X.; Zhang, Q.; Zhou, S. Immunoregulatory and antitumor activity of schizophyllan under ultrasonic treatment. Int. J. Biol. Macromol. 2015, 80, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, S.; Wang, X.; Zhang, L.; Cheung, P.C.K. Advances in lentinan: Isolation, structure, chain conformation and bioactivities. Food Hydrocoll. 2011, 25, 196–206. [Google Scholar] [CrossRef]
- Meng, Y.; Shi, X.; Cai, L.; Zhang, S.; Ding, K.; Nie, S.; Luo, C.; Xu, X.; Zhang, L. Triple-helix conformation of a polysaccharide determined with light scattering, afm, and molecular dynamics simulation. Macromolecules 2018, 51, 10150–10159. [Google Scholar] [CrossRef]
- Xu, S.; Lin, Y.; Huang, J.; Li, Z.; Xu, X.; Zhang, L. Construction of high strength hollow fibers by self-assembly of a stiff polysaccharide with short branches in water. J. Mater. Chem. A 2013, 1, 4198–4206. [Google Scholar] [CrossRef]
- Ping, Z.H.; Xu, H.; Liu, T.; Huang, J.C.; Meng, Y.; Xu, X.J.; Li, W.H.; Zhang, L.N. Anti-hepatoma activity of the stiff branched beta-D-glucan and effects of molecular weight. J. Mat. Chem. B 2016, 4, 4565–4573. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Jia, L.M.; Fang, J.N. A beta-D-glucan isolated from the fruiting bodies of Hericium erinaceus and its aqueous conformation. Carbohydr. Res. 2006, 341, 791–795. [Google Scholar] [CrossRef]
- Wu, D.; Tang, C.; Liu, Y.; Li, Q.; Wang, W.; Zhou, S.; Zhang, Z.; Cui, F.; Yang, Y. Structural elucidation and immunomodulatory activity of a beta-D-glucan prepared by freeze-thawing from Hericium erinaceus. Carbohydr. Polym. 2019, 222, 114996. [Google Scholar] [CrossRef]
- Cui, H.; Zhu, X.; Huo, Z.; Liao, B.; Huang, J.; Wang, Z.; Song, C.; Hu, X.; Fang, J. A beta-glucan from Grifola frondosa effectively delivers therapeutic oligonucleotide into cells via dectin-1 receptor and attenuates TNFalpha gene expression. Int. J. Biol. Macromol. 2020, 149, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Iino, K.; Ohno, N.; Suzuki, I.; Miyazaki, T.; Yadomae, T.; Oikawa, S.; Sato, K. Structural characterisation of a neutral antitumour β-d-glucan extracted with hot sodium hydroxide from cultured fruit bodies of Grifola frondosa. Carbohydr. Res. 1985, 141, 111–119. [Google Scholar] [CrossRef]
- Fang, J.; Wang, Y.; Lv, X.; Shen, X.; Ni, X.; Ding, K. Structure of a β-glucan from Grifola frondosa and its antitumor effect by activating Dectin-1/Syk/NF-κB signaling. Glycoconj. J. 2012, 29, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-R.; Hwang, J.; Koh, H.-J.; Jang, K.; Lee, J.-D.; Choi, J.; Yang, C.-S. The targeted delivery of the c-Src peptide complexed with schizophyllan to macrophages inhibits polymicrobial sepsis and ulcerative colitis in mice. Biomaterials 2016, 89, 1–13. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, structure and biofunctional activities of laminarin from brown algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Shin, H.J.; Oh, S.J.; Kim, S.I.; Won Kim, H.; Son, J.-H. Conformational characteristics of β-glucan in laminarin probed by terahertz spectroscopy. Appl. Phys. Lett. 2009, 94, 111911. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, X.; Zhang, L. Gel formation and low-temperature intramolecular conformation transition of a triple-helical polysaccharide lentinan in water. Biopolymers 2008, 89, 852–861. [Google Scholar] [CrossRef]
- Kao, P.F.; Wang, S.H.; Hung, W.T.; Liao, Y.H.; Lin, C.M.; Yang, W.B. Structural characterization and antioxidative activity of low-molecular-weights beta-1,3-glucan from the residue of extracted ganoderma lucidum fruiting bodies. J. Biomed. Biotechnol. 2012, 2012, 673764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, J.; Tang, Q.; Yang, Y.; Guo, Q.; Wang, Q.; Wu, D.; Cui, S.W. Physicochemical characterization of a high molecular weight bioactive beta-D-glucan from the fruiting bodies of Ganoderma lucidum. Carbohydr. Polym. 2014, 101, 968–974. [Google Scholar] [CrossRef]
- Aimanianda, V.; Clavaud, C.; Simenel, C.; Fontaine, T.; Delepierre, M.; Latgé, J.-P. Cell wall β-(1, 6)-glucan of Saccharomyces cerevisiae: Structural characterization and in situ synthesis. J. Biol. Chem. 2009, 284, 13401–13412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chen, S.; Nie, S.; Cui, S.W.; Wang, Q.; Phillips, A.O.; Phillips, G.O.; Xie, M. Structural characterization and chain conformation of water-soluble β-glucan from wild cordyceps sinensis. J. Agric. Food Chem. 2019, 67, 12520–12527. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Wood, P. Relationships Between Structural Features, Molecular Weight and Rheological Properties of Cereal β-D-Glucans. In Hydrocolloids; Elsevier: Amstardam, The Netherlands, 2000; pp. 159–168. [Google Scholar]
- De Arcangelis, E.; Djurle, S.; Andersson, A.A.M.; Marconi, E.; Messia, M.C.; Andersson, R. Structure analysis of β-glucan in barley and effects of wheat β-glucanase. J. Cereal Sci. 2019, 85, 175–181. [Google Scholar] [CrossRef]
- Roubroeks, J.; Andersson, R.; Åman, P. Structural features of (1→3),(1→4)-β-D-glucan and arabinoxylan fractions isolated from rye bran. Carbohydr. Polym. 2000, 42, 3–11. [Google Scholar] [CrossRef]
- Wood, P.; Weisz, J.; Blackwell, B. Molecular characterization of cereal β-D-glucans. Structural analysis of oat β-D-glucan and rapid structural evaluation of β-D-glucans from different sources by high-performance liquid chromatography of oligosaccharides released by lichenase. Cereal Chem. 1991, 68, 31–39. [Google Scholar]
- Wood, P.; Weisz, J.; Blackwell, B. Structural studies of (1→3),(1→4)-β-D-glucans by 13C-nuclear magnetic resonance spectroscopy and by rapid analysis of cellulose-like regions using high-performance anion-exchange chromatography of oligosaccharides released by lichenase. Cereal Chem. 1994, 71, 301–307. [Google Scholar]
- Beer, M.U.; Wood, P.J.; Weisz, J.; Fillion, N. Effect of cooking and storage on the amount and molecular weight of (1→3)(1→4)-β-D-glucan extracted from oat products by an in vitro digestion system. Cereal Chem. 1997, 74, 705–709. [Google Scholar] [CrossRef]
- Yuan, H.J.; Lan, P.; He, Y.; Li, C.L.; Ma, X. Effect of the modifications on the physicochemical and biological properties of beta-glucan—A critical review. Molecules 2020, 25, 57. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Zhou, F.; Xu, X.; Zhang, L. Uptake of intraperitoneally administrated triple helical β-glucan for antitumor activity in murine tumor models. J. Mat. Chem. B 2017, 5, 9337–9345. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Zhang, H. Recent progress on curdlan provided by functionalization strategies. Food Hydrocoll. 2017, 68, 128–135. [Google Scholar] [CrossRef]
- Geller, A.; Shrestha, R.; Yan, J. Yeast-derived beta-glucan in cancer: Novel uses of a traditional therapeutic. Int. J. Mol. Sci. 2019, 20, 618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Lu, F.Z.; Xu, X.J.; Zhang, L.N. Extended chain conformation of beta-glucan and its effect on antitumor activity. J. Mat. Chem. B 2017, 5, 5623–5631. [Google Scholar] [CrossRef]
- Yanaki, T.; Norisuye, T.; Fujita, H. Triple helix of Schizophyllum commune polysaccharide in dilute solution. 3. Hydrodynamic properties in water. Macromolecules 1980, 13, 1462–1466. [Google Scholar] [CrossRef]
- Xu, S.; Xu, X.; Zhang, L. Effect of heating on chain conformation of branched β-glucan in water. J. Phys. Chem. B 2013, 117, 8370–8377. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, L.; Xu, X. Morphologies and conformation transition of lentinan in aqueous NaOH solution. Biopolym. Orig. Res. Biomol. 2004, 75, 187–195. [Google Scholar]
- Xu, X.; Zhang, X.; Zhang, L.; Wu, C. Collapse and association of denatured lentinan in water/dimethlysulfoxide solutions. Biomacromolecules 2004, 5, 1893–1898. [Google Scholar] [CrossRef]
- Edge, C.; Parekh, R.; Rademacher, T.; Wormald, M.; Dwek, R. Fast sequencing of oligosaccharides using arrays of enzymes. Nature 1992, 358, 693–694. [Google Scholar] [CrossRef]
- Venkataraman, G.; Shriver, Z.; Raman, R.; Sasisekharan, R. Sequencing complex polysaccharides. Science 1999, 286, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, J.; Hahm, H.; Seeberger, P.H.; Pagel, K. Identification of carbohydrate anomers using ion mobility–mass spectrometry. Nature 2015, 526, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Rief, M.; Oesterhelt, F.; Heymann, B.; Gaub, H.E. Single molecule force spectroscopy on polysaccharides by atomic force microscopy. Science 1997, 275, 1295–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-Q.; Bruun, L.; Pierson, E.S.; Cresti, M. Periodic deposition of arabinogalactan epitopes in the cell wall of pollen tubes of Nicotiana tabacum L. Planta 1992, 188, 532–538. [Google Scholar] [CrossRef]
- Wu, X.; Delbianco, M.; Anggara, K.; Michnowicz, T.; Pardo-Vargas, A.; Bharate, P.; Sen, S.; Pristl, M.; Rauschenbach, S.; Schlickum, U.; et al. Imaging single glycans. Nature 2020, 582, 375–378. [Google Scholar] [CrossRef]
- Li, B.; Zhang, N.; Feng, Q.; Li, H.; Wang, D.; Ma, L.; Liu, S.; Chen, C.; Wu, W.; Jiao, L. The core structure characterization and of ginseng neutral polysaccharide with the immune-enhancing activity. Int. J. Biol. Macromol. 2019, 123, 713–722. [Google Scholar] [CrossRef]
- Lee, Y.C. Carbohydrate analyses with high-performance anion-exchange chromatography. J. Chromatogr. A 1996, 720, 137–149. [Google Scholar] [CrossRef]
- Kono, H.; Kondo, N.; Isono, T.; Ogata, M.; Hirabayashi, K. Characterization of the secondary structure and order–disorder transition of a β-(1→3, 1→6)-glucan from Aureobasidium pullulans. Int. J. Biol. Macromol. 2020, 154, 1382–1391. [Google Scholar] [CrossRef]
- Roden, M.; Shulman, G.I. The integrative biology of type 2 diabetes. Nature 2019, 576, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, A.; Wessel, J.; Willems, S.M.; Zhao, W.; Robertson, N.R.; Chu, A.Y.; Gan, W.; Kitajima, H.; Taliun, D.; Rayner, N.W. Refining the accuracy of validated target identification through coding variant fine-mapping in type 2 diabetes. Nat. Genet. 2018, 50, 559–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varanko, A.K.; Chilkoti, A. Molecular and materials engineering for delivery of peptide drugs to treat type 2 diabetes. Adv. Healthc. Mater. 2019, 8, 1801509. [Google Scholar] [CrossRef] [PubMed]
- Cabré, C.; Colungo, C.; Vinagre, I.; Jansà, M.; Conget, I. Frequency and awareness of hypoglycemia in patients with Type 2 Diabetes treated with two or more insulin injections in primary care outpatient clinics. Prim. Care Diabetes 2020, 14, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Esteve, E.; Tremaroli, V.; Khan, M.T.; Caesar, R.; Mannerås-Holm, L.; Ståhlman, M.; Olsson, L.M.; Serino, M.; Planas-Fèlix, M. Metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes, contributing to the therapeutic effects of the drug. Nat. Med. 2017, 23, 850. [Google Scholar] [CrossRef]
- Cabreiro, F.; Au, C.; Leung, K.-Y.; Vergara-Irigaray, N.; Cochemé, H.M.; Noori, T.; Weinkove, D.; Schuster, E.; Greene, N.D.; Gems, D. Metformin retards aging in C. elegans by altering microbial folate and methionine metabolism. Cell 2013, 153, 228–239. [Google Scholar] [CrossRef] [Green Version]
- Nauck, M.A.; Meininger, G.; Sheng, D.; Terranella, L.; Stein, P.P.; Sitagliptin Study Group. Efficacy and safety of the dipeptidyl peptidase-4 inhibitor, sitagliptin, compared with the sulfonylurea, glipizide, in patients with type 2 diabetes inadequately controlled on metformin alone: A randomized, double-blind, non-inferiority trial. Diabetes Obes. Metab. 2007, 9, 194–205. [Google Scholar] [CrossRef]
- Proks, P.; Reimann, F.; Green, N.; Gribble, F.; Ashcroft, F. Sulfonylurea stimulation of insulin secretion. Diabetes 2002, 51, S368–S376. [Google Scholar] [CrossRef] [Green Version]
- Pajvani, U.B.; Hawkins, M.; Combs, T.P.; Rajala, M.W.; Doebber, T.; Berger, J.P.; Wagner, J.A.; Wu, M.; Knopps, A.; Xiang, A.H. Complex distribution, not absolute amount of adiponectin, correlates with thiazolidinedione-mediated improvement in insulin sensitivity. J. Biol. Chem. 2004, 279, 12152–12162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, A.; Kohan, D.E.; Nelson, R.D.; Gonzalez, F.J.; Yang, T. Collecting duct-specific deletion of peroxisome proliferator-activated receptor γ blocks thiazolidinedione-induced fluid retention. Proc. Natl. Acad. Sci. USA 2005, 102, 9406–9411. [Google Scholar] [CrossRef] [Green Version]
- Drucker, D.J. Biological actions and therapeutic potential of the glucagon-like peptides. Gastroenterology 2002, 122, 531–544. [Google Scholar] [CrossRef] [Green Version]
- Bettge, K.; Kahle, M.; Abd El Aziz, M.S.; Meier, J.J.; Nauck, M.A. Occurrence of nausea, vomiting and diarrhoea reported as adverse events in clinical trials studying glucagon-like peptide-1 receptor agonists: A systematic analysis of published clinical trials. Diabetes Obes. Metab. 2017, 19, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Herman, G.A.; Stevens, C.; van Dyck, K.; Bergman, A.; Yi, B.; de Smet, M.; Snyder, K.; Hilliard, D.; Tanen, M.; Tanaka, W. Pharmacokinetics and pharmacodynamics of sitagliptin, an inhibitor of dipeptidyl peptidase IV, in healthy subjects: Results from two randomized, double-blind, placebo-controlled studies with single oral doses. Clin. Pharmacol. Ther. 2005, 78, 675–688. [Google Scholar] [CrossRef] [PubMed]
- Fadini, G.P.; Avogaro, A. Cardiovascular effects of DPP-4 inhibition: Beyond GLP-1. Vasc. Pharmacol. 2011, 55, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Lambeir, A.-M.; Scharpé, S.; de Meester, I. DPP4 inhibitors for diabetes—What next? Biochem. Pharmacol. 2008, 76, 1637–1643. [Google Scholar] [CrossRef]
- Taylor, S.I.; Blau, J.E.; Rother, K.I. SGLT2 inhibitors may predispose to ketoacidosis. J. Clin. Endocrinol. Metab. 2015, 100, 2849–2852. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.; Menichini, F. Natural products as α-amylase and α-glucosidase inhibitors and their hypoglycaemic potential in the treatment of diabetes: An update. Mini Rev. Med. Chem. 2010, 10, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Pino, J.L.; Mujica, V.; Arredondo, M. Effect of dietary supplementation with oat β-glucan for 3 months in subjects with type 2 diabetes: A randomized, double-blind, controlled clinical trial. J. Funct. Foods 2021, 77, 104311. [Google Scholar] [CrossRef]
- Maki, K.; Galant, R.; Samuel, P.; Tesser, J.; Witchger, M.; Ribaya-Mercado, J.; Blumberg, J.; Geohas, J. Effects of consuming foods containing oat β-glucan on blood pressure, carbohydrate metabolism and biomarkers of oxidative stress in men and women with elevated blood pressure. Eur. J. Clin. Nutr. 2007, 61, 786–795. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Yang, C.; Liu, B.; Lin, L.; Sarker, S.D.; Nahar, L.; Yu, H.; Cao, H.; Xiao, J. Bioactive compounds from marine macroalgae and their hypoglycemic benefits. Trends Food Sci. Technol. 2018, 72, 1–12. [Google Scholar] [CrossRef]
- Nishinari, K.; Takemasa, M.; Fang, Y.; Hossain, K.S.; Tsumura, Y.; Sone, Y.; Fujiwara, M.; Habu, D.; Emoto, M. Effects of xyloglucan with different molar masses on glucose in blood. Food Hydrocoll. 2020, 108, 105727. [Google Scholar] [CrossRef]
- Wood, P.; Beer, M.; Butler, G. Evaluation of role of concentration and molecular weight of oat β-glucan in determining effect of viscosity on plasma glucose and insulin following an oral glucose load. Br. J. Nutr. 2000, 84, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Tosh, S.M.; Bordenave, N. Emerging science on benefits of whole grain oat and barley and their soluble dietary fibers for heart health, glycemic response, and gut microbiota. Nutr. Rev. 2020, 78, 13–20. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Zhou, S.; Yan, M.; Tang, Q.; Zhang, J. Structure and chain conformation of bioactive β-D-glucan purified from water extracts of Ganoderma lucidum unbroken spores. Int. J. Biol. Macromol. 2021, 180, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Atluri, S.; Zhang, Z.; Gidley, M.J.; Li, E.; Gilbert, R.G. Structural reasons for inhibitory effects of pectin on α-amylase enzyme activity and in-vitro digestibility of starch. Food Hydrocoll. 2021, 114, 106581. [Google Scholar] [CrossRef]
- Gong, X.; Sui, L.; Morton, J.; Brennan, M.A.; Brennan, C.S. Investigation of nutritional and functional effects of rice bran protein hydrolysates by using Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) guidelines: A review. Trends Food Sci. Technol. 2021, 110, 798–811. [Google Scholar] [CrossRef]
- Ma, B.; Feng, T.; Zhang, S.; Zhuang, H.; Chen, D.; Yao, L. The inhibitory effects of Hericium erinaceus β-glucan on in vitro starch digestion. Front. Nutr. 2020, 7, 348. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.P.; Aluko, R.E.; Hati, S.; Solanki, D. Bioactive peptides in the management of lifestyle-related diseases: Current trends and future perspectives. Crit. Rev. Food Sci. Nutr. 2021, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Gunness, P.; Gidley, M.J. Barley β-glucan effects on emulsification and in vitro lipolysis of canola oil are modulated by molecular size, mixing method, and emulsifier type. Food Hydrocoll. 2020, 103, 105643. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Z.; Tian, Y.; Song, Z.; Ai, L. Interaction between barley β-glucan and corn starch and its effects on the in vitro digestion of starch. Int. J. Biol. Macromol. 2019, 141, 240–246. [Google Scholar] [CrossRef]
- Gong, L.; Feng, D.; Wang, T.; Ren, Y.; Liu, Y.; Wang, J. Inhibitors of α-amylase and α-glucosidase: Potential linkage for whole cereal foods on prevention of hyperglycemia. Food Sci. Nutr. 2020, 8, 6320–6337. [Google Scholar] [CrossRef]
- Fukuhara, G. Analytical supramolecular chemistry: Colorimetric and fluorimetric chemosensors. J. Photochem. Photobiol. C Photochem. Rev. 2020, 42, 100340. [Google Scholar] [CrossRef]
- Tan, H.; Nie, S. Functional hydrocolloids, gut microbiota and health: Picking food additives for personalized nutrition. FEMS Microbiol. Rev. 2021, 45, fuaa065. [Google Scholar] [CrossRef]
- Velikonja, A.; Lipoglavšek, L.; Zorec, M.; Avguštin, G. Alterations in gut microbiota composition and metabolic parameters after dietary intervention with barley beta glucans in patients with high risk for metabolic syndrome development. Anaerobe 2019, 55, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ames, N.P.; Tun, H.M.; Tosh, S.M.; Jones, P.J.; Khafipour, E. High molecular weight barley β-glucan alters gut microbiota toward reduced cardiovascular disease risk. Front. Microbiol. 2016, 7, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Wang, Y.; Yang, G.; Zhang, Q.; Meng, L.; Xin, Y.; Jiang, X. The role of short-chain fatty acids in intestinal barrier function, inflammation, oxidative stress, and colonic carcinogenesis. Pharmacol. Res. 2021, 165, 105420. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Hu, X.; Guo, Q.; Cui, S.W.; Xian, Y.; You, S.; Chen, X.; Xu, C.; Gao, X. Physicochemical properties and regulatory effects on db/db diabetic mice of β-glucans extracted from oat, wheat and barley. Food Hydrocoll. 2014, 37, 60–68. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name/Abbr. | Source | Extraction Solvent | Type of Glucan | Structure a | Ref. |
|---|---|---|---|---|---|
| Curdlan | Alcaligenes faecalis var. | NaOH | β-1,3 glucan | (A) a = 1, b = 0 | [25,26] |
| APP | Auricularia auricula | NaCl | β-1,3 glucan | (A) a = 1, b = 2 | [32,33,34] |
| DIP | Dictyophora indusiata | Water | β-1,3 glucan | (A) a = 2, b = 1 | [22,23,24] |
| HEP | Hericium erinaceus | Water | β-1,3 glucan | (A) a = 2, b = 1 | [35,36] |
| GFP | Grifola frondosa | Water | β-1,3 glucan | (A) a = 2, b = 1 | [37,38,39] |
| Schizophyllan | Schizophyllum | Water | β-1,3 glucan | (A) a = 2, b = 1 | [30,40] |
| Laminarin | Algae | Water | β-1,3 glucan | (A) a = 2, b = 1 | [41,42] |
| Lentinan | Lentinula edodes | NaCl/NaOH | β-1,3 glucan | (A) a = 3, b = 2 | [31,43] |
| GLP | Ganoderma lucidum | Water | β-1,3 glucan | (A) a = 5, b = 1 | [44,45] |
| YBG | Saccharomyces cerevisiae | NaOH | β-1,3 glucan | (A) a = 5, b = 1 | [46] |
| CSP | Wild Cordyceps sinensis | Water | β-1,3 glucan | (A) a = 5, b = 2 | [47] |
| WBG | Wheat | Water | β-1,3-1,4 glucan | (B) DP3:DP4 = 3.0–4.5 | [21,48] |
| BBG | Barley | Water/NaOH | β-1,3-1,4 glucan | (B) DP3:DP4 = 1.7–3.3 | [48,49] |
| RBG | Rye | Water | β-1,3-1,4 glucan | (B) DP3:DP4 = 1.8–3.1 | [50,51,52] |
| OBG | Oat | Water | β-1,3-1,4 glucan | (B) DP3:DP4 = 1.5–2.2 | [48,53] |
| Type | Drug Name | Mechanisms | Side-Effects | Ref. |
|---|---|---|---|---|
| Biguanides | Metformin, Phenformin | Lowering fasting plasma insulin concentration; enhancing insulin sensitivity; changing gut microbiota composition; promoting functional shifts in gut microbiome. | Gastrointestinal disorders; folate deficiency; increasing homocysteine levels | [8,76,77] |
| Sulfonylureas | Glibenclamide, Glipizide | As insulin secretagogues to stimulate insulin secretion. | Gastrointestinal disorders, headache | [9,78,79] |
| Thiazolidinediones | Rosiglitazone, Pioglitazone | Improving insulin sensitivity by up-regulation of adipokine. | Peripheral and pulmonary edema; fluid retention. | [80,81] |
| GLP-1 | Liraglutide, Semaglutide | Suppressing glucagon release; delaying gastric emptying and increasing satiety. | Nausea, vomiting and diarrhoea | [82,83] |
| DPP-4 inhibitors | Vidagliptin, Saxagliptin | Enhancing incretin axis; improving meal-stimulated insulin secretion by sparing incretin hormones. | Nausea and gastrointestinal problems | [78,84,85,86] |
| SGLT2 inhibitors | Dapagliflozin, Cangliflozin | Inhibition of renal glucose reabsorption to lower plasma glucose levers. | Increasing the risk of developing diabetic ketoacidosis. | [6,87] |
| Enzyme inhibitors | α-amylase inhibitors, α-glucosidase inhibitors, | Reduction in the rate of glucose absorption in post-prandial blood | Lactic acidosis, diarrhoea, liver function disorders. | [88] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, Y.; Xu, X.; Gilbert, R.G.; Sullivan, M.A. A Review on the Structure and Anti-Diabetic (Type 2) Functions of β-Glucans. Foods 2022, 11, 57. https://doi.org/10.3390/foods11010057
Wan Y, Xu X, Gilbert RG, Sullivan MA. A Review on the Structure and Anti-Diabetic (Type 2) Functions of β-Glucans. Foods. 2022; 11(1):57. https://doi.org/10.3390/foods11010057
Chicago/Turabian StyleWan, Yujun, Xiaojuan Xu, Robert G. Gilbert, and Mitchell A. Sullivan. 2022. "A Review on the Structure and Anti-Diabetic (Type 2) Functions of β-Glucans" Foods 11, no. 1: 57. https://doi.org/10.3390/foods11010057