
New Insights into the Chemical Reactivity of Dry-Cured Fermented Sausages: Focus on Nitrosation, Nitrosylation and Oxidation

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
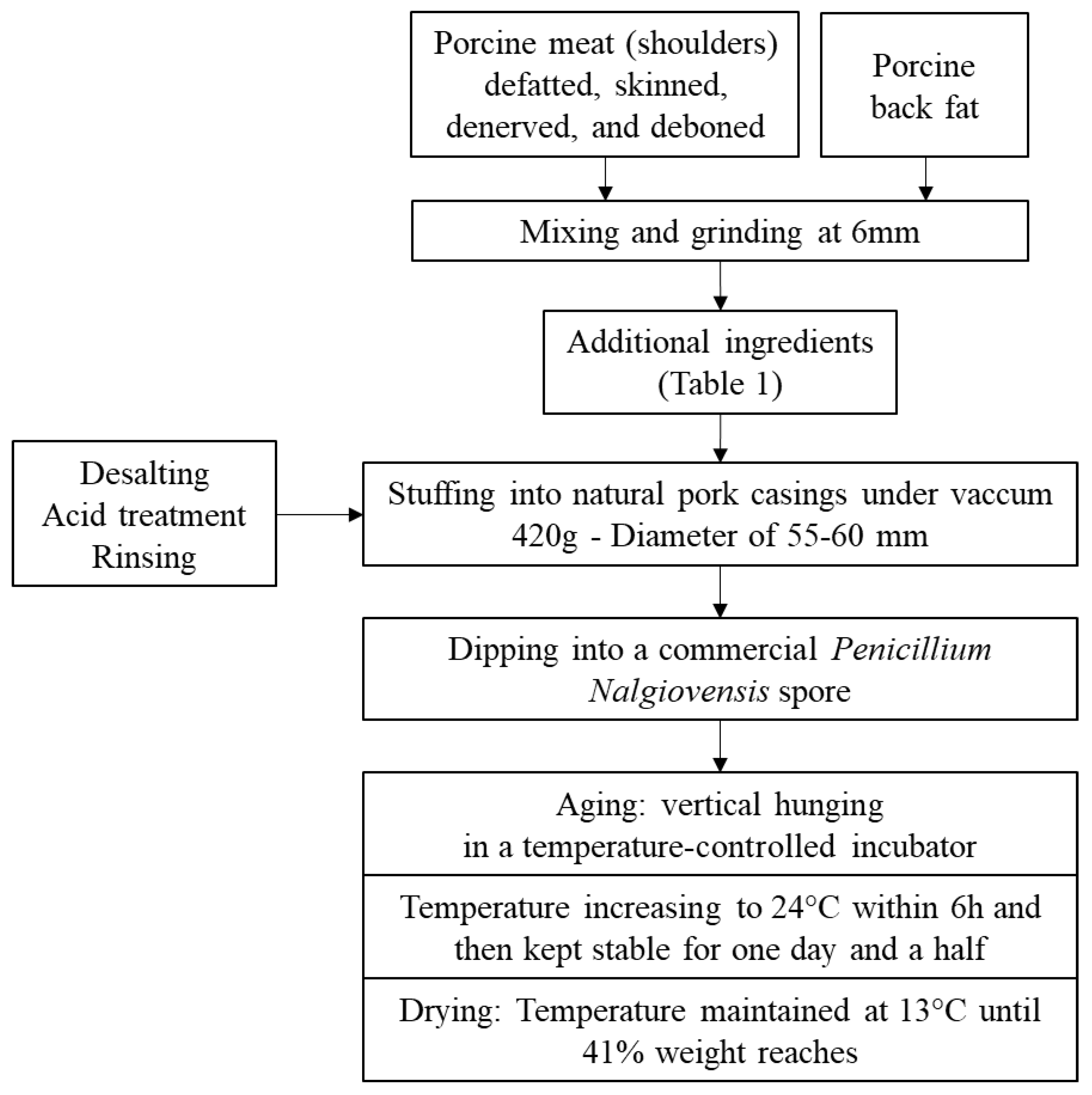
2.2. Preparation of Dry-Cured Fermented Meat Samples
2.3. Biochemical Characterization
2.3.1. Determination of Nitrite and Nitrate Content
2.3.2. Determination of Nitrosothiol Content
2.3.3. Determination of Non-Volatile Nitrosamine Content
2.3.4. Evaluation of Free Iron Content
2.3.5. Determination of Heme Iron Nitrosylation
2.3.6. Lipid Oxidation Measurement
2.3.7. Protein Oxidation Measurement
2.3.8. Proteolysis Index Measurement
2.4. Statistical Analysis
3. Results and Discussion
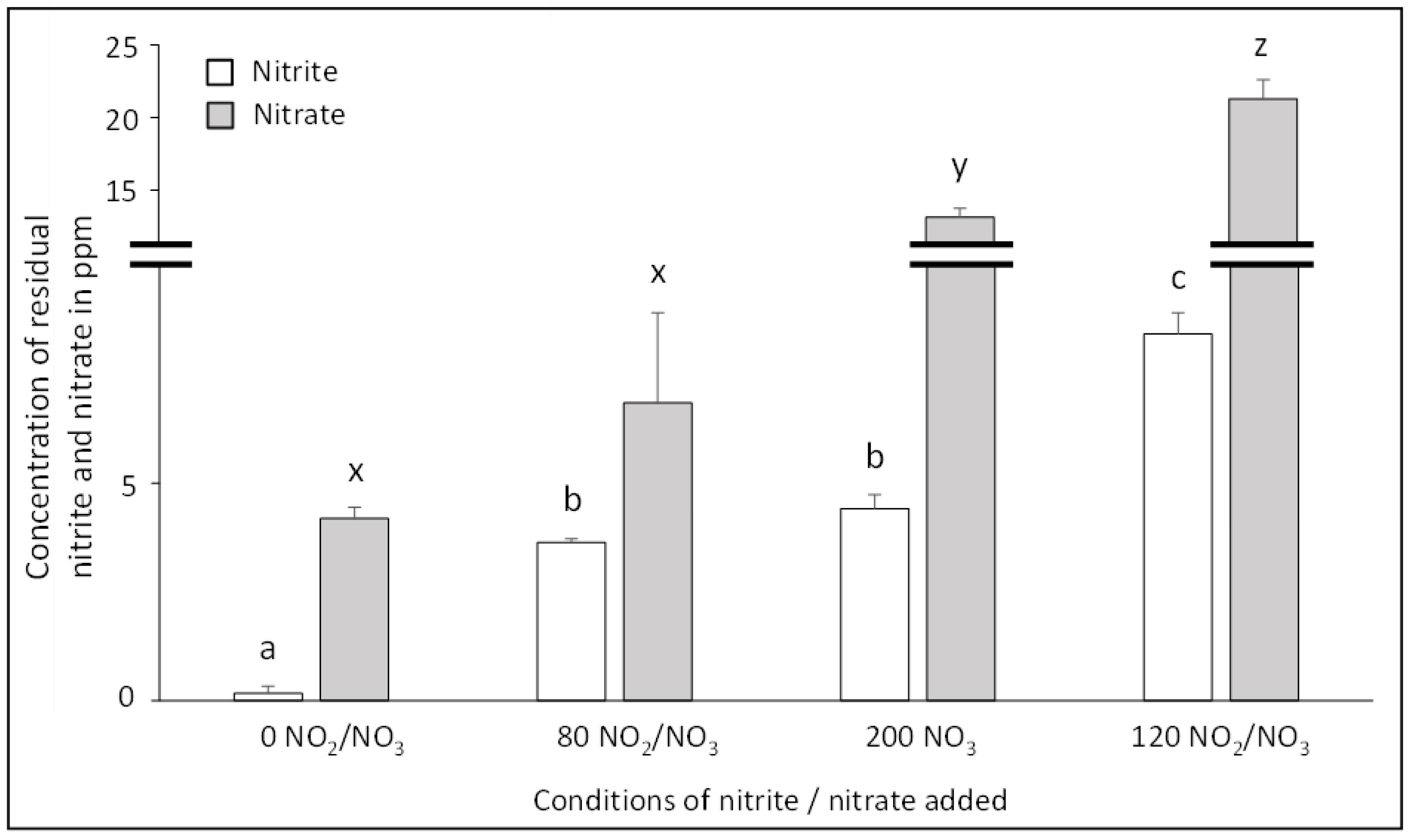
3.1. Impact of Sodium Nitrite/Nitrate Added to Residual Nitrite and Nitrate
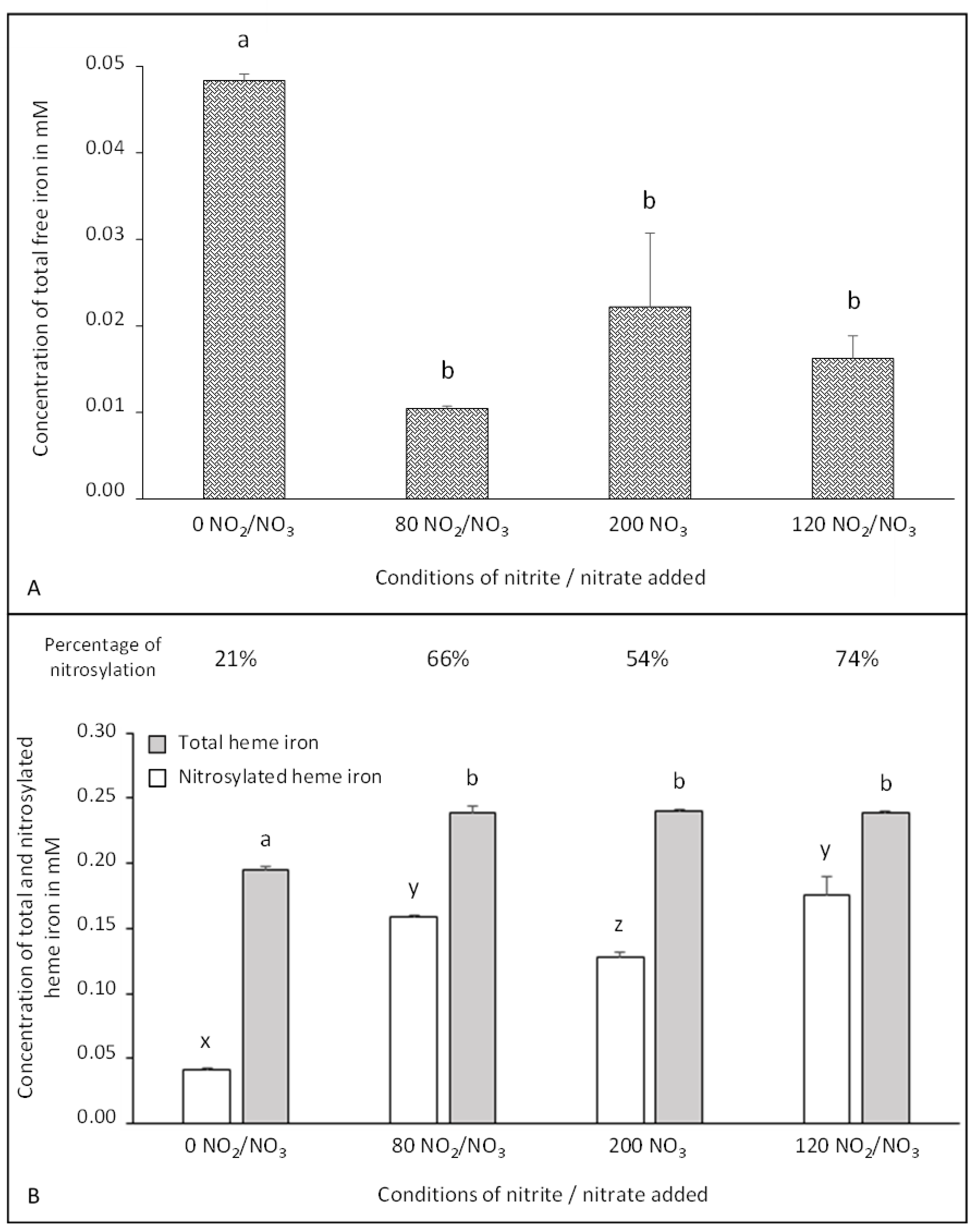
3.2. Impact of Sodium Nitrite/Nitrate Added to Free and Heme Iron
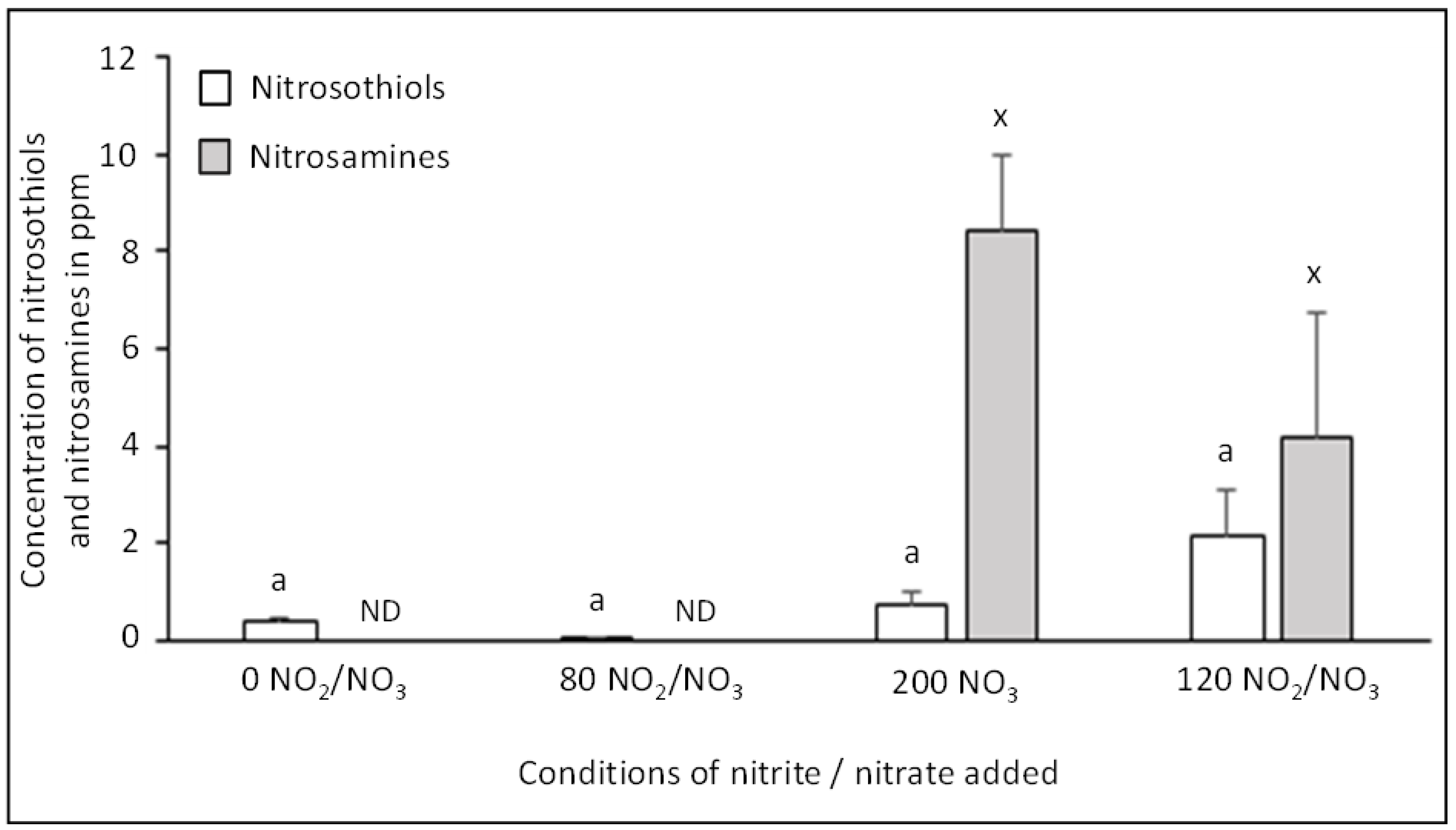
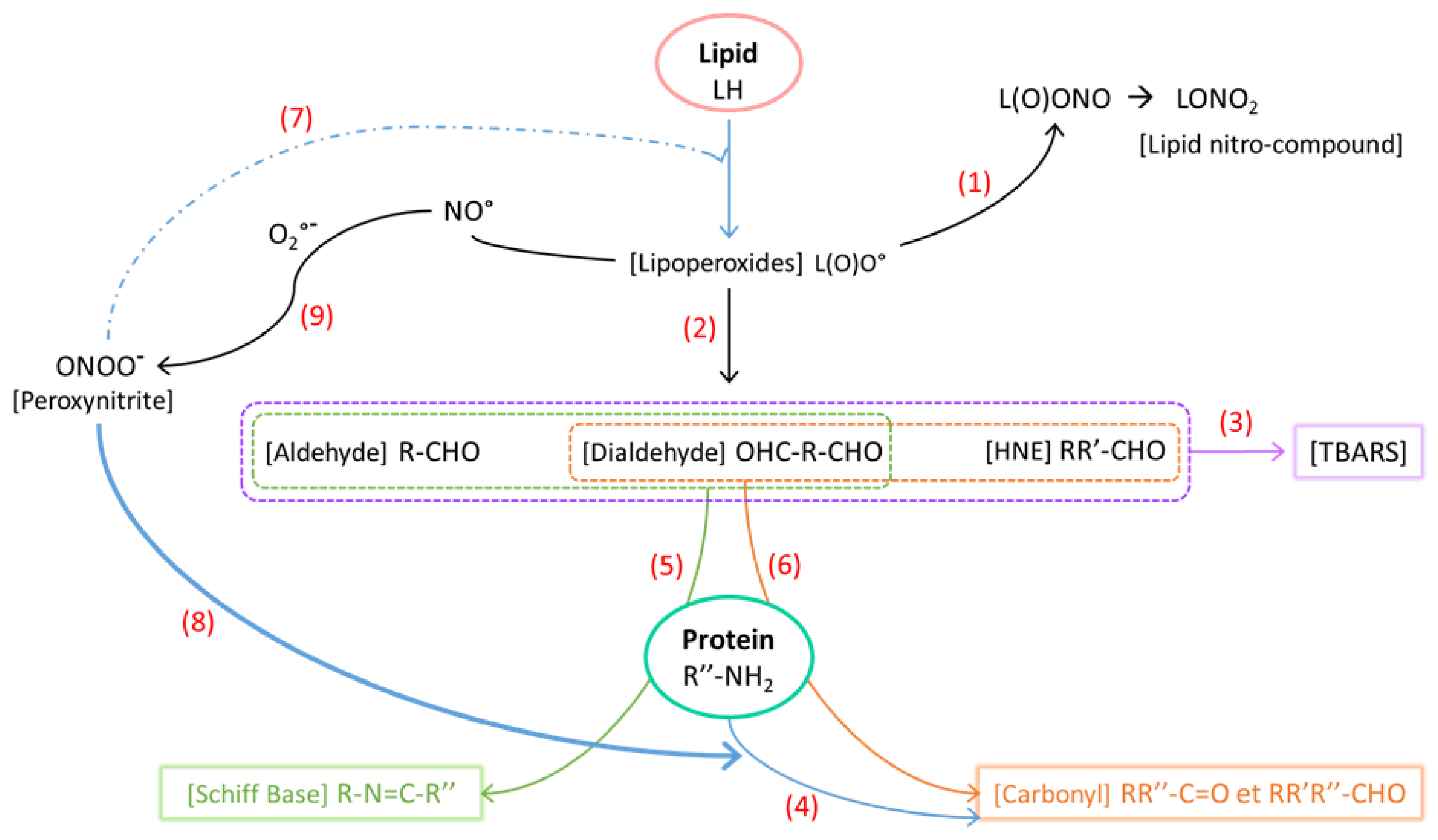
3.3. Impact of Sodium Nitrite/Nitrate Added on Nitrosation Reactions
3.4. Impact of Sodium Nitrite/Nitrate Added to Lipid and Protein Oxidation
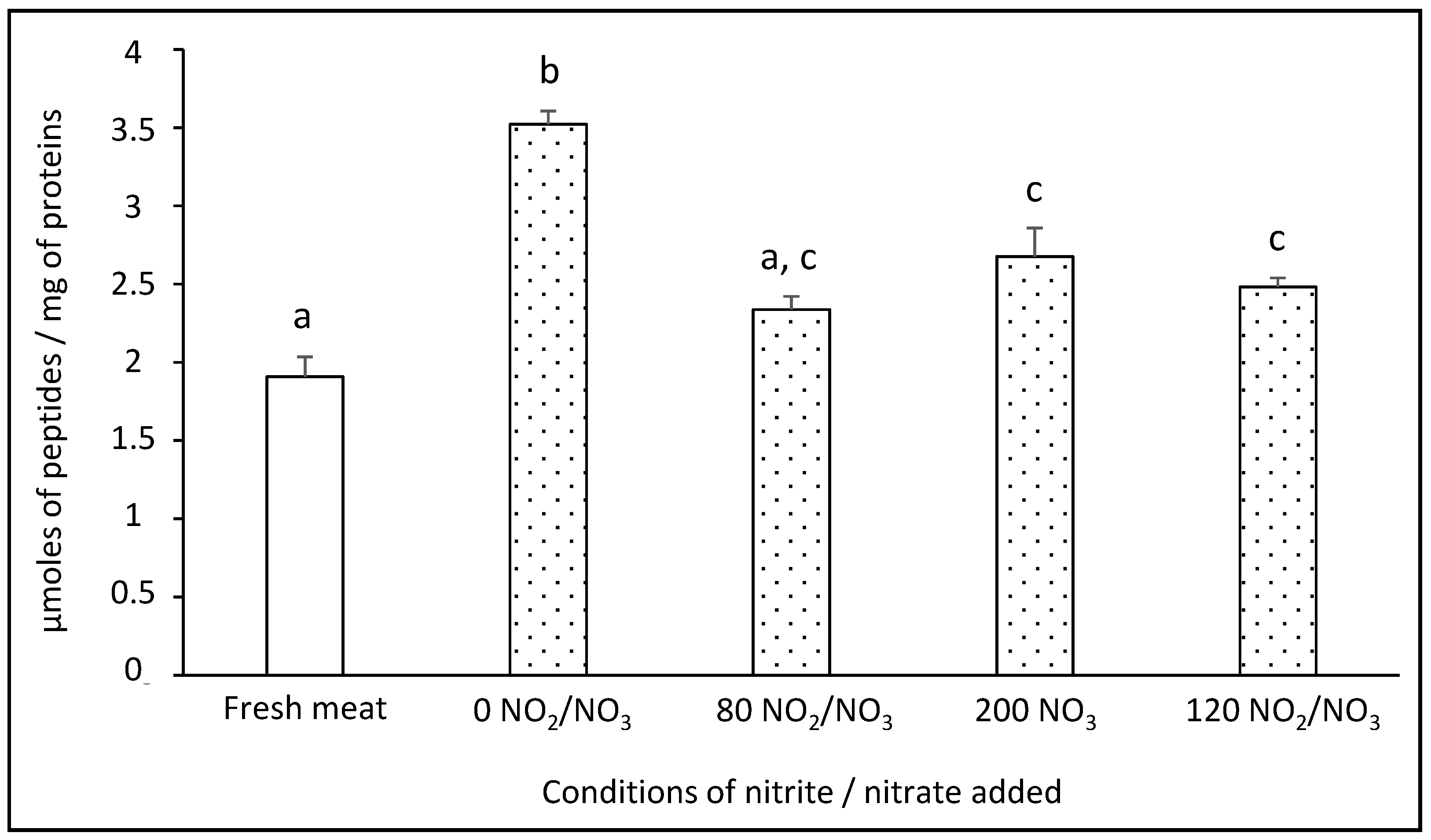
3.5. Impact of Added Sodium Nitrite/Nitrate on the Proteolysis Index
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hébel, P. La consommation de charcuteries en France. Cah. Nutr. Diététique 2019, 54, 5S16–5S22. [Google Scholar] [CrossRef]
- Hammes, W.P. Metabolism of nitrate in fermented meats: The characteristic feature of a specific group of fermented foods. Food Microbiol. 2012, 29, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Christieans, S.; Picgirard, L.; Parafita, E.; Lebert, A.; Gregori, T. Impact of reducing nitrate/nitrite levels on the behavior of Salmonella Typhimurium and Listeria monocytogenes in French dry fermented sausages. Meat Sci. 2018, 137, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, S.; Van Nieuwenhuysen, T.; Crèvecoeur, S.; Vanleyssem, R.; Thimister, J.; Denayer, S.; Jeuge, S.; Daube, G.; Clinquart, A.; Fremaux, B. Influence of reduced levels or suppression of sodium nitrite on the outgrowth and toxinogenesis of psychrotrophic Clostridium botulinum Group II type B in cooked ham. Int. J. Food Microbiol. 2020, 334, 108853. [Google Scholar] [CrossRef]
- Berardo, A.; De Maere, H.; Stavropoulou, D.; Rysman, T.; Leroy, F.; De Smet, S. Effect of sodium ascorbate and sodium nitrite on protein and lipid oxidation in dry fermented sausages. Meat Sci. 2016, 121, 359–364. [Google Scholar] [CrossRef]
- Sottero, B.; Leonarduzzi, G.; Testa, G.; Gargiulo, S.; Poli, G.; Biasi, F. Lipid Oxidation Derived Aldehydes and Oxysterols Between Health and Disease. Eur. J. Lipid Sci. Technol. 2019, 121, 1700047. [Google Scholar] [CrossRef] [Green Version]
- Honikel, K.-O. The use and control of nitrate and nitrite for the processing of meat products. Meat Sci. 2008, 78, 68–76. [Google Scholar] [CrossRef]
- De Mey, E.; De Maere, H.; Paelinck, H.; Fraeye, I. VolatileN-nitrosamines in meat products: Potential precursors, influence of processing, and mitigation strategies. Crit. Rev. Food Sci. Nutr. 2017, 57, 2909–2923. [Google Scholar] [CrossRef]
- Tricker, A.R.; Preussmann, R. Carcinogenic N-nitrosamines in the diet: Occurrence, formation, mechanisms and carcinogenic potential Mutat. Res. Genet. Toxicol. 1991, 259, 277–289. [Google Scholar] [CrossRef]
- Kuhnle, G.G.C.; Bingham, S.A. Dietary meat, endogenous nitrosation and colorectal cancer. Biochem. Soc. Trans. 2007, 35, 1355–1357. [Google Scholar] [CrossRef] [PubMed]
- Kostka, T.; Fohrer, J.; Guigas, C.; Briviba, K.; Seiwert, N.; Fahrer, J.; Steinberg, P.; Empl, M.T. Synthesis and in vitro characterization of the genotoxic, mutagenic and cell-transforming potential of nitrosylated heme. Arch. Toxicol. 2020, 94, 3911–3927. [Google Scholar] [CrossRef]
- Institut du Porc. Code des Usages de la Charcuterie, de la Salaison et des Conserves de Viande. Available online: https://www.ifip.asso.fr/fr/content/code-des-usages-de-la-charcuterie-de-la-salaison-et-des-conserves-de-viandes (accessed on 14 April 2017).
- Majou, D.; Christieans, S. Mechanisms of the bactericidal effects of nitrate and nitrite in cured meats. Meat Sci. 2018, 145, 273–284. [Google Scholar] [CrossRef]
- Perea-Sanz, L.; Montero, R.; Belloch, C.; Flores, M. Nitrate reduction in the fermentation process of salt reduced dry sausages: Impact on microbial and physicochemical parameters and aroma profile. Int. J. Food Microbiol. 2018, 282, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Safa, H.; Gatellier, P.; Lebert, A.; Picgirard, L.; Mirade, P.-S. Effect of Combined Salt and Animal Fat Reductions on Physicochemical and Biochemical Changes During the Manufacture of Dry-Fermented Sausages. Food Bioprocess Technol. 2015, 8, 2109–2122. [Google Scholar] [CrossRef]
- Toldra, F. Proteolysis and lipolysis in flavour development of dry-cured meat products. Meat Sci. 1998, 49, 101–110. [Google Scholar] [CrossRef]
- Suntsova, T.P.; Beda, N.V.; Nedospasov, A.A. Structural features of proteins responsible for resistanceof tryptophan residues to nitrosylation. IUBMB Life 2002, 54, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Bonifacie, A.; Aubry, L.; Gatellier, P.; Santé-Lhoutellier, V.; Théron, L. Determination of nitroso-compounds in food products. MethodsX 2021, 8, 101289. [Google Scholar] [CrossRef]
- Gaston, B. Nitric oxide and thiol groups. Biochim. Biophys. Acta 1999, 1411, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Stolze, K.; Dadak, A.; Liu, Y.; Nohl, H. Hydroxylamine and phenol-induced formation of methemoglobin and free radical intermediates in erythrocytes. Biochem. Pharmacol. 1996, 52, 1821–1829. [Google Scholar] [CrossRef]
- Bonifacie, A.; Promeyrat, A.; Nassy, G.; Gatellier, P.; Santé-Lhoutellier, V.; Théron, L. Chemical reactivity of nitrite and ascorbate in a cured and cooked meat model implication in nitrosation, nitrosylation and oxidation. Food Chem. 2021, 348, 129073. [Google Scholar] [CrossRef]
- Hornsey, H.C. The colour of cooked cured pork—Estimation of the Nitric oxide-Haem Pigments. J. Sci. Food Agric. 1956, 7, 534–540. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Mercier, Y.; Gatellier, P.; Viau, M.; Remignon, H.; Renerre, M. Effect of dietary fat and vitamin E on colour stability and on lipid and protein oxidation in Turkey meat during storage. Meat Sci. 1998, 48, 301–318. [Google Scholar] [CrossRef]
- Oliver, C.N.; Ahn, B.; Moermant, E.J.; Goldstein, S.; Stadtman, E.R. Age-related changes in oxidized proteins. J. Biol. Chem. 1987, 262, 5488–5491. [Google Scholar] [CrossRef]
- Morzel, M.; Gatellier, P.; Sayd, T.; Renerre, M.; Laville, E. Chemical oxidation decreases proteolytic susceptibility of skeletal muscle myofibrillar proteins. Meat Sci. 2006, 73, 536–543. [Google Scholar] [CrossRef]
- Harkouss, R.; Mirade, P.-S.; Gatellier, P. Development of a rapid, specific and efficient procedure for the determination of proteolytic activity in dry-cured ham: Definition of a new proteolysis index. Meat Sci. 2012, 92, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Iammarino, M.; Di Taranto, A. Nitrite and nitrate in fresh meats: A contribution to the estimation of admissible maximum limits to introduce in directive 95/2/EC. Int. J. Food Sci. Technol. 2012, 47, 1852–1858. [Google Scholar] [CrossRef]
- Laloo, A.E.E.; Wei, J.; Wang, D.; Narayanasamy, S.; Van Wonterghem, I.; Waite, D.; Steen, J.; Kaysen, A.; Heintz-Buschart, A.; Wang, Q.; et al. Mechanisms of Persistence of the Ammonia-Oxidizing BacteriaNitrosomonasto the Biocide Free Nitrous Acid. Environ. Sci. Technol. 2018, 52, 5386–5397. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.Y. Insights into the physiology of ammonia-oxidizing microorganisms. Curr. Opin. Chem. Biol. 2019, 49, 9–15. [Google Scholar] [CrossRef]
- Marco, A.; Navarro, J.L.; Flores, M. The influence of nitrite and nitrate on microbial, chemical and sensory parameters of slow dry fermented sausage. Meat Sci. 2006, 73, 660–673. [Google Scholar] [CrossRef]
- Sebranek, J.G.; Bacus, J.N. Cured meat products without direct addition of nitrate or nitrite: What are the issues? Meat Sci. 2007, 77, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Sallan, S.; Kaban, G.; Kaya, M. Nitrosamines in sucuk: Effects of black pepper, sodium ascorbate and cooking level. Food Chem. 2019, 288, 341–346. [Google Scholar] [CrossRef]
- Bechaux, J.; De La Pomélie, D.; Theron, L.; Santé-Lhoutellier, V.; Gatellier, P. Iron-catalysed chemistry in the gastrointestinal tract: Mechanisms, kinetics and consequences. A review. Food Chem. 2018, 268, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Juckett, M.; Zheng, Y.; Yuan, H.; Pastor, T.; Antholine, W.; Weber, M.; Vercelloti, G. Heme and the endothelium-effects of nitric oxide on catalytic iron and heme degradation by heme oxygenase. J. Biol. Chem. 1998, 273, 23388–23397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purchas, R.W.; Simcock, D.C.; Knight, T.W.; Wilkinson, B.H.P. Variation in the form of iron in beef and lamb meat and losses of iron during cooking and storage. Int. J. Food Sci. Technol. 2003, 38, 827–837. [Google Scholar] [CrossRef]
- Higuero, N.; Moreno, I.; Lavado, G.; Vidal-Aragón, M.C.; Cava, R. Reduction of nitrate and nitrite in Iberian dry cured loins and its effects during drying process. Meat Sci. 2020, 163, 108062. [Google Scholar] [CrossRef]
- Ibañez, C.; Quintanilla, L.; Cid, C.; Astiasarán, I.; Bello, J. Dry fermented sausages elaborated with Lactobacillus plantarum-Staphylococcus carnosus part I: Effect of partial replacement of NaCl with KCl on the stability and the nitrosation process. Meat Sci. 1996, 44, 227–234. [Google Scholar] [CrossRef]
- Herrmann, S.S.; Duedahl-Olesen, L.; Granby, K. Occurrence of volatile and non-volatile N-nitrosamines in processed meat products and the role of heat treatment. Food Control. 2015, 48, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Rubbo, H.; Batthyany, C.; Radi, R. Nitric Oxide: Oxygen Radical Interactions in Atherosclerosis. Biol. Res. 2000, 33, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, A.; Morcuende, D.; Estévez, M. Effect of Curing Agents on the Oxidative and Nitrosative Damage to Meat Proteins during Processing of Fermented Sausages. J. Food Sci. 2014, 79, C1331–C1342. [Google Scholar] [CrossRef] [PubMed]
- Estévez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Estévez, M. Oxidative damage to poultry: From farm to fork. Poult. Sci. 2015, 94, 1368–1378. [Google Scholar] [CrossRef]
- Callura, J.C. Activated Carbon Catalyzed Nitrosamine Formation via Amine Nitrosation. Ph.D. Thesis, Georgia Institute of Technology, Atlanta, GA, USA, 2014. [Google Scholar]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric Oxide and Peroxynitrite in Health and Disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [Green Version]
- Arnér, E.S.; Holmgren, A. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef]
- Schmitz, J.; Gilberg, E.; Löser, R.; Bajorath, J.; Bartz, U.; Gütschow, M. Cathepsin B: Active site mapping with peptidic substrates and inhibitors. Bioorganic Med. Chem. 2019, 27, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Saura, M.; Zaragoza, C.; McMillan, A.; Quick, R.A.; Hohenadl, C.; Lowenstein, J.M.; Lowenstein, C.J. An antiviral mechanism of nitric oxide: Inhibition of a viral protease. Immunity 1999, 10, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Aro Aro, J.M.; Nyam-Osor, P.; Tsuji, K.; Shimada, K.; Fukushima, M.; Sekikawa, M. The effect of starter cultures on proteolytic changes and amino acid content in fermented sausages. Food Chem. 2010, 119, 279–285. [Google Scholar] [CrossRef]
- Casaburi, A.; Di Monaco, R.; Cavella, S.; Toldrá, F.; Ercolini, D.; Villani, F. Proteolytic and lipolytic starter cultures and their effect on traditional fermented sausages ripening and sensory traits. Food Microbiol. 2008, 25, 335–347. [Google Scholar] [CrossRef]
- Gallego, M.; Mora, L.; Escudero, E.; Toldrá, F. Bioactive peptides and free amino acids profiles in different types of European dry-fermented sausages. Int. J. Food Microbiol. 2018, 276, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Candogan, K.; Wardlaw, F.B.; Acton, J.C. Effect of starter culture on proteolytic changes during processing of fermented beef sausages. Food Chem. 2009, 116, 731–737. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 NO2/NO3 | 80 NO2/NO3 | 200 NO3 | 120 NO2/NO3 | |
|---|---|---|---|---|
| Meat (shoulders) | 870 | |||
| Back fat | 130 | |||
| Ferment GY2 (Lb. sakei, S. carnosus, S. xylosus) | 0.15 | |||
| Ground grey pepper | 1.50 | |||
| Dextrose | 5.50 | |||
| Lactose | 6.00 | |||
| Natural salt (NaCl) | 26.00 | 12.67 | 26.00 | 6.00 |
| Nitrite salt (0.6% of NaNO2) | 0.00 | 13.33 | 0.00 | 20.00 |
| Potassium nitrate (KNO3) | 0.00 | 0.10 | 0.24 | 0.14 |
| 0 NO2/NO3 | 80 NO2/NO3 | 200 NO3 | 120 NO2/NO3 | |
|---|---|---|---|---|
| Dose NaNO2 (ppm) | 0 | 80 | 0 | 120 |
| Dose NaNO3 (ppm) | 0 | 80 | 200 | 120 |
| Concentration of total NO added (mM) | 0 | 3 | 3.2 | 4.5 |
| 0 NO2/NO3 | 80 NO2/NO3 | 200 NO3 | 120 NO2/NO3 | |
|---|---|---|---|---|
| TBARS µg MDA/g lipids | 12.67 a ± 0.39 | 7.34 b ± 0.41 | 8.39 b ± 0.89 | 6.05 b ± 0.91 |
| Carbonyl groups nmoles/mg proteins | 2.44 a ± 0.06 | 2.74 b ± 0.04 | 2.79 b ± 0.12 | 2.79 b ± 0.06 |
| Free thiol groups nmoles/mg proteins | 33.72 a ± 1.87 | 32.65 a ± 3.38 | 30.68 a ± 2.12 | 26.26 a ± 1.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonifacie, A.; Gatellier, P.; Promeyrat, A.; Nassy, G.; Picgirard, L.; Scislowski, V.; Santé-Lhoutellier, V.; Théron, L. New Insights into the Chemical Reactivity of Dry-Cured Fermented Sausages: Focus on Nitrosation, Nitrosylation and Oxidation. Foods 2021, 10, 852. https://doi.org/10.3390/foods10040852
Bonifacie A, Gatellier P, Promeyrat A, Nassy G, Picgirard L, Scislowski V, Santé-Lhoutellier V, Théron L. New Insights into the Chemical Reactivity of Dry-Cured Fermented Sausages: Focus on Nitrosation, Nitrosylation and Oxidation. Foods. 2021; 10(4):852. https://doi.org/10.3390/foods10040852
Chicago/Turabian StyleBonifacie, Aline, Philippe Gatellier, Aurélie Promeyrat, Gilles Nassy, Laurent Picgirard, Valérie Scislowski, Véronique Santé-Lhoutellier, and Laetitia Théron. 2021. "New Insights into the Chemical Reactivity of Dry-Cured Fermented Sausages: Focus on Nitrosation, Nitrosylation and Oxidation" Foods 10, no. 4: 852. https://doi.org/10.3390/foods10040852