
Molecular Analytical Assessment of Thermally Precipitated α-Lactalbumin after Resolubilization


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. α-La Precipitation, Separation, and Refolding Procedure
2.3. Determination of Resolubilization Degree
2.4. Production of Native α-La as Analytical Standard
2.5. Characterization of Elution Time by RP-HPLC Analysis
2.6. Determination of Denaturation Temperature by DSC
2.7. Analysis of the Secondary Structure by FTIR
2.8. Data Evaluation and Statistics
3. Results and Discussion
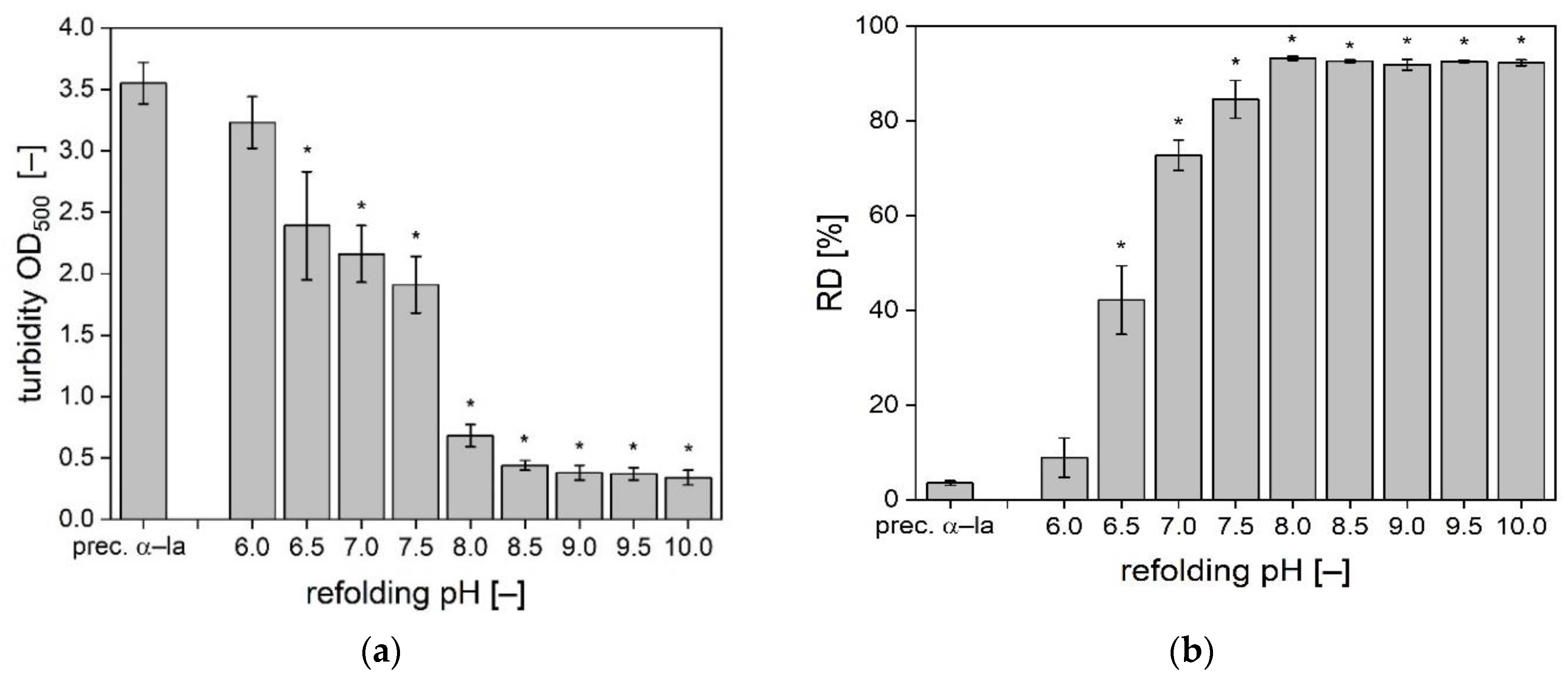
3.1. Solubilization of α-La Precipitates
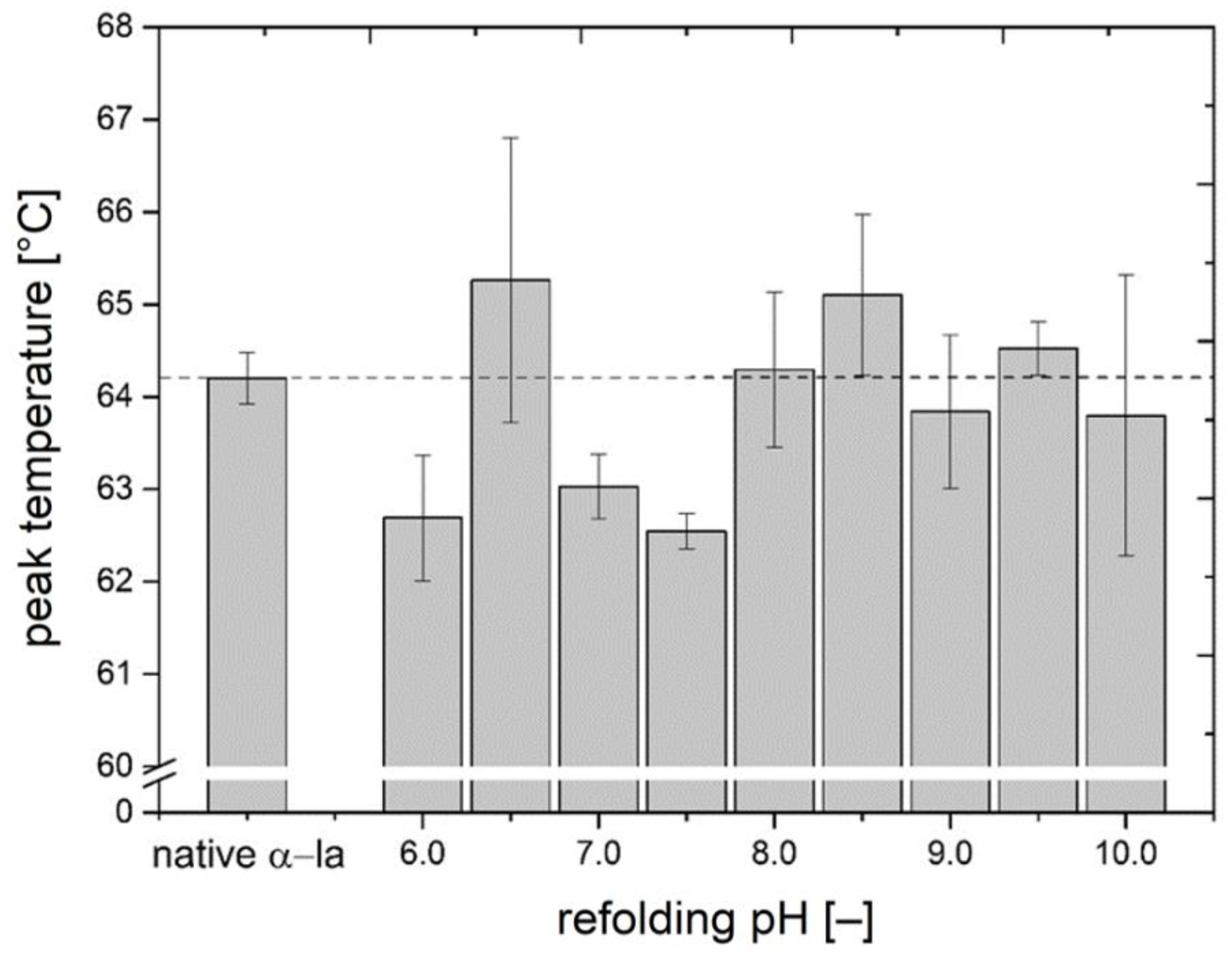
3.2. Investigation of Thermal Stability
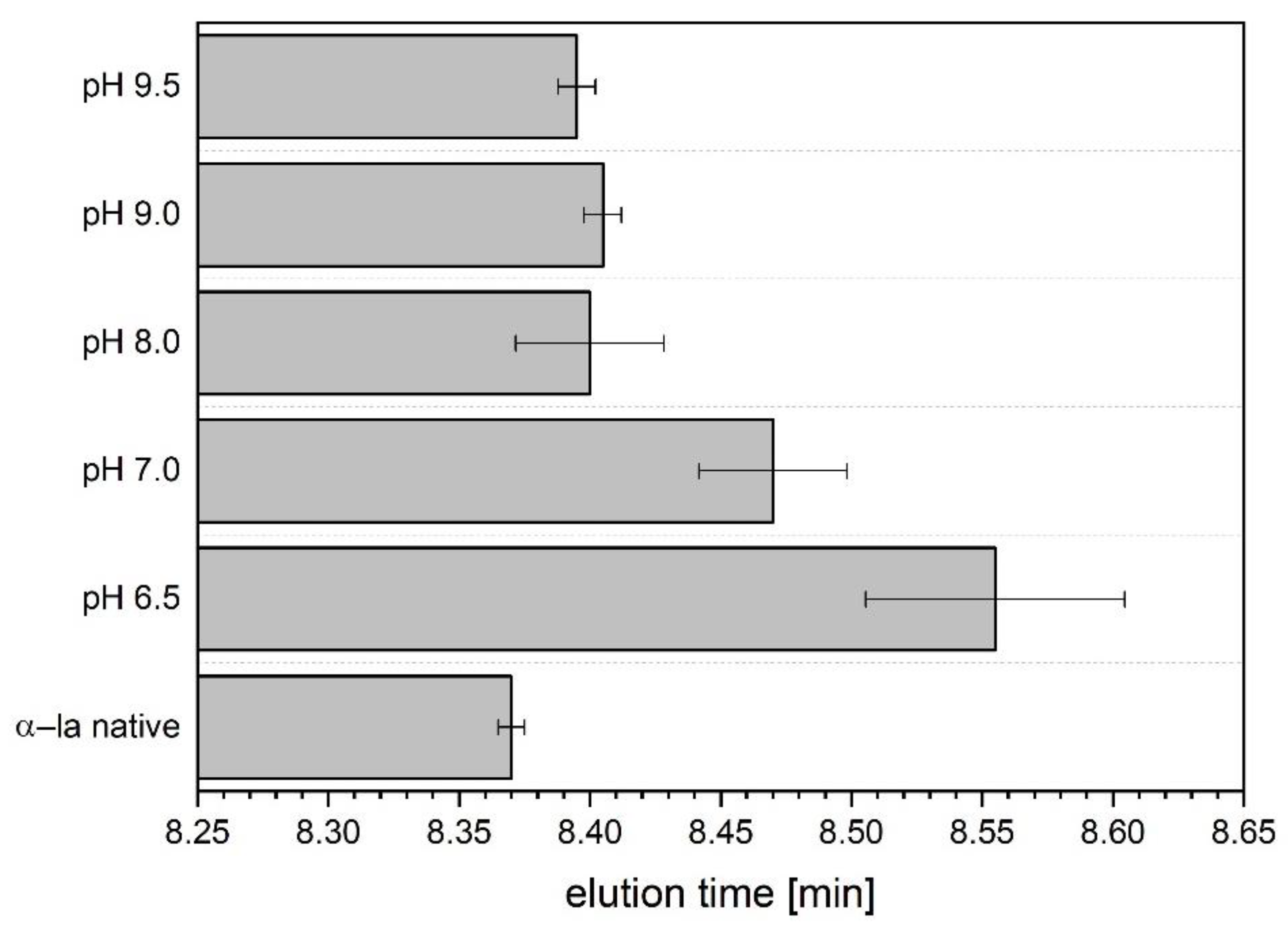
3.3. Investigation of Hydrophobicity
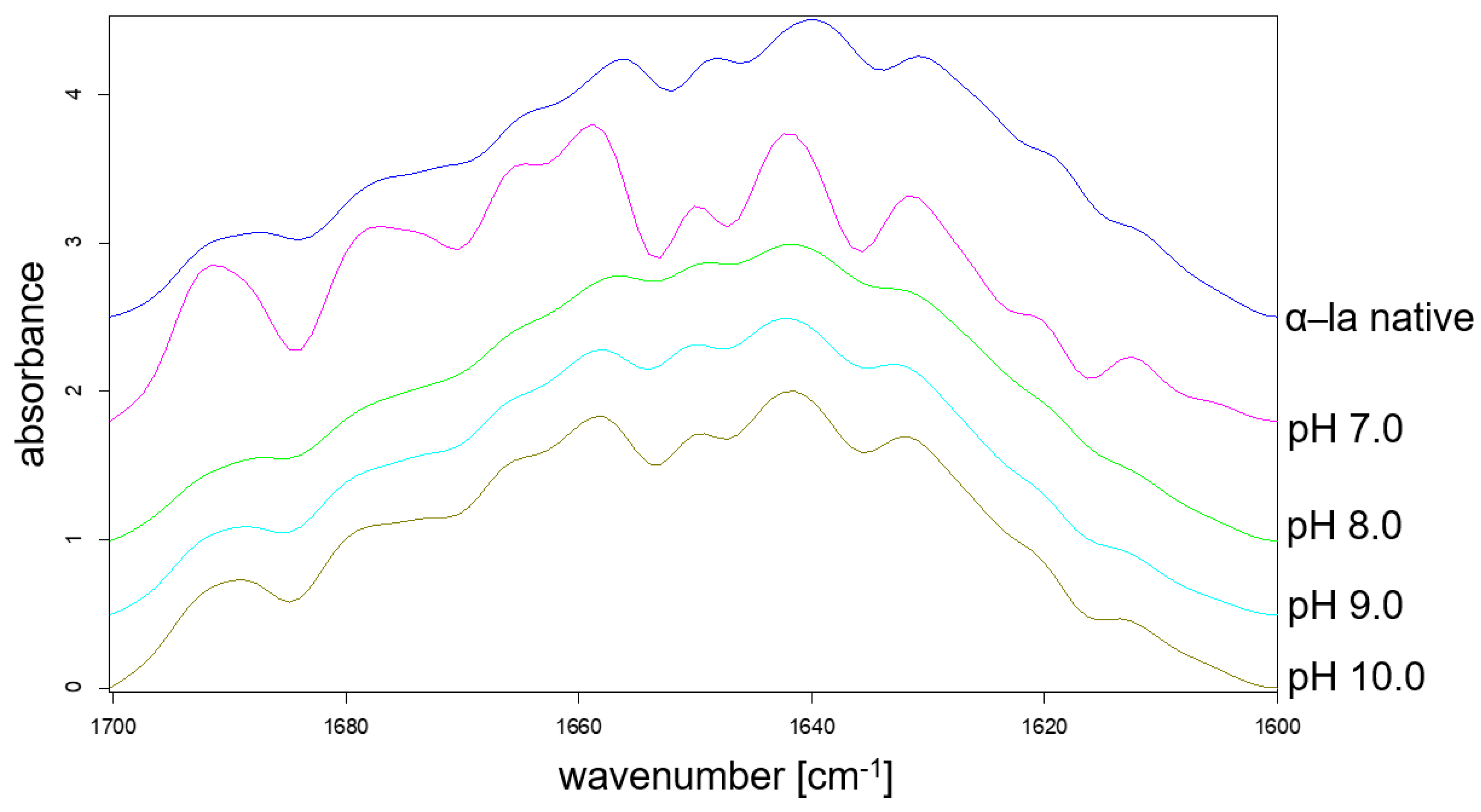
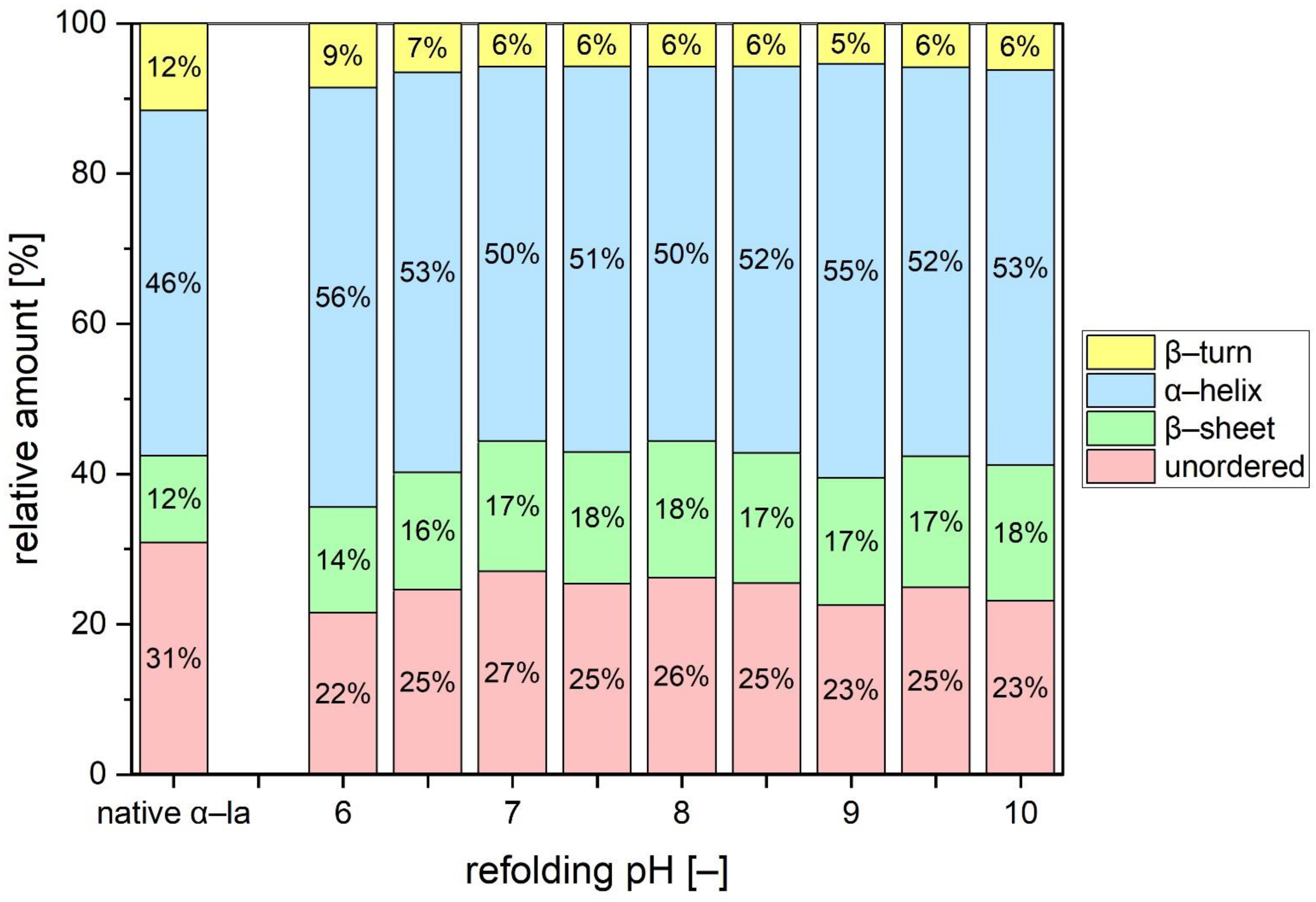
3.4. Analysis of Secondary Structure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muuronen, K.; Partanen, R.; Heidebrecht, H.J.; Kulozik, U. Effects of conventional processing methods on whey proteins in production of native whey powder. Int. Dairy J. 2021, 116, 104959. [Google Scholar] [CrossRef]
- Lien, E.L.; Davis, A.M.; Euler, A.R. Growth and safety in term infants fed reduced-protein formula with added bovine alpha-lactalbumin. J. Pediatr. Gastroenterol. Nutr. 2004, 38, 170–176. [Google Scholar] [CrossRef] [Green Version]
- Chatterton, D.E.; Smithers, G.; Roupas, P.; Brodkorb, A. Bioactivity of β-lactoglobulin and α-lactalbumin-Technological implications for processing. Int. Dairy J. 2006, 16, 1229–1240. [Google Scholar] [CrossRef]
- Permyakov, E.A. α-Lactalbumin, Amazing Calcium-Binding Protein. Biomolecules 2020, 10, 1210. [Google Scholar] [CrossRef]
- Heine, W.E.; Klein, P.D.; Reeds, P.J. The importance of α-lactalbumin in infant nutrition. Nutr. J. 1991, 121, 277–283. [Google Scholar] [CrossRef]
- Stanciuc, N.; Rapeanu, G. An overview of bovine [alpha]-lactalbumin structure and functionality. Ann. Univ. Dunarea Jos Galati Fascicle VI Food Technol. 2010, 34, 82. [Google Scholar]
- Expósito, I.L.; Recio, I. Antibacterial activity of peptides and folding variants from milk proteins. Int. Dairy J. 2006, 16, 1294–1305. [Google Scholar] [CrossRef]
- Delgado, Y.; Morales-Cruz, M.; Hernández-Román, J.; Hernández, G.; Griebenow, K. Development of HAMLET-like cytochrome c-oleic acid nanoparticles for cancer therapy. J. Nanomed. Nanotechnol. 2015, 6, 1. [Google Scholar]
- Hoque, M.; Nanduri, R.; Gupta, J.; Mahajan, S.; Gupta, P.; Saleemuddin, M. Oleic acid complex of bovine α-lactalbumin induces eryptosis in human and other erythrocytes by a Ca2+-independent mechanism. Biochim Biophys Acta Gen. Subj. 2015, 1850, 1729–1739. [Google Scholar] [CrossRef]
- Toro-Sierra, J.; Tolkach, A.; Kulozik, U. Fractionation of α-Lactalbumin and β-Lactoglobulin from Whey Protein Isolate Using Selective Thermal Aggregation, an Optimized Membrane Separation Procedure and Resolubilization Techniques at Pilot Plant Scale. Food Bioprocess Technol. 2013, 6, 1032–1043. [Google Scholar] [CrossRef]
- Patel, S. Emerging trends in nutraceutical applications of whey protein and its derivatives. J. Food Sci. Technol. 2015, 52, 6847–6858. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Cano, F.J.; Marín-Gallén, S.; Castell, M.; Rodríguez-Palmero, M.; Rivero, M.; Franch, A.; Castellote, C. Bovine whey protein concentrate supplementation modulates maturation of immune system in suckling rats. Br. J. Nutr. 2007, 98, S80–S84. [Google Scholar] [CrossRef] [Green Version]
- Prussick, R.; Prussick, L.; Gutman, J. Psoriasis improvement in patients using glutathione-enhancing, nondenatured whey protein isolate: A pilot study. J. Clin. Aesthet. Dermatol. 2013, 6, 23. [Google Scholar] [PubMed]
- Navis, M.; Muncan, V.; Sangild, P.T.; Willumsen, L.M.; Koelink, P.J.; Wildenberg, M.E.; Abrahamse, E.; Thymann, T.; Van Elburg, R.M.; Renes, I.B. Beneficial Effect of Mildly Pasteurized Whey Protein on Intestinal Integrity and Innate Defense in Preterm and Near-Term Piglets. Nutrients 2020, 12, 1125. [Google Scholar] [CrossRef] [Green Version]
- Navis, M.; Schwebel, L.; Kappel, S.S.; Muncan, V.; Sangild, P.T.; Abrahamse, E.; Aunsholt, L.; Thymann, T.; van Elburg, R.M.; Renes, I.B. Mildly pasteurized whey protein promotes gut tolerance in immature piglets compared with extensively heated whey protein. Nutrients 2020, 12, 3391. [Google Scholar] [CrossRef] [PubMed]
- Abbring, S.; Kusche, D.; Roos, T.C.; Diks, M.A.P.; Hols, G.; Garssen, J.; Baars, T.; van Esch, B.C.A.M. Milk processing increases the allergenicity of cow’s milk—Preclinical evidence supported by a human proof-of-concept provocation pilot. Clin. Exp. Allergy 2019, 49, 1013–1025. [Google Scholar] [CrossRef] [Green Version]
- Abbring, S.; Hols, G.; Garssen, J.; van Esch, B.C. Raw cow’s milk consumption and allergic diseases–the potential role of bioactive whey proteins. Eur. J. Pharmacol. 2019, 843, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Haller, N.; Kulozik, U. Continuous centrifugal separation of selectively precipitated α-lactalbumin. Int. Dairy J. 2020, 101, 104566. [Google Scholar] [CrossRef]
- Kella, N.K.D.; Kinsella, J.E. Enhanced thermodynamic stability of β-lactoglobulin at low pH. A possible mechanism. Biochem. J. 1988, 255, 113–118. [Google Scholar] [CrossRef]
- Kronman, M.J.; Andreotti, R.; Vitols, R. Inter-and intramolecular interactions of α-lactalbumin. II. Aggregation reactions at acid pH. Biochemistry 1964, 3, 1152–1160. [Google Scholar] [CrossRef]
- Bernal, V.; Jelen, P. Effect of calcium binding on thermal denaturation of bovine α-lactalbumin. J. Dairy Sci. 1984, 67, 2452–2454. [Google Scholar] [CrossRef]
- Alomirah, H.F.; Alli, I. Separation and characterization of β-lactoglobulin and α-lactalbumin from whey and whey protein preparations. Int. Dairy J. 2004, 14, 411–419. [Google Scholar] [CrossRef]
- Lucena, M.E.; Alvarez, S.; Menéndez, C.; Riera, F.A.; Alvarez, R. α-Lactalbumin precipitation from commercial whey protein concentrates. Sep. Purif. Technol. 2007, 52, 446–453. [Google Scholar]
- Bramaud, C.; Aimar, P.; Daufin, G. Whey protein fractionation: Isoelectric precipitation of α-lactalbumin under gentle heat treatment. Biotechnol. Bioeng. 1997, 56, 391–397. [Google Scholar] [CrossRef]
- Bonnaillie, L.M.; Tomasula, P.M. Kinetics, aggregation behavior and optimization of the fractionation of whey protein isolate with hydrochloric acid. Food Bioprocess Technol. 2012, 90, 737–747. [Google Scholar] [CrossRef]
- Pedersen, J.B.; Fojan, P.; Sorensen, J.; Petersen, S.B. Towards control of aggregational behaviour of α-lactalbumin at acidic pH. J. Fluoresc. 2006, 16, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Gésan-Guiziou, G.; Daufin, G.; Timmer, M.; Allersma, D.; van der Horst, C. Process steps for the preparation of purified fractions of α-lactalbumin and β-lactoglobulin from whey protein concentrates. J. Dairy Res. 1999, 66, 225–236. [Google Scholar] [CrossRef]
- Lucena, M.E.; Alvarez, S.; Menéndez, C.; Riera, F.A.; Alvarez, R. Beta-lactoglobulin removal from whey protein concentrates. Sep. Purif. Technol. 2006, 52, 310–316. [Google Scholar] [CrossRef]
- Fernández, A.; Menéndez, V.; Riera, F.A. α-Lactalbumin solubilisation from a precipitated whey protein concentrates fraction: pH and calcium concentration effects. Int. J. Food Sci. Technol. 2012, 47, 467–474. [Google Scholar] [CrossRef]
- Kronman, M.J.; Brew, K.; Sinha, S.K. Characteristics of the Binding of Ca2+ and Other Divalent Metal Ions to Bovine a-Lactalbumin. J. Biol. Chem. 1981, 256, 8582–8587. [Google Scholar] [CrossRef]
- Bramaud, C.; Aimar, P.; Daufin, G. Optimisation of a whey protein fractionation process based on the selective precipitation of α-lactalbumin. Le Lait 1997, 77, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.; Yu, S. Fourier transform infrared spectroscopic analysis of protein secondary structures. Acta Biochim. Biophys. Sin. 2007, 39, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Loughlin, I.B.; Kelly, P.M.; Murray, B.A.; FitzGerald, R.J.; Brodkorb, A. Concentrated whey protein ingredients: A Fourier transformed infrared spectroscopy investigation of thermally induced denaturation. Int. J. Dairy Technol. 2015, 68, 349–356. [Google Scholar] [CrossRef]
- Haller, N.; Greßlinger, A.S.; Kulozik, U. Separation of aggregated β-lactoglobulin with optimised yield in a decanter centrifuge. Int. Dairy J. 2021, 114, 104918. [Google Scholar] [CrossRef]
- Carbonaro, M.; Nucara, A. Secondary structure of food proteins by Fourier transform spectroscopy in the mid-infrared region. Amino Acids 2010, 38, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Lam, R.S.; Nickerson, M.T. The effect of pH and temperature pre-treatments on the structure, surface characteristics and emulsifying properties of alpha-lactalbumin. Food Chem. 2015, 173, 163–170. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, J.; Zhang, W.; Liu, C.; Jauregi, P.; Huang, M. Modification of heat-induced whey protein gels by basic amino acids. Food Hydrocoll. 2020, 100, 105397. [Google Scholar] [CrossRef]
- Zhong, H.; Gilmanshin, R.; Callender, R. An FTIR study of the complex melting behavior of α-lactalbumin. J. Phys. Chem. B 1999, 103, 3947–3953. [Google Scholar] [CrossRef]
- Boye, J.I.; Alli, I. Thermal denaturation of mixtures of α-lactalbumin and β-lactoglobulin: A differential scanning calorimetric study. Food Res. Int. 2000, 33, 673–682. [Google Scholar] [CrossRef]
- McGuffey, M.K.; Epting, K.L.; Kelly, R.M.; Foegeding, E.A. Denaturation and aggregation of three α-lactalbumin preparations at neutral pH. J. Agric. Food Chem. 2005, 53, 3182–3190. [Google Scholar] [CrossRef]
- Hsu, D.J.; Leshchev, D.; Kosheleva, I.; Kohlstedt, K.L.; Chen, L.X. Unfolding bovine α-lactalbumin with T-jump: Characterizing disordered intermediates via time-resolved x-ray solution scattering and molecular dynamics simulations. J. Chem. Phys. 2021, 154, 105101. [Google Scholar] [CrossRef]
- Bushmarina, N.A.; Blanchet, C.E.; Vernier, G.; Forge, V. Cofactor effects on the protein folding reaction: Acceleration of α-lactalbumin refolding by metal ions. Protein Sci. 2006, 15, 659–671. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.Y.; Li, L. Pathway of oxidative folding of α-lactalbumin: A model for illustrating the diversity of disulfide folding pathways. Biochemistry 2002, 41, 8405–8413. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.M.; Leaver, J.; Banks, J.M. Influence of other whey proteins on the heat-induced aggregation of α-lactalbumin. Int. Dairy J. 1993, 3, 719–727. [Google Scholar] [CrossRef]
- Rose, G.D.; Geselowitz, A.R.; Lesser, G.J.; Lee, R.H.; Zehfus, M.H. Hydrophobicity of amino acid residues in globular proteins. Science 1985, 229, 834–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhooren, A.; Chedad, A.; Farkas, V.; Majer, Z.; Joniau, M.; van Dael, H.; Hanssens, I. Tryptophan to phenylalanine substitutions allow differentiation of short-and long-range conformational changes during denaturation of goat α-lactalbumin. Proteins 2005, 60, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Stănciuc, N.; Râpeanu, G.; Bahrim, G.; Aprodu, I. pH and heat-induced structural changes of bovine apo-α-lactalbumin. Food Chem. 2012, 131, 956–963. [Google Scholar] [CrossRef]
- Goers, J.; Permyakov, S.E.; Permyakov, E.A.; Uversky, V.N.; Fink, A.L. Conformational prerequisites for α-lactalbumin fibrillation. Biochemistry 2002, 41, 12546–12551. [Google Scholar] [CrossRef]
- Boye, J.I.; Alli, I.; Ismail, A.A. Use of differential scanning calorimetry and infrared spectroscopy in the study of thermal and structural stability of α-lactalbumin. J. Agric. Food Chem. 1997, 45, 1116–1125. [Google Scholar] [CrossRef]
- Troullier, A.; Reinstädler, D.; Dupont, Y.; Naumann, D.; Forge, V. Transient non-native secondary structures during the refolding of α-lactalbumin detected by infrared spectroscopy. Nat. Struct. Biol. 2000, 7, 78–86. [Google Scholar]
- Veprintsev, D.B.; Permyakov, S.E.; Permyakov, E.A.; Rogov, V.V.; Cawthern, K.M.; Berliner, L.J. Co-operative thermal transitions of bovine and human apo-α-lactalbumins: Evidence for a new intermediate state. Febs Lett. 1997, 412, 625–628. [Google Scholar] [CrossRef] [Green Version]
- Kuhlman, B.; Boice, J.A.; Wu, W.J.; Fairman, R.; Raleigh, D.P. Calcium binding peptides from α-lactalbumin: Implications for protein folding and stability. Biochemistry 1997, 36, 4607–4615. [Google Scholar] [CrossRef]
- Dobson, C.M.; Šali, A.; Karplus, M. Protein folding: A perspective from theory and experiment. Angew. Chem. Int. Ed. 1998, 37, 868–893. [Google Scholar] [CrossRef]
- Hosseini-Nia, T.; Ismail, A.A.; Kubow, S. Effect of High Hydrostatic Pressure on the Secondary Structures of BSA and Apo-and Holo-α-Lactalbumin Employing Fourier Transform Infrared Spectroscopy. J. Food Sci. 2002, 67, 1341–1347. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haller, N.; Maier, I.; Kulozik, U. Molecular Analytical Assessment of Thermally Precipitated α-Lactalbumin after Resolubilization. Foods 2021, 10, 2231. https://doi.org/10.3390/foods10092231
Haller N, Maier I, Kulozik U. Molecular Analytical Assessment of Thermally Precipitated α-Lactalbumin after Resolubilization. Foods. 2021; 10(9):2231. https://doi.org/10.3390/foods10092231
Chicago/Turabian StyleHaller, Nicole, Isabel Maier, and Ulrich Kulozik. 2021. "Molecular Analytical Assessment of Thermally Precipitated α-Lactalbumin after Resolubilization" Foods 10, no. 9: 2231. https://doi.org/10.3390/foods10092231