
Design of an In Vitro Model to Screen the Chemical Reactivity Induced by Polyphenols and Vitamins during Digestion: An Application to Processed Meat

 and
and
Abstract
:1. Introduction
2. Materials and Methods
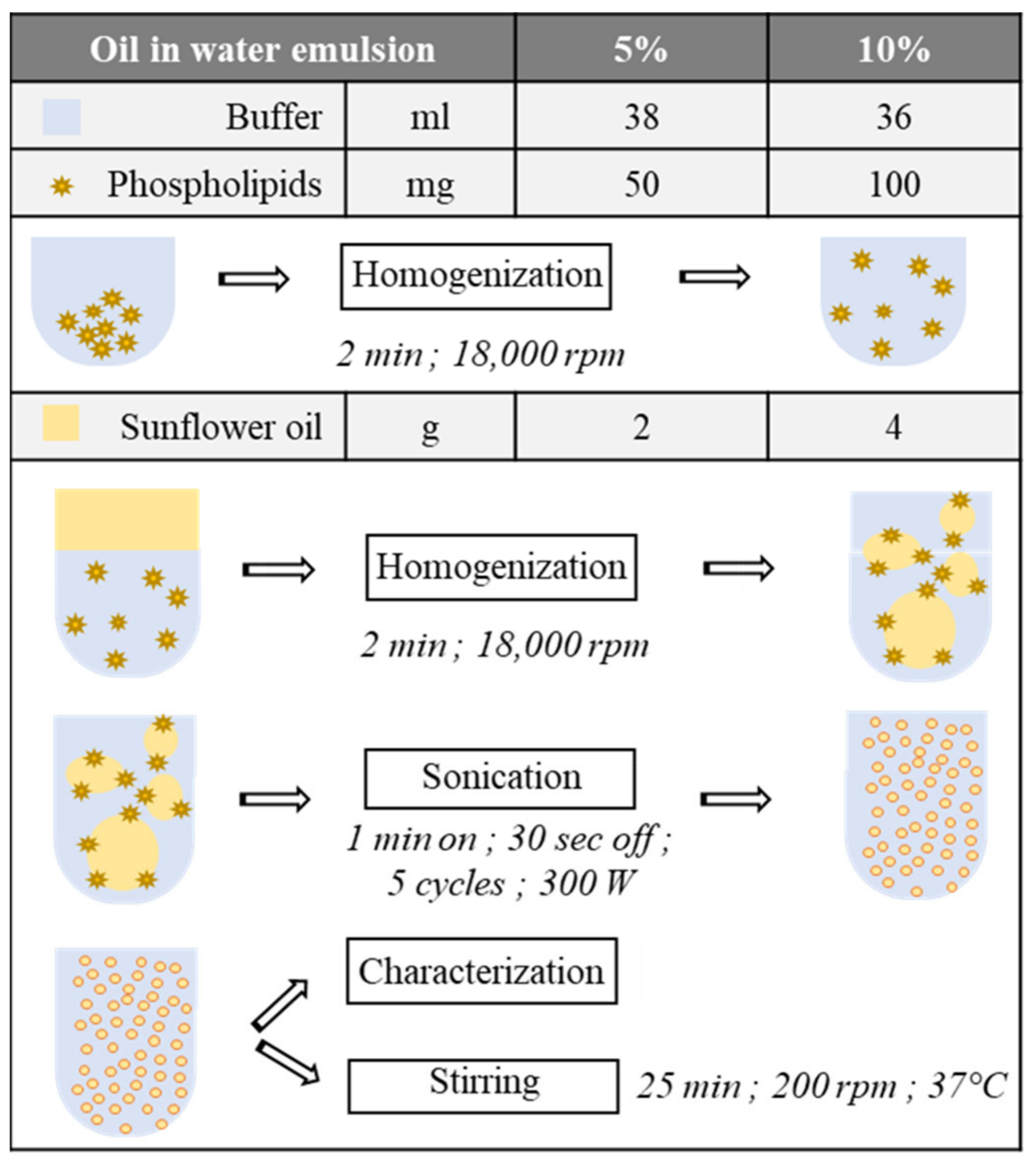
2.1. Oil in Water Emulsion
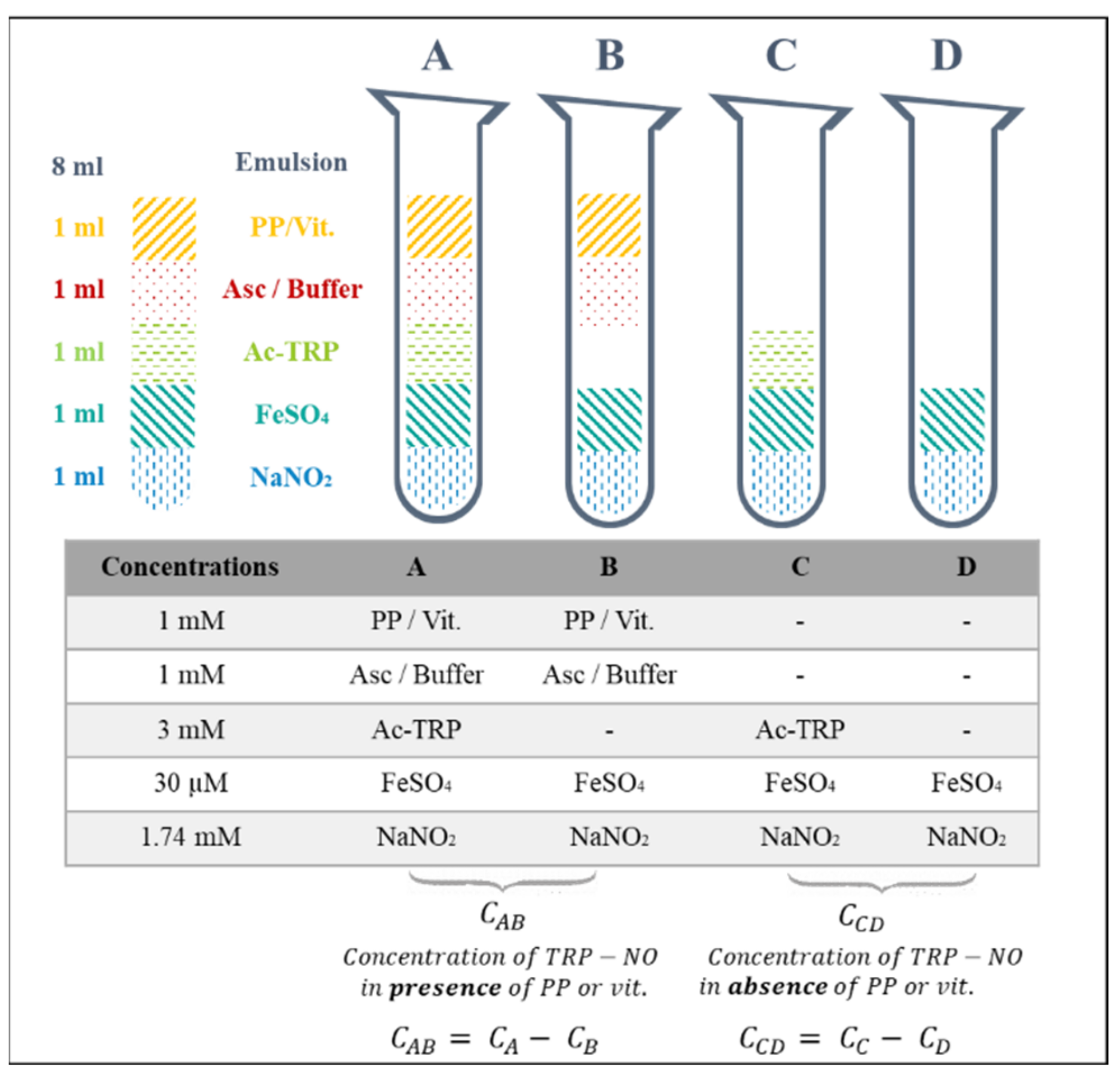
2.2. Screening Model
2.3. Biochemical Assays
2.4. Statistical Analysis
3. Results and Discussion
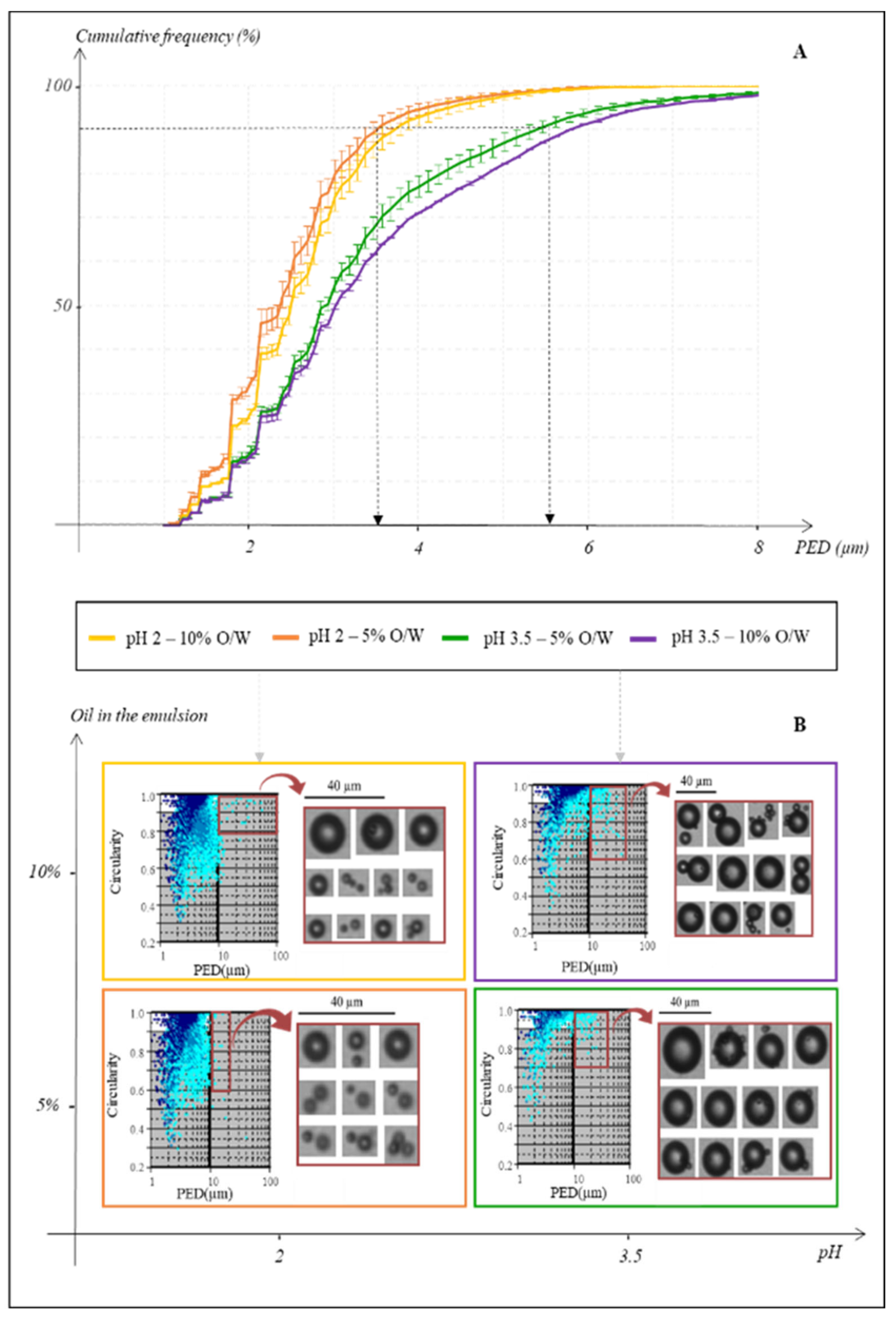
3.1. Optimization of the Heterogeneous Reactional Medium
3.2. Chemical Reactivity Modulation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- IARC. IARC Monographs evaluate consumption of red meat and processed meat. Press Release 2018, 240, 1–2. [Google Scholar]
- Gushgari, A.J.; Halden, R.U. Critical review of major sources of human exposure to N-nitrosamines. Chemosphere 2018, 210, 1124–1136. [Google Scholar] [CrossRef]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V. Mechanisms of Oxidative Processes in Meat and Toxicity Induced by Postprandial Degradation Products: A Review. Compr. Rev. Food Sci. Food Saf. 2017, 16, 96–123. [Google Scholar] [CrossRef]
- Nieva-Echevarria, B.; Goicoechea, E.; Guillen, M.D. Food lipid oxidation under gastrointestinal digestion conditions: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Esterbauer, H. Cytotoxicity and genotoxicity of lipid-oxidation products. Am. J. Clin. Nutr. 1993, 57, 779S–786S. [Google Scholar] [CrossRef] [Green Version]
- Negre-Salvayre, A.; Auge, N.; Ayala, V.; Basaga, H.; Boada, J.; Brenke, R.; Chapple, S.; Cohen, G.; Feher, J.; Grune, T. Pathological aspects of lipid peroxidation. Free Radic. Res. 2010, 44, 1125–1171. [Google Scholar] [CrossRef] [PubMed]
- Kumar, Y.; Yadav, D.N.; Ahmad, T.; Narsaiah, K. Recent Trends in the Use of Natural Antioxidants for Meat and Meat Products. Compr. Rev. Food Sci. Food Saf. 2015, 14, 796–812. [Google Scholar] [CrossRef] [Green Version]
- Dias, J.S. Nutritional Quality and Health Benefits of Vegetables: A Review. Food Nutr. Sci. 2012, 3, 1354–1374. [Google Scholar] [CrossRef] [Green Version]
- Bartsch, H.; Ohshima, H.; Pignatelli, B. Inhibitors of endogenous nitrosation mechanisms and implications in human cancer prevention. Mutat. Res. 1988, 202, 307–324. [Google Scholar] [CrossRef]
- Pierre, F.H.; Martin, O.C.; Santarelli, R.L.; Tache, S.; Naud, N.; Gueraud, F.; Audebert, M.; Dupuy, J.; Meunier, N.; Attaix, D. Calcium and alpha-tocopherol suppress cured-meat promotion of chemically induced colon carcinogenesis in rats and reduce associated biomarkers in human volunteers. Am. J. Clin. Nutr. 2013, 98, 1255–1562. [Google Scholar] [CrossRef] [Green Version]
- Kanner, J.; Selhub, J.; Shpaizer, A.; Rabkin, B.; Shacham, I.; Tirosh, O. Redox homeostasis in stomach medium by foods: The Postprandial Oxidative Stress Index (POSI) for balancing nutrition and human health. Redox. Biol. 2017, 12, 929–936. [Google Scholar] [CrossRef]
- Quedebeaux, B.; Eisa, H.M. Horticulture and Human Health: Contributions of Fruits and Vegetables. HortScience 1990, 25, 1473–1532. [Google Scholar]
- Craig, W.; Beck, L. Phytochemicals: Health Protective Effects. Can. J. Diet. Pract. Res. 1999, 60, 78–84. [Google Scholar] [PubMed]
- Wargovitch, M.J. Anticancer Properties of Fruits and Vegetables. HortScience 2000, 35, 573–575. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, M.S. Nutrition and cancer: A review of the evidence for an anti-cancer diet. Nutr. J. 2004, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Zhao, K.; Whiteman, M. The Gastrointestinal Tract: A Major Site of Antioxidant Action? Free Radic. Res. 2000, 33, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Kerem, Z.; Chetrit, D.; Shoseyov, O.; Regev-shoshani, G. Protection of Lipids from Oxidation by Epicatechin, trans-Resveratrol, and Gallic and Caffeic Acids in Intestinal Model Systems. J. Agric. Food Chem. 1993, 54, 10288–10293. [Google Scholar] [CrossRef]
- Barbieri, G.; Bergamaschi, M.; Saccani, G.; Caruso, G.; Santangelo, A.; Tulumello, R.; Vibhute, B.; Barbieri, G. Processed Meat and Polyphenols: Opportunities, Advantages, and Difficulties. J. AOAC Int. 2019, 102, 1401–1406. [Google Scholar] [CrossRef]
- Lee, S.Y.; Munerol, B.; Pollard, S.; Youdim, K.A.; Pannala, A.S.; Kuhnle, G.G.; Debnam, E.S.; Rice-Evans, C.; Spencer, J.P. The reaction of flavanols with nitrous acid protects against N-nitrosamine formation and leads to the formation of nitroso derivatives which inhibit cancer cell growth. Free Radic. Biol. Med. 2006, 40, 323–334. [Google Scholar] [CrossRef]
- Weir, T.L.; Trikha, S.R.J.; Thompson, H.J. Diet and cancer risk reduction: The role of diet-microbiota interactions and microbial metabolites. Semin. Cancer Biol. 2020, 70, 53–60. [Google Scholar] [CrossRef]
- Hrudey, S.E.; Bull, R.J.; Cotruvo, J.A.; Paoli, G.; Wilson, M. Drinking water as a proportion of total human exposure to volatile N-nitrosamines. Risk Anal. 2013, 33, 2179–2208. [Google Scholar] [CrossRef]
- Abraham, S.K.; Khandelwal, N. Ascorbic acid and dietary polyphenol combinations protect against genotoxic damage induced in mice by endogenous nitrosation. Mutat. Res. 2013, 757, 167–172. [Google Scholar] [CrossRef] [PubMed]
- d’Ischia, M.; Napolitano, A.; Manini, P.; Panzella, L. Secondary targets of nitrite-derived reactive nitrogen species: Nitrosation/nitration pathways, antioxidant defense mechanisms and toxicological implications. Chem. Res. Toxicol. 2011, 24, 2071–2092. [Google Scholar] [CrossRef] [PubMed]
- Hirota, S.; Takahama, U. Reactions of Apple Fruit Polyphenols with Nitrite under Conditions of the Gastric Lumen: Generation of Nitric Oxide and Formation of Nitroso Catechins. Food Sci. Technol. Res. 2014, 20, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Takahama, U.; Hirota, S. Possible Reactions of Dietary Phenolic Compounds with Salivary Nitrite and Thiocyanate in the Stomach. Antioxid. Basel 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peri, L.; Pietraforte, D.; Scorza, G.; Napolitano, A.; Fogliano, V.; Minetti, M. Apples increase nitric oxide production by human saliva at the acidic pH of the stomach: A new biological function for polyphenols with a catechol group? Free Radic. Biol. Med. 2005, 39, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Rao, G.S.; Osborn, J.C.; Adatia, M.R. Drug-Nitrite Interactions in Human Saliva: Effects of Food Constituents on Carcinogenic N-Nitrosamine Formation. J. Dent. Res. 1982, 6, 768–771. [Google Scholar] [CrossRef]
- Kanner, J.; Lapidot, T. The Stomach as a Bioreactor: Dietary Lipid Peroxidation in the Gastric Fluid and the Effects of Plant-Derived Antioxidants. Free Radic. Biol. Med. 2001, 31, 1388–1395. [Google Scholar] [CrossRef]
- Kenmogne-Domguia, H.B.; Meynier, A.; Boulanger, C.; Genot, C. Lipid Oxidation in Food Emulsions under Gastrointestinal-Simulated Conditions: The Key Role of Endogenous Tocopherols and Initiator. Food Dig. 2012, 3, 46–52. [Google Scholar] [CrossRef]
- Boléa, G.; Ginies, C.; Vallier, M.-J.; Dufour, C. Lipid protection by polyphenol-rich apple matrices is modulated by pH and pepsin in in vitro gastric digestion. Food Funct. 2019, 10, 3942–3954. [Google Scholar] [CrossRef]
- Lorrain, B.; Dangles, O.; Loonis, M.; Armand, M.; Dufour, C. Dietary iron-initiated lipid oxidation and its inhibition by polyphenols in gastric conditions. J. Agric. Food Chem. 2012, 60, 9074–9081. [Google Scholar] [CrossRef]
- Armand, M.; Borel, P.; Dubois, C.; Senft, M.; Peyrot, J.; Salducci, J.; Lafont, H.; Lairon, D. Characterization of emulsions and lipolysis of dietary lipids in the human stomach. Am. J. Physiol.-Gastrointest. Liver Physiol. 1994, 266, G372–G381. [Google Scholar] [CrossRef] [PubMed]
- Fraqueza, M.J.; Laranjo, M.; Elias, M.; Patarata, L. Microbiological hazards associated with salt and nitrite reduction in cured meat products: Control strategies based on antimicrobial effect of natural ingredients and protective microbiota. Curr. Opin. Food Sci. 2021, 38, 32–39. [Google Scholar] [CrossRef]
- Combet, E.; El Mesmari, A.; Preston, T.; Crozier, A.; McColl, K.E. Dietary phenolic acids and ascorbic acid: Influence on acid-catalyzed nitrosative chemistry in the presence and absence of lipids. Free Radic. Biol. Med. 2010, 48, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Promeyrat, A.; Gatellier, P.; Lebret, B.; Kajak-Siemaszko, K.; Aubry, L.; Santé-Lhoutellier, V. Evaluation of protein aggregation in cooked meat. Food Chem. 2010, 121, 412–417. [Google Scholar] [CrossRef]
- Strassner, J.E. Effect of pH on Interfacial Films and Stability of Crude Oil-Water Emulsions. J. Pet. Technol. 1968, 20, 303–312. [Google Scholar] [CrossRef]
- Raschke, D.; Knorr, D. Rapid monitoring of cell size, vitality and lipid droplet development in the oleaginous yeast Waltomyces lipofer. J. Microbiol. Methods 2009, 79, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Bonifacie, A.; Promeyrat, A.; Nassy, G.; Gatellier, P.; Sante-Lhoutellier, V.; Theron, L. Chemical reactivity of nitrite and ascorbate in a cured and cooked meat model implication in nitrosation, nitrosylation and oxidation. Food Chem. 2021, 348, 129073. [Google Scholar] [CrossRef]
- IFIP. Code des Usages de la Charcuterie, de la Salaison et des Conserves de Viande; IFIP: Paris, France, 2017. [Google Scholar]
- De La Pomelie, D.; Sante-Lhoutellier, V.; Gatellier, P. Mechanisms and kinetics of tryptophan N-nitrosation in a gastro-intestinal model. Food Chem. 2017, 218, 487–495. [Google Scholar] [CrossRef]
- Nakai, H.; Cassens, R.G.; Greaser, M.L.; Woolford, G. Significance of the reaction of nitrite with tryptophan. Inst. Food Technol. 1978, 43, 1857–1860. [Google Scholar] [CrossRef]
- Macheboeuf, D.; Morgavi, D.P.; Papon, Y.; Mousset, J.L.; Arturo-Schaan, M. Dose–response effects of essential oils on in vitro fermentation activity of the rumen microbial population. Anim. Feed. Sci. Technol. 2008, 145, 335–350. [Google Scholar] [CrossRef]
- Keceli, G.; Moore, C.D.; Toscano, J.P. Comparison of HNO reactivity with tryptophan and cysteine in small peptides. Bioorg. Med. Chem. Lett. 2014, 24, 3710–3713. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- McClements, D.J. Nanoemulsions versus microemulsions: Terminology, differences, and similarities. Soft Matter 2012, 8, 1719–1729. [Google Scholar] [CrossRef]
- Goodarzi, F.; Zendehboudi, S. A Comprehensive Review on Emulsions and Emulsion Stability in Chemical and Energy Industries. Can. J. Chem. Eng. 2018, 97, 281–309. [Google Scholar] [CrossRef] [Green Version]
- Combet, E.; Paterson, S.; Iijima, K.; Winter, J.; Mullen, W.; Crozier, A.; Preston, T.; McColl, K.E. Fat transforms ascorbic acid from inhibiting to promoting acid-catalysed N-nitrosation. Gut 2007, 56, 1678–1684. [Google Scholar] [CrossRef] [PubMed]
- Berton-Carabin, C.C.; Ropers, M.-H.; Genot, C. Lipid Oxidation in Oil-in-Water Emulsions: Involvement of the Interfacial Layer. Compr. Rev. Food Sci. Food Saf. 2014, 13, 945–977. [Google Scholar] [CrossRef]
- Cillard, J.; Cillard, P. Mécanismes de la peroxydation lipidique et des anti-oxydations. Ol. Corps Gras Lipides 2006, 13, 24–29. [Google Scholar] [CrossRef]
- Tannenbaum, S.R.; Wishnok, J.S.; Leaf, C.D. Inhibition of nitrosamine formation by ascorbic acid. Am. J. Clin. Nutr. 1991, 53, 247S–250S. [Google Scholar] [CrossRef] [PubMed]
- Hanuka Katz, I.; Eran Nagar, E.; Okun, Z.; Shpigelman, A. The Link between Polyphenol Structure, Antioxidant Capacity and Shelf-Life Stability in the Presence of Fructose and Ascorbic Acid. Molecules 2020, 25, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Lipid Oxidation (nmol MDA Equivalent/mg Lipids) | N-Nitrosation (%) | ||||
| Emulsion | Emulsion | Buffer | Emulsion | Emulsion | |
| - | Ascorbate | - | - | Ascorbate | |
| Control | 0.462 ± 0.052 | 0.367 ± 0.044 | 8.148 ± 1.601 | 5.825 ± 0.983 | 5.096 ± 1.044 |
| Ascorbate | 0.367 ± 0.044 NS | 0.592 ± 0.068 NS | 6.857 ± 2.653 NS | 5.096 ± 1.044 NS | 4.245 ± 1.045 NS |
| Trolox | 0.397 ± 0.051 NS | 0.454 ± 0.077 NS | 7.384 ± 1.283 NS | 4.354 ± 0.327 NS | 4.687 ± 2.096 NS |
| Chlorogenic Acid | 0.057 ± 0.013 *** | 0.058 ± 0.020 *** | 5.500 ± 2.864 NS | 6.630 ± 3.310 NS | 4.337 ± 1.564 NS |
| Rutin | 0.052 ± 0.008 *** | 0.035 ± 0.007 *** | 12.743 ± 1.940 *** | 9.112 ± 1.317 *** | 10.458 ± 0.893 *** |
| Naringenin | 0.227 ± 0.030 *** | 0.196 ± 0.032 *** | 7.485 ± 0.917 NS | 6.215 ± 2.176 NS | 6.026 ± 0.417 NS |
| Naringin | 0.287 ± 0.017 *** | 0.407 ± 0.013 NS | 8.351 ± 0.592 NS | 6.618 ± 1.398 NS | 5.053 ± 1.376 NS |
 |  | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keuleyan, E.; Bonifacie, A.; Gatellier, P.; Ferreira, C.; Blinet, S.; Promeyrat, A.; Nassy, G.; Santé-Lhoutellier, V.; Théron, L. Design of an In Vitro Model to Screen the Chemical Reactivity Induced by Polyphenols and Vitamins during Digestion: An Application to Processed Meat. Foods 2021, 10, 2230. https://doi.org/10.3390/foods10092230
Keuleyan E, Bonifacie A, Gatellier P, Ferreira C, Blinet S, Promeyrat A, Nassy G, Santé-Lhoutellier V, Théron L. Design of an In Vitro Model to Screen the Chemical Reactivity Induced by Polyphenols and Vitamins during Digestion: An Application to Processed Meat. Foods. 2021; 10(9):2230. https://doi.org/10.3390/foods10092230
Chicago/Turabian StyleKeuleyan, Eléna, Aline Bonifacie, Philippe Gatellier, Claude Ferreira, Sylvie Blinet, Aurélie Promeyrat, Gilles Nassy, Véronique Santé-Lhoutellier, and Laëtitia Théron. 2021. "Design of an In Vitro Model to Screen the Chemical Reactivity Induced by Polyphenols and Vitamins during Digestion: An Application to Processed Meat" Foods 10, no. 9: 2230. https://doi.org/10.3390/foods10092230