
Ultrasonic Freezing Reduces Protein Oxidation and Myofibrillar Gel Quality Loss of Common Carp (Cyprinus carpio) during Long-Time Frozen Storage


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Frozen Fish
2.3. Extraction of Fish MP
2.4. Carbonyl Content
2.5. Dityrosine Content
2.6. Free Amine Content
2.7. Surface Hydrophobicity
2.8. Particle Size
2.9. Rheological Properties
2.10. The Water Holding Capacity of Fish MP Gel
2.11. The Gel Strength of Fish MP
2.12. Low Field Nuclear Magnetic (LF-NMR)
2.13. The Microstructure Fish MP Gel
2.14. Statistical Analysis
3. Results and Discussion
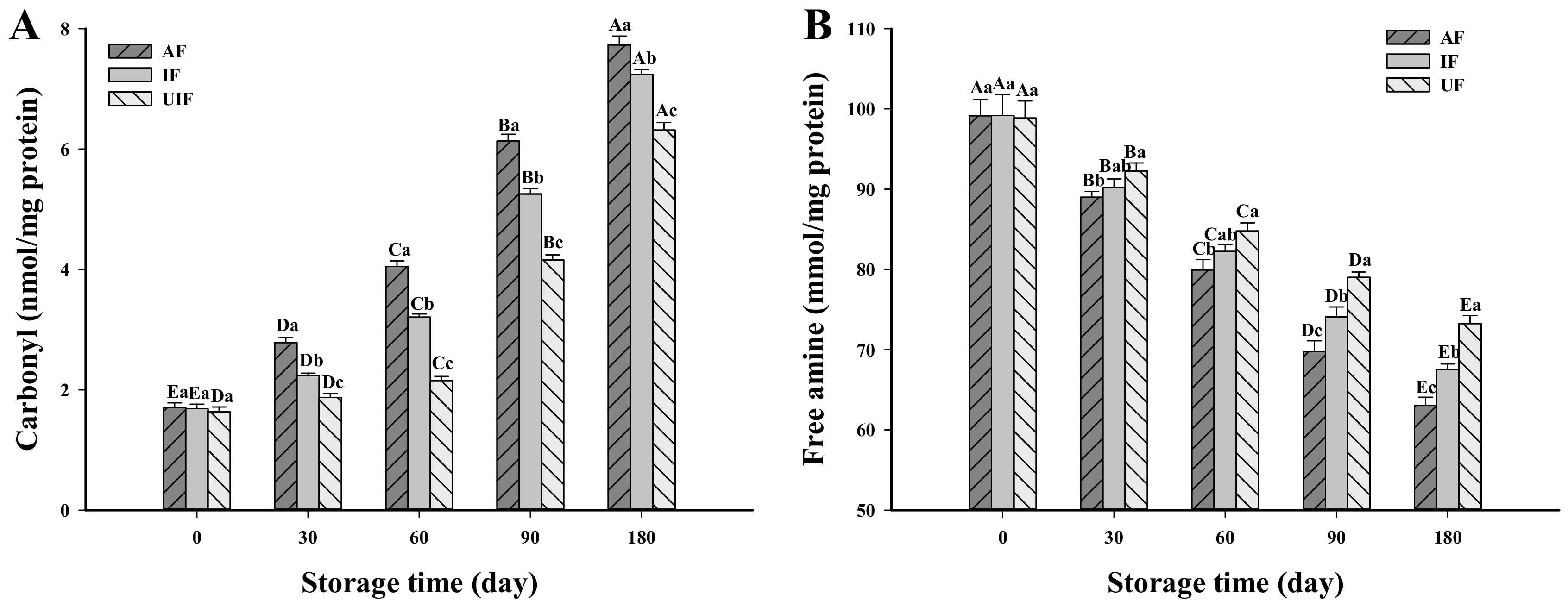
3.1. Change in Carbonyl Content
3.2. Change in Free Amine Content
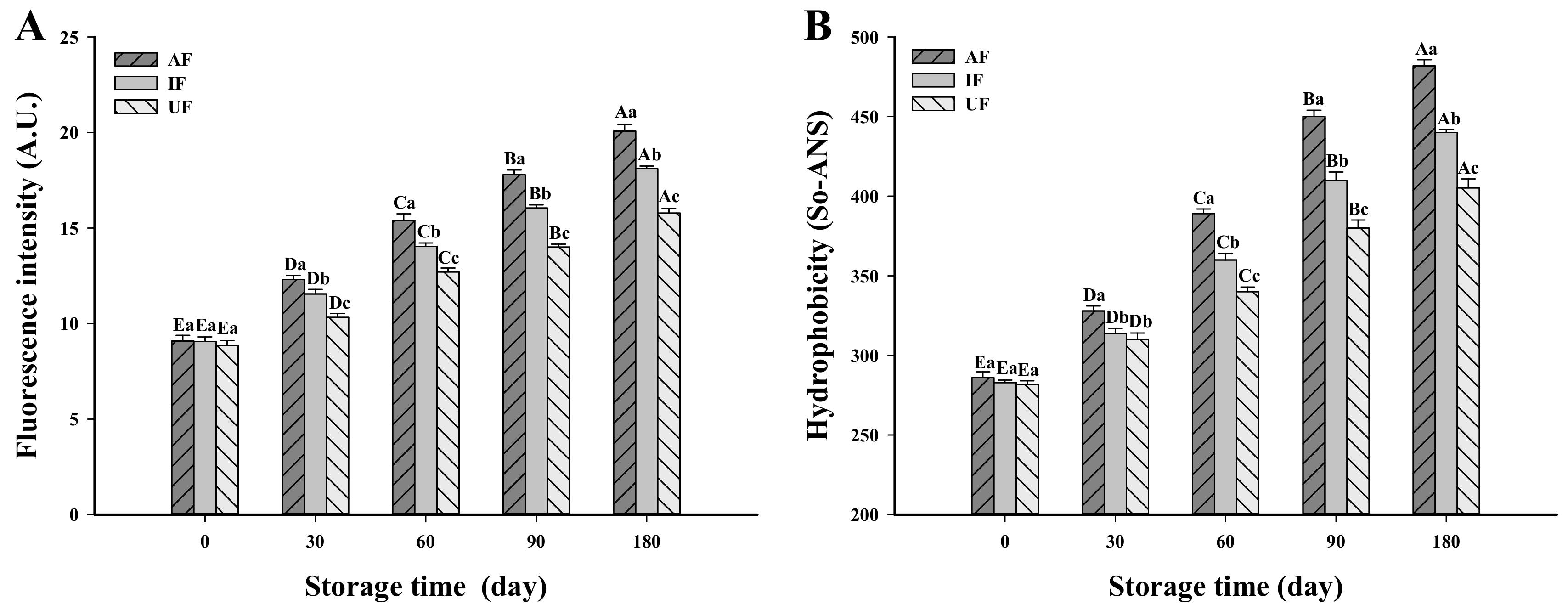
3.3. Change in Dityrosine Content
3.4. Change in Surface Hydrophobicity
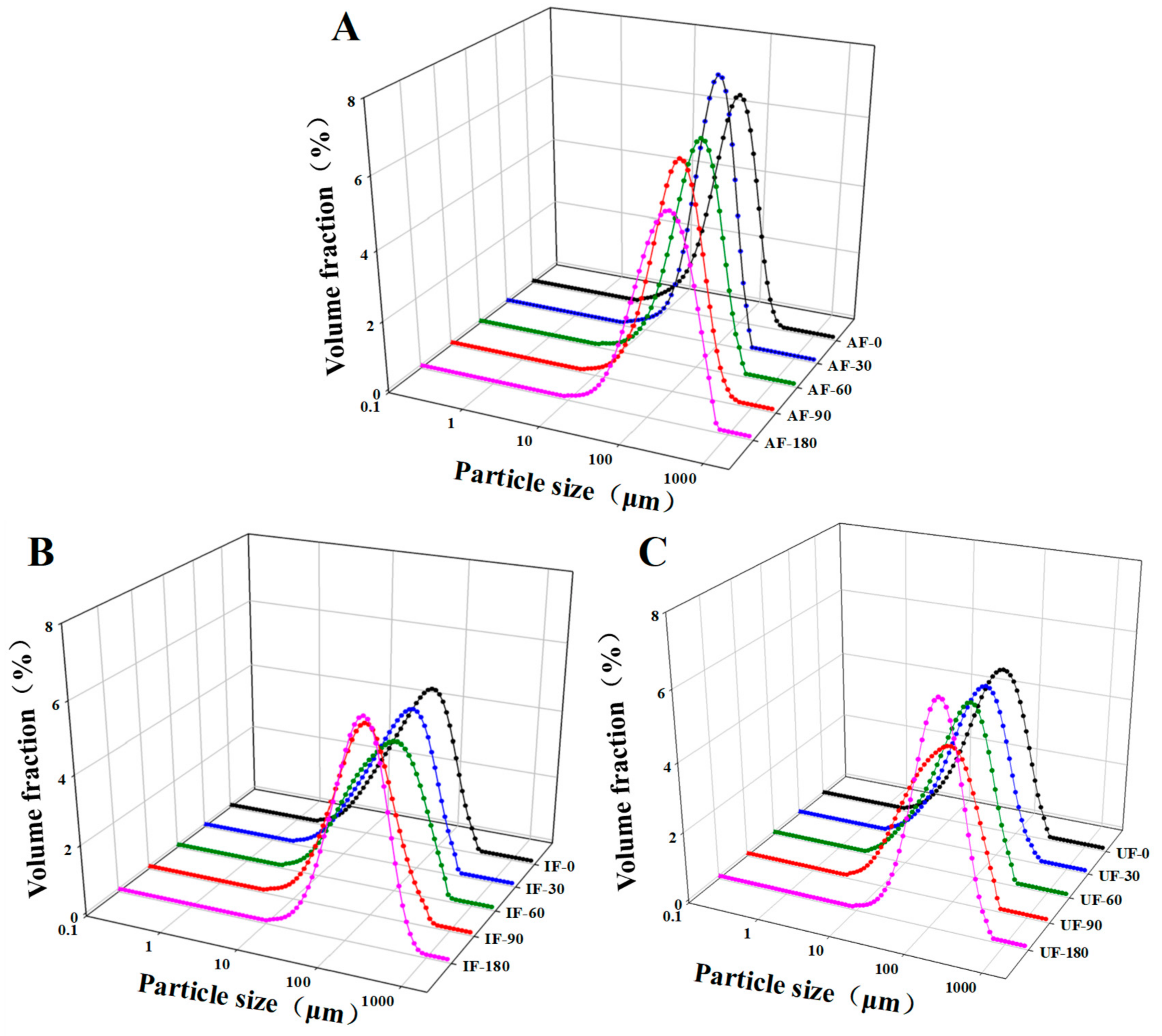
3.5. Particle Size Distribution
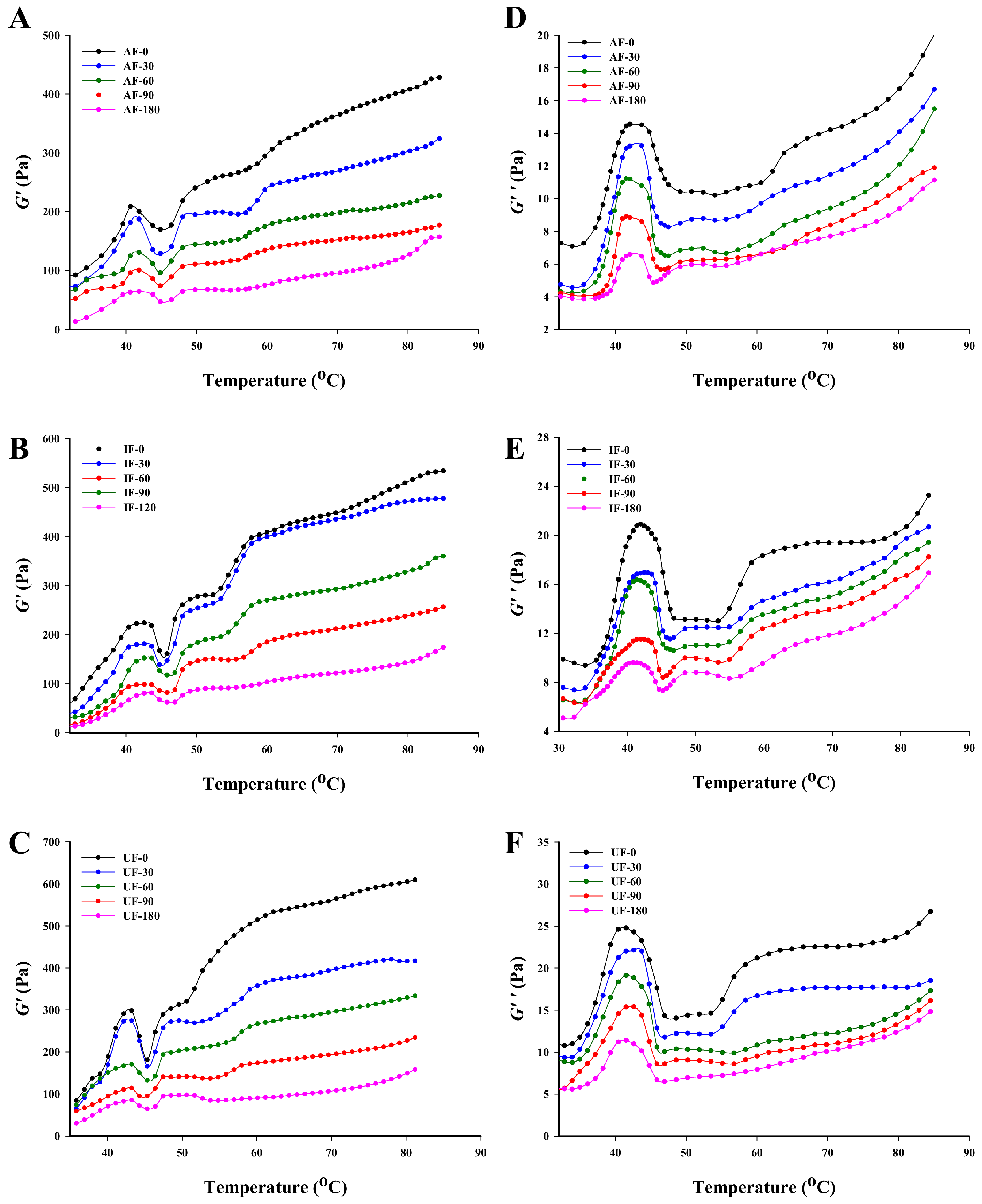
3.6. Rheological Properties
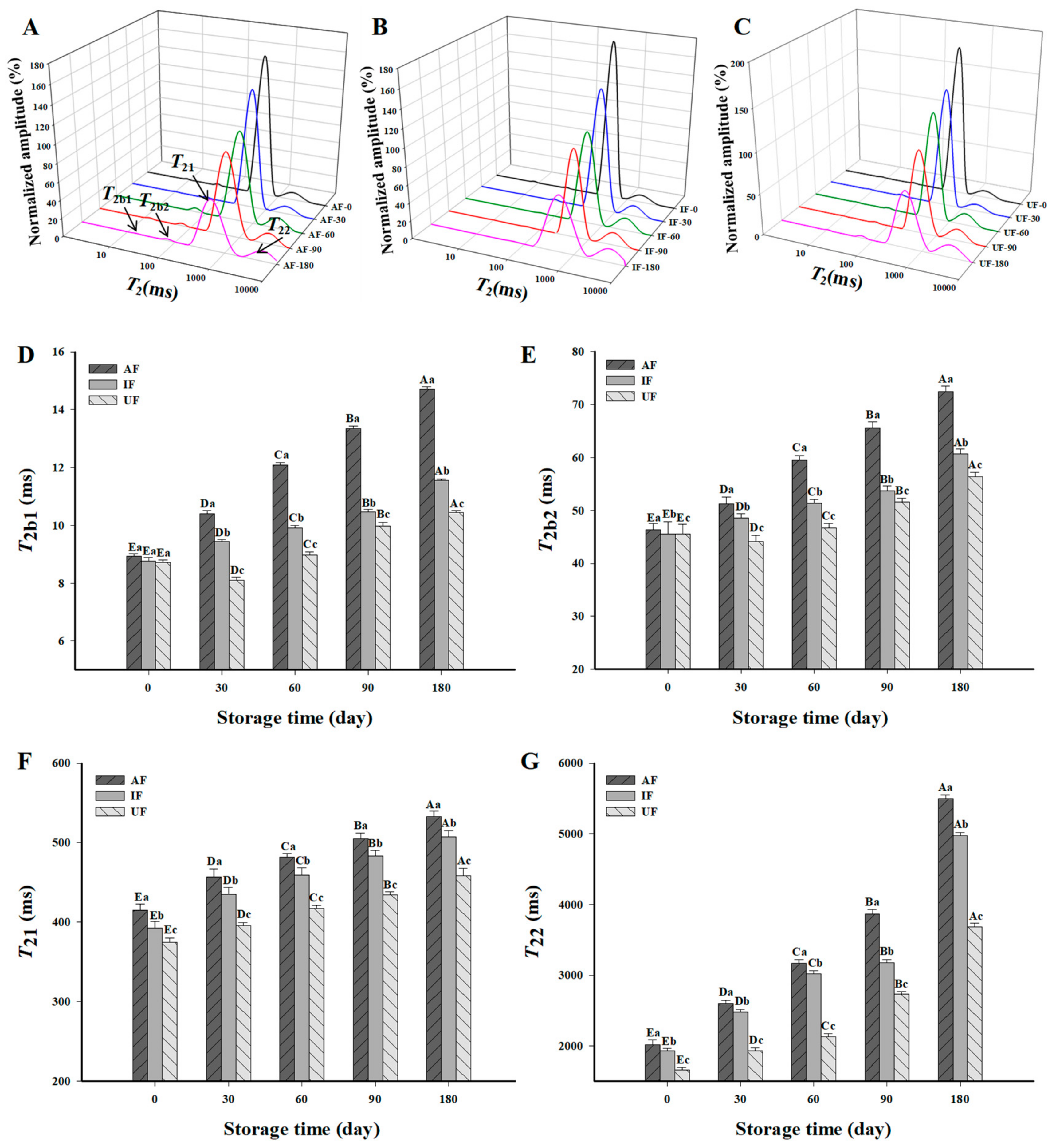
3.7. Water Distribution in Fish Protein Gel
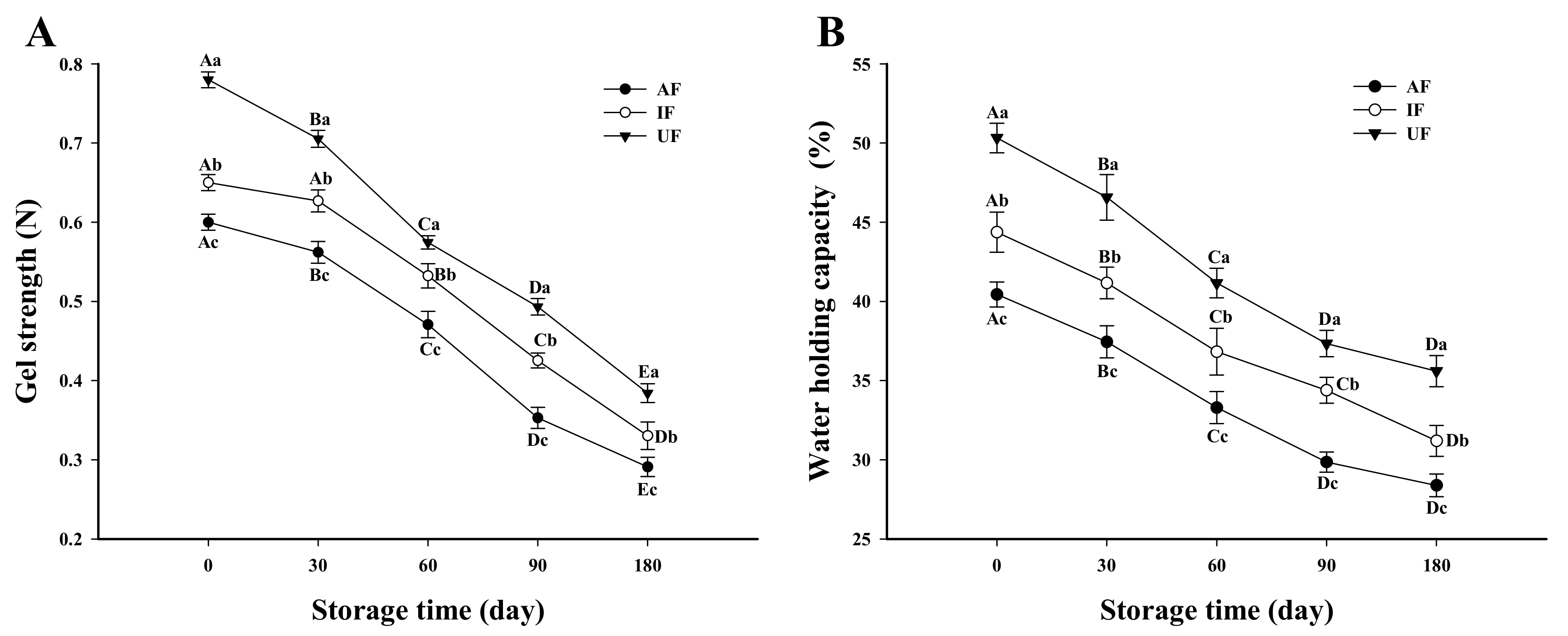
3.8. Gel Strength
3.9. WHC of Fish Protein Gel
3.10. Microstructure of Fish Protein Gel
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, D.; Zhao, H.; Muhammad, A.I.; Song, L.; Guo, M.; Liu, D. The comparison of ultrasound-assisted thawing, air thawing and water immersion thawing on the quality of slow/fast freezing bighead carp (Aristichthys nobilis) fillets. Food Chem. 2020, 320, 126614. [Google Scholar] [CrossRef]
- Li, D.; Qin, N.; Zhang, L.; Li, Q.; Prinyawiwatkul, W.; Luo, Y. Degradation of adenosine triphosphate, water loss and textural changes in frozen common carp (Cyprinus carpio) fillets during storage at different temperatures. Int. J. Refrig. 2019, 98, 294–301. [Google Scholar] [CrossRef]
- Tironi, V.; De Lamballerie, M.; Le-Bail, A. Quality changes during the frozen storage of sea bass (Dicentrarchus labrax) muscle after pressure shift freezing and pressure assisted thawing. Innov. Food Sci. Emerg. Technol. 2010, 11, 565–573. [Google Scholar] [CrossRef]
- Li, B.; Sun, D.-W. Novel methods for rapid freezing and thawing of foods—A review. J. Food Eng. 2002, 54, 175–182. [Google Scholar] [CrossRef]
- Tian, Y.; Chen, Z.; Zhu, Z.; Sun, D.-W. Effects of tissue pre-degassing followed by ultrasound-assisted freezing on freezing efficiency and quality attributes of radishes. Ultrason. Sonochem. 2020, 67, 105162. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Liu, X.; Wang, H.; Zhang, H. Effect of ultrasound-assisted freezing on the textural characteristics of dough and the structural characterization of wheat gluten. J. Food Sci. Technol. 2019, 56, 3380–3390. [Google Scholar] [CrossRef] [PubMed]
- Nowak, K.W.; Zielinska, M.; Waszkielis, K.M. The effect of ultrasound and freezing/thawing treatment on the physical properties of blueberries. Food Sci. Biotechnol. 2019, 28, 741–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Haili, N.; Mingcheng, Z.; Xia, X.; Kong, B. Influence of ultrasound-assisted immersion freezing on the freezing rate and quality of porcine longissimus muscles. Meat Sci. 2018, 136, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Zhang, M.; Chitrakar, B.; Bhandari, B. Application of power ultrasound in freezing and thawing Processes: Effect on process efficiency and product quality. Ultrason. Sonochem. 2020, 68, 105230. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Sun, F.; Xia, X.; Xu, H.; Kong, B. The comparison of ultrasound-assisted immersion freezing, air freezing and immersion freezing on the muscle quality and physicochemical properties of common carp (Cyprinus carpio) during freezing storage. Ultrason. Sonochem. 2019, 51, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Zhao, X.; Zhang, C.; Xia, X.; Sun, F.; Kong, B. Ultrasound-assisted immersion freezing accelerates the freezing process and improves the quality of common carp (Cyprinus carpio) at different power levels. LWT 2019, 108, 106–112. [Google Scholar] [CrossRef]
- Sun, Q.; Chen, Q.; Xia, X.; Kong, B.; Diao, X. Effects of ultrasound-assisted freezing at different power levels on the structure and thermal stability of common carp (Cyprinus carpio) proteins. Ultrason. Sonochem. 2019, 54, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Fang, C.-D.; Hao, G.-J.; Zhang, Y.-Y. Effect of kappa-carrageenan oligosaccharides on myofibrillar protein oxidation in peeled shrimp (Litopenaeus vannamei) during long-term frozen storage. Food Chem. 2018, 245, 254–261. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhou, F.; Zhang, L.; Wang, H.; Wang, X.-C. Effect of different extent of protein oxidation on the frozen storage stability of muscle protein in obscure pufferfish (Takifugu obscurus). LWT 2021, 137, 110416. [Google Scholar] [CrossRef]
- An, Y.; You, J.; Xiong, S.; Yin, T. Short-term frozen storage enhances cross-linking that was induced by transglutaminase in surimi gels from silver carp (Hypophthalmichthys molitrix). Food Chem. 2018, 257, 216–222. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, C.; Li, Q.; Xia, X.; Kong, B. Changes in functional properties of common carp (Cyprinus carpio) myofibrillar protein as affected by ultrasound-assisted freezing. J. Food Sci. 2020, 85, 2879–2888. [Google Scholar] [CrossRef] [PubMed]
- Utrera, M.; Armenteros, M.; Ventanas, S.; Solano, F.; Estévez, M. Pre-freezing raw hams affects quality traits in cooked hams: Potential influence of protein oxidation. Meat Sci. 2012, 92, 596–603. [Google Scholar] [CrossRef]
- Panepucci, R.A.; Siufi, J.L.; Silva, W.A.; Proto-Siquiera, R.; Neder, L.; Orellana, M.; Rocha, V.; Covas, D.T.; Zago, M.A. Comparison of Gene Expression of Umbilical Cord Vein and Bone Marrow-Derived Mesenchymal Stem Cells. STEM CELLS 2004, 22, 1263–1278. [Google Scholar] [CrossRef] [PubMed]
- Utrera, M.; Parra, V.; Estévez, M. Protein oxidation during frozen storage and subsequent processing of different beef muscles. Meat Sci. 2014, 96, 812–820. [Google Scholar] [CrossRef]
- Faustman, C.; Sun, Q.; Mancini, R.; Suman, S.P. Myoglobin and lipid oxidation interactions: Mechanistic bases and control. Meat Sci. 2010, 86, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.-C.; Zou, Y.-H.; Cheng, Y.-P.; Xing, L.-J.; Zhou, G.-H.; Zhang, W.-G. Effects of power ultrasound on oxidation and structure of beef proteins during curing processing. Ultrason. Sonochem. 2016, 33, 47–53. [Google Scholar] [CrossRef]
- Nikoo, M.; Benjakul, S.; Rahmanifarah, K. Hydrolysates from marine sources as cryoprotective substances in seafoods and seafood products. Trends Food Sci. Technol. 2016, 57, 40–51. [Google Scholar] [CrossRef]
- Estévez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Zhang, M.; Li, F.; Diao, X.; Kong, B.; Xia, X. Moisture migration, microstructure damage and protein structure changes in porcine longissimus muscle as influenced by multiple freeze-thaw cycles. Meat Sci. 2017, 133, 10–18. [Google Scholar] [CrossRef]
- Cao, Y.; Xiong, Y.L. Chlorogenic acid-mediated gel formation of oxidatively stressed myofibrillar protein. Food Chem. 2015, 180, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.-J.; Lee, D.-I.; Lee, C.-H.; Ahn, I.-S. A dityrosine-based substrate for a protease assay: Application for the selective assessment of papain and chymopapain activity. Anal. Chim. Acta 2012, 723, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Gillies, D.; Wagner, G.; Howell, N.K. ESR and NMR spectroscopy studies on protein oxidation and formation of dityrosine in emulsions containing oxidised methyl linoleate. Food Chem. Toxicol. 2006, 44, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Colombo, G.; Clerici, M.; Giustarini, D.; Portinaro, N.; Badalamenti, S.; Rossi, R.; Milzani, A.; Dalle-Donne, I. A central role for intermolecular dityrosine cross-linking of fibrinogen in high molecular weight advanced oxidation protein product (AOPP) formation. Biochim. et Biophys. Acta (BBA) Gen. Subj. 2015, 1850, 1–12. [Google Scholar] [CrossRef]
- Ragnarsson, K.; Regenstein, J.M. Changes in Electrophoretic Patterns of Gadoid and Non-gadoid Fish Muscle during Frozen Storage. J. Food Sci. 1989, 54, 819–823. [Google Scholar] [CrossRef]
- Ang, J.F.; Hultin, H.O. Denaturation of Cod Myosin during Freezing after Modification with Formaldehyde. J. Food Sci. 1989, 54, 814–818. [Google Scholar] [CrossRef]
- Careche, M.; Li-Chan, E. Structural Changes in Cod Myosin after Modification with Formaldehyde or Frozen Storage. J. Food Sci. 1997, 62, 717–723. [Google Scholar] [CrossRef] [Green Version]
- Wagner, J.R.; Añón, M.C. Effect of frozen storage on protein denaturation in bovine muscle. Int. J. Food Sci. Technol. 1986, 21, 547–558. [Google Scholar] [CrossRef]
- Badii, F.; Howell, N.K. A comparison of biochemical changes in cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) fillets during frozen storage. J. Sci. Food Agric. 2001, 82, 87–97. [Google Scholar] [CrossRef]
- Resendiz-Vazquez, J.; Ulloa, J.; Urías-Silvas, J.; Bautista-Rosales, P.; Ramírez-Ramírez, J.; Rosas-Ulloa, P.; González-Torres, L. Effect of high-intensity ultrasound on the technofunctional properties and structure of jackfruit (Artocarpus heterophyllus) seed protein isolate. Ultrason. Sonochemistry 2017, 37, 436–444. [Google Scholar] [CrossRef]
- Zhao, J.; Dong, F.; Li, Y.; Kong, B.; Liu, Q. Effect of freeze–thaw cycles on the emulsion activity and structural characteristics of soy protein isolate. Process. Biochem. 2015, 50, 1607–1613. [Google Scholar] [CrossRef]
- Promeyrat, A.; Gatellier, P.; Lebret, B.; Kajak-Siemaszko, K.; Aubry, L.; Santé-Lhoutellier, V. Evaluation of protein aggregation in cooked meat. Food Chem. 2010, 121, 412–417. [Google Scholar] [CrossRef]
- Walayat, N.; Xiong, Z.; Xiong, H.; Moreno, H.M.; Li, Q.; Nawaz, A.; Zhang, Z.; Wang, P.; Niaz, N. The effectiveness of egg white protein and β-cyclodextrin during frozen storage: Functional, rheological and structural changes in the myofibrillar proteins of Culter alburnus. Food Hydrocoll. 2020, 105, 105842. [Google Scholar] [CrossRef]
- Zhao, Y.-Y.; Wang, P.; Zou, Y.-F.; Li, K.; Kang, Z.-L.; Xu, X.-L.; Zhou, G.-H. Effect of pre-emulsification of plant lipid treated by pulsed ultrasound on the functional properties of chicken breast myofibrillar protein composite gel. Food Res. Int. 2014, 58, 98–104. [Google Scholar] [CrossRef]
- Li, Y.; Kong, B.; Xia, X.; Liu, Q.; Diao, X. Structural changes of the myofibrillar proteins in common carp (cyprinus carpio) muscle exposed to a hydroxyl radical-generating system. Process Biochem. 2013, 48, 863–870. [Google Scholar] [CrossRef]
- Xuan, Y.-F.; Zhang, Y.; Zhao, Y.-Y.; Zheng, Z.; Jiang, S.-T.; Zhong, X.-Y. Effect of hydroxypropylmethylcellulose on transition of water status and physicochemical properties of wheat gluten upon frozen storage. Food Hydrocoll. 2017, 63, 35–42. [Google Scholar] [CrossRef]
- Chen, H.; Diao, J.; Li, Y.; Chen, Q.; Kong, B. The effectiveness of clove extracts in the inhibition of hydroxyl radical oxidation-induced structural and rheological changes in porcine myofibrillar protein. Meat Sci. 2016, 111, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Regenstein, J.M.; Zhou, P.; Yang, Y. Effects of high intensity ultrasound modification on physicochemical property and water in myofibrillar protein gel. Ultrason. Sonochem. 2017, 34, 960–967. [Google Scholar] [CrossRef]
- Alinovi, M.; Corredig, M.; Mucchetti, G.; Carini, E. Water status and dynamics of high-moisture Mozzarella cheese as affected by frozen and refrigerated storage. Food Res. Int. 2020, 137, 109415. [Google Scholar] [CrossRef]
- Wang, P.; Xu, L.; Nikoo, M.; Ocen, D.; Wu, F.; Yang, N.; Jin, Z.; Xu, X. Effect of frozen storage on the conformational, thermal and microscopic properties of gluten: Comparative studies on gluten-, glutenin- and gliadin-rich fractions. Food Hydrocoll. 2014, 35, 238–246. [Google Scholar] [CrossRef]
- Deaño, L.C.; Tovar, C.A.; Borderias, J.; Fernandez-Martín, F. Gelation process in two different squid (Dosidicus gigas) surimis throughout frozen storage as affected by several cryoprotectants: Thermal, mechanical and dynamic rheological properties. J. Food Eng. 2011, 107, 107–116. [Google Scholar] [CrossRef]
- Benjakul, S.; Visessanguan, W.; Thongkaew, C.; Tanaka, M. Effect of frozen storage on chemical and gel-forming properties of fish commonly used for surimi production in Thailand. Food Hydrocoll. 2005, 19, 197–207. [Google Scholar] [CrossRef]
- Hu, H.; Wu, J.; Li-Chan, E.C.; Zhu, L.; Zhang, F.; Xu, X.; Fan, G.; Wang, L.; Huang, X.; Pan, S. Effects of ultrasound on structural and physical properties of soy protein isolate (SPI) dispersions. Food Hydrocoll. 2013, 30, 647–655. [Google Scholar] [CrossRef]
- Wu, W.; Hua, Y.; Lin, Q.; Xiao, H. Effects of oxidative modification on thermal aggregation and gel properties of soy protein by peroxyl radicals. Int. J. Food Sci. Technol. 2011, 46, 1891–1897. [Google Scholar] [CrossRef]
- Xue, S.; Xu, X.; Shan, H.; Wang, H.; Yang, J.; Zhou, G. Effects of high-intensity ultrasound, high-pressure processing, and high-pressure homogenization on the physicochemical and functional properties of myofibrillar proteins. Innov. Food Sci. Emerg. Technol. 2018, 45, 354–360. [Google Scholar] [CrossRef]
- Du, L.; Betti, M. Chicken collagen hydrolysate cryoprotection of natural actomyosin: Mechanism studies during freeze-thaw cycles and simulated digestion. Food Chem. 2016, 211, 791–802. [Google Scholar] [CrossRef]
- Shenouda, S.Y. Theories of Protein Denaturation During Frozen Storage of Fish Flesh. Adv. Food Res. 1980, 26, 275–311. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Storage Time (Day) | |||||
|---|---|---|---|---|---|
| 0 | 30 | 60 | 90 | 180 | |
| AF | 43.9 ± 0.7 Ea | 60.2 ± 0.7 Da | 67.7 ± 0.6 Ca | 81.6 ± 0.9 Ba | 113.4 ± 1.0 Aa |
| IF | 29.3 ± 0.9 Eb | 38.3 ± 0.9 Db | 42.5 ± 0.7 Cb | 54.5 ± 0.8 Bb | 109.5 ± 0.9 Ab |
| UF | 27.9 ± 0.6 Eb | 31.2 ± 0.4 Dc | 34.6 ± 0.5 Cc | 38.8 ± 0.7 Bc | 96.3 ± 0.8 Ac |
| Storage Time (Day) | ||||||
|---|---|---|---|---|---|---|
| 0 | 30 | 60 | 90 | 180 | ||
| A2b1 | AF | 0.32 ± 0.01 Ab | 0.32 ± 0.01 Aa | 0.28 ± 0.01 Ba | 0.25 ± 0.01 Ca | 0.24 ± 0.01 Ca |
| IF | 0.34 ± 0.01 Ab | 0.33 ± 0.01 Aa | 0.29 ± 0.01 Ba | 0.25 ± 0.01 Ca | 0.24 ± 0.01 Ca | |
| UF | 0.38 ± 0.01 Aa | 0.34 ± 0.01 Ba | 0.29 ± 0.01 Ca | 0.26 ± 0.01 Da | 0.24 ± 0.01 Da | |
| A2b1 | AF | 0.33 ± 0.01 Ac | 0.31 ± 0.01 ABb | 0.29 ± 0.01 BCb | 0.26 ± 0.01 CDa | 0.25 ± 0.01 Da |
| IF | 0.35 ± 0.01 Ab | 0.32 ± 0.01 Bb | 0.29 ± 0.01 Cb | 0.26 ± 0.01 Da | 0.26 ± 0.01 Da | |
| UF | 0.39 ± 0.01 Aa | 0.35 ± 0.01 Ba | 0.31 ± 0.01 Ca | 0.27 ± 0.01 Da | 0.26 ± 0.01 Ea | |
| A21 | AF | 92.62 ± 0.07 Ac | 90.87 ± 0.17 Bc | 87.13 ± 0.09 Cc | 85.83 ± 0.10 Dc | 81.4 ± 0.07 Ec |
| IF | 93.00 ± 0.06 Ab | 92.25 ± 0.03 Bb | 88.09 ± 0.08 Cb | 86.44 ± 0.10 Db | 82.78 ± 0.09 Eb | |
| UF | 93.35 ± 0.04 Aa | 92.57 ± 0.09 Ba | 89.58 ± 0.07 Ca | 87.82 ± 0.13 Da | 84.75 ± 0.11 Ea | |
| A22 | AF | 6.73 ± 0.09 Ac | 8.41 ± 0.07 Bc | 12.33 ± 0.05 Cc | 13.69 ± 0.07 Dc | 18.11 ± 0.05 Ec |
| IF | 6.31 ± 0.03 Ab | 7.11 ± 0.06 Bb | 11.32 ± 0.04 Cb | 13.03 ± 0.03 Db | 16.75 ± 0.08 Eb | |
| UF | 5.83 ± 0.04 Aa | 6.75 ± 0.07 Ba | 9.77 ± 0.09 Ca | 11.69 ± 0.03 Da | 14.72 ± 0.06 Ea | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Q.; Kong, B.; Liu, S.; Zheng, O.; Zhang, C. Ultrasonic Freezing Reduces Protein Oxidation and Myofibrillar Gel Quality Loss of Common Carp (Cyprinus carpio) during Long-Time Frozen Storage. Foods 2021, 10, 629. https://doi.org/10.3390/foods10030629
Sun Q, Kong B, Liu S, Zheng O, Zhang C. Ultrasonic Freezing Reduces Protein Oxidation and Myofibrillar Gel Quality Loss of Common Carp (Cyprinus carpio) during Long-Time Frozen Storage. Foods. 2021; 10(3):629. https://doi.org/10.3390/foods10030629
Chicago/Turabian StyleSun, Qinxiu, Baohua Kong, Shucheng Liu, Ouyang Zheng, and Chao Zhang. 2021. "Ultrasonic Freezing Reduces Protein Oxidation and Myofibrillar Gel Quality Loss of Common Carp (Cyprinus carpio) during Long-Time Frozen Storage" Foods 10, no. 3: 629. https://doi.org/10.3390/foods10030629