
PRF Lysates Enhance the Proliferation and Migration of Oral Squamous Carcinoma Cell Lines


Abstract
:1. Introduction
2. Material and Methods
2.1. Oral Squamous Cell Carcinoma Cell Lines HSC2, TR146, and Primary Cells
2.2. PRF Lysates, Buffy Coat, and Platelet-Poor Plasma
2.3. RNA Sequencing
2.4. Reverse Transcription Quantitative Real-Time PCR
2.5. Proliferation Assay
2.6. Scratch Assay
2.7. Statistical Analysis
3. Results
3.1. RNAseq Analysis
3.2. RT-PCR Analysis
3.3. 3[H]-Thymidine Incorporations and Cell Counting
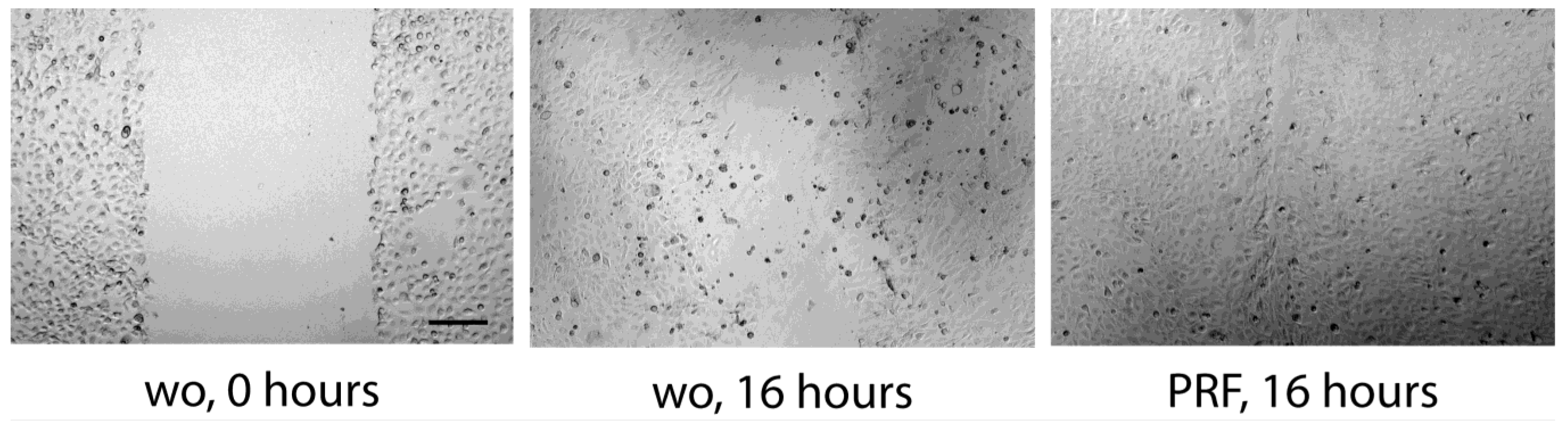
3.4. Scratch Assay
4. Discussion
5. Conclusions and Future Research
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dohan, D.M.; Choukroun, J.; Diss, A.; Dohan, S.L.; Dohan, A.J.; Mouhyi, J.; Gogly, B. Platelet-rich fibrin (PRF): A second-generation platelet concentrate. Part I: Technological concepts and evolution. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2006, 101, e37–e44. [Google Scholar] [CrossRef]
- Castro, A.B.; Van Dessel, J.; Temmerman, A.; Jacobs, R.; Quirynen, M. Effect of different platelet-rich fibrin matrices for ridge preservation in multiple tooth extractions: A split-mouth randomized controlled clinical trial. J. Clin. Periodontol. 2021, 48, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Temmerman, A.; Vandessel, J.; Castro, A.; Jacobs, R.; Teughels, W.; Pinto, N.; Quirynen, M. The use of leucocyte and platelet-rich fibrin in socket management and ridge preservation: A split-mouth, randomized, controlled clinical trial. J. Clin. Periodontol. 2016, 43, 990–999. [Google Scholar] [CrossRef] [PubMed]
- Cortellini, S.; Castro, A.B.; Temmerman, A.; Van Dessel, J.; Pinto, N.; Jacobs, R.; Quirynen, M. Leucocyte- and platelet-rich fibrin block for bone augmentation procedure: A proof-of-concept study. J. Clin. Periodontol. 2018, 45, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Miron, R.J.; Moraschini, V.; Fujioka-Kobayashi, M.; Zhang, Y.; Kawase, T.; Cosgarea, R.; Jepsen, S.; Bishara, M.; Canullo, L.; Shirakata, Y.; et al. Use of platelet-rich fibrin for the treatment of periodontal intrabony defects: A systematic review and meta-analysis. Clin. Oral Investig. 2021, 25, 2461–2478. [Google Scholar] [CrossRef] [PubMed]
- Miron, R.J.; Moraschini, V.; Del Fabbro, M.; Piattelli, A.; Fujioka-Kobayashi, M.; Zhang, Y.; Saulacic, N.; Schaller, B.; Kawase, T.; Cosgarea, R.; et al. Use of platelet-rich fibrin for the treatment of gingival recessions: A systematic review and meta-analysis. Clin. Oral Investig. 2020, 24, 2543–2557. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Tomikawa, K.; Yamamoto, T.; Shiomi, N.; Shimoe, M.; Hongo, S.; Yamashiro, K.; Yamaguchi, T.; Maeda, H.; Takashiba, S. Smad2 decelerates re-epithelialization during gingival wound healing. J. Dent. Res. 2012, 91, 764–770. [Google Scholar] [CrossRef]
- Mohanty, S.; Pathak, H.; Dabas, J. Platelet rich fibrin: A new covering material for oral mucosal defects. J. Oral Biol. Craniofac. Res. 2014, 4, 144–146. [Google Scholar] [CrossRef]
- Paulmann, A.; Strauss, S.; Limbourg, A.; Vogt, P.M. Platelet-derived concentrates influence human keratinocyte proliferation in vitro and induce wound healing in a prospective case series of chronic wounds of different entities in vivo. Innov. Surg. Sci. 2022, 7, 45–58. [Google Scholar] [CrossRef]
- Jafar, H.; Hasan, M.; Al-Hattab, D.; Saleh, M.; Ameereh, L.A.; Khraisha, S.; Younes, N.; Awidi, A. Platelet lysate promotes the healing of long-standing diabetic foot ulcers: A report of two cases and in vitro study. Heliyon 2020, 6, e03929. [Google Scholar] [CrossRef] [PubMed]
- Bayer, A.; Tohidnezhad, M.; Berndt, R.; Lippross, S.; Behrendt, P.; Kluter, T.; Pufe, T.; Jahr, H.; Cremer, J.; Rademacher, F.; et al. Platelet-released growth factors inhibit proliferation of primary keratinocytes in vitro. Ann. Anat. 2018, 215, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, M.; Oliva, A.; Buonaiuto, C.; Di Feo, A.; Di Pasquale, R.; Passaro, I.; Guida, L. In vitro cell-type specific biological response of human periodontally related cells to platelet-rich plasma. J. Periodontal Res. 2005, 40, 489–495. [Google Scholar] [CrossRef]
- Okuda, K.; Kawase, T.; Momose, M.; Murata, M.; Saito, Y.; Suzuki, H.; Wolff, L.F.; Yoshie, H. Platelet-rich plasma contains high levels of platelet-derived growth factor and transforming growth factor-beta and modulates the proliferation of periodontally related cells in vitro. J. Periodontol. 2003, 74, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Feller, L.L.; Khammissa, R.R.; Kramer, B.B.; Lemmer, J.J. Oral squamous cell carcinoma in relation to field precancerisation: Pathobiology. Cancer Cell Int. 2013, 13, 31. [Google Scholar] [CrossRef]
- Ganesh, D.; Sreenivasan, P.; Ohman, J.; Wallstrom, M.; Braz-Silva, P.H.; Giglio, D.; Kjeller, G.; Hasseus, B. Potentially Malignant Oral Disorders and Cancer Transformation. Anticancer Res. 2018, 38, 3223–3229. [Google Scholar] [CrossRef]
- Rivera, C. Essentials of oral cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 11884–11894. [Google Scholar]
- Jiang, S.; Dong, Y. Human papillomavirus and oral squamous cell carcinoma: A review of HPV-positive oral squamous cell carcinoma and possible strategies for future. Curr. Probl. Cancer 2017, 41, 323–327. [Google Scholar] [CrossRef]
- Villa, A.; Sonis, S. Oral leukoplakia remains a challenging condition. Oral Dis. 2018, 24, 179–183. [Google Scholar] [CrossRef]
- Kurago, Z.B. Etiology and pathogenesis of oral lichen planus: An overview. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2016, 122, 72–80. [Google Scholar] [CrossRef]
- Duan, W.; Wu, Z.; Mei, X.; Li, J.; Lu, H.; Liang, Y.; Lao, X.; Liao, G. Intratumoral platelet microthrombi in oral squamous cell carcinoma: A marker of lymph node metastasis. Oral Dis. 2023. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Cui, W.; Pei, Y.; Xu, D. Platelets promote invasion and induce epithelial to mesenchymal transition in ovarian cancer cells by TGF-beta signaling pathway. Gynecol. Oncol. 2019, 153, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, Y.; Li, D.; Zhang, L.; Wang, K.; Zuo, Y.; Gartner, T.K.; Liu, J. Platelet P2Y12 is involved in murine pulmonary metastasis. PLoS ONE 2013, 8, e80780. [Google Scholar] [CrossRef]
- Haemmerle, M.; Taylor, M.L.; Gutschner, T.; Pradeep, S.; Cho, M.S.; Sheng, J.; Lyons, Y.M.; Nagaraja, A.S.; Dood, R.L.; Wen, Y.; et al. Platelets reduce anoikis and promote metastasis by activating YAP1 signaling. Nat. Commun. 2017, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Gay, L.J.; Felding-Habermann, B. Platelets alter tumor cell attributes to propel metastasis: Programming in transit. Cancer Cell 2011, 20, 553–554. [Google Scholar] [CrossRef]
- Park, S.Y.; Lee, S.K.; Lim, M.; Kim, B.; Hwang, B.O.; Cho, E.S.; Zhang, X.; Chun, K.S.; Chung, W.Y.; Song, N.Y. Direct Contact with Platelets Induces Podoplanin Expression and Invasion in Human Oral Squamous Cell Carcinoma Cells. Biomol. Ther. 2022, 30, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Panahipour, L.; Oladzad Abbasabadi, A.; Gruber, R. Oral Cell Lysates Reduce the Inflammatory Response of Activated Macrophages. J. Clin. Med. 2023, 12, 1701. [Google Scholar] [CrossRef] [PubMed]
- Panahipour, L.; Nasserzare, S.; Amer, Z.; Brucke, F.; Stahli, A.; Kreissl, A.; Haiden, N.; Gruber, R. The anti-inflammatory effect of milk and dairy products on periodontal cells: An in vitro approach. Clin. Oral Investig. 2019, 23, 1959–1966. [Google Scholar] [CrossRef]
- Panahipour, L.; Moghaddam, D.M.; Nasirzade, J.; Kargarpour, Z.; Gruber, R. RNAseq of TGF-beta receptor type I kinase-dependent genes in oral fibroblast exposed to milk. BMC Oral Health 2021, 21, 581. [Google Scholar] [CrossRef] [PubMed]
- Goodlad, R.A. Quantification of epithelial cell proliferation, cell dynamics, and cell kinetics in vivo. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6, e274. [Google Scholar] [CrossRef]
- Polo, S.E.; Almouzni, G. Histone metabolic pathways and chromatin assembly factors as proliferation markers. Cancer Lett. 2005, 220, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gerlitz, G. The Emerging Roles of Heterochromatin in Cell Migration. Front. Cell Dev. Biol. 2020, 8, 394. [Google Scholar] [CrossRef] [PubMed]
- Bayer, A.; Wijaya, B.; Mobus, L.; Rademacher, F.; Rodewald, M.; Tohidnezhad, M.; Pufe, T.; Drucke, D.; Glaser, R.; Harder, J. Platelet-Released Growth Factors and Platelet-Rich Fibrin Induce Expression of Factors Involved in Extracellular Matrix Organization in Human Keratinocytes. Int. J. Mol. Sci. 2020, 21, 4404. [Google Scholar] [CrossRef] [PubMed]
- Misiura, M.; Guszczyn, T.; Oscilowska, I.; Baszanowska, W.; Palka, J.; Miltyk, W. Platelet-Rich Plasma Promotes the Proliferation of Human Keratinocytes via a Progression of the Cell Cycle. A Role of Prolidase. Int. J. Mol. Sci. 2021, 22, 936. [Google Scholar] [CrossRef] [PubMed]
- Duan, G.H.; Ren, Z.Q.; Du, B.; Shao, W.; Dong, H.J.; Du, A.C. Platelet-rich plasma protects human keratinocytes from UVB-induced apoptosis by attenuating inflammatory responses and endoplasmic reticulum stress. J. Cosmet. Dermatol. 2023, 22, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-H.; Shen, S.-Y.; Zhao, J.-H.; Chang, Y.-C. Platelet-rich fibrin modulates cell proliferation of human periodontally related cells in vitro. J. Dental. Sci. 2009, 4, 130–135. [Google Scholar] [CrossRef]
- Carney, B.C.; Simbulan-Rosenthal, C.M.; Gaur, A.; Browne, B.J.; Moghe, M.; Crooke, E.; Moffatt, L.T.; Shupp, J.W.; Rosenthal, D.S. Inorganic polyphosphate in platelet rich plasma accelerates re-epithelialization in vitro and in vivo. Regen. Ther. 2020, 15, 138–148. [Google Scholar] [CrossRef]
- Xu, Y.; Lin, Z.; He, L.; Qu, Y.; Ouyang, L.; Han, Y.; Xu, C.; Duan, D. Platelet-Rich Plasma-Derived Exosomal USP15 Promotes Cutaneous Wound Healing via Deubiquitinating EIF4A1. Oxid. Med. Cell. Longev. 2021, 2021, 9674809. [Google Scholar] [CrossRef]
- Lovisolo, F.; Carton, F.; Gino, S.; Migliario, M.; Reno, F. Platelet rich plasma-derived microvesicles increased in vitro wound healing. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 9658–9664. [Google Scholar] [CrossRef]
- Yang, S.C.; Lin, C.F.; Alshetaili, A.; Aljuffali, I.A.; Chien, M.Y.; Fang, J.Y. Combining the dual antibacterial and regenerative activities of platelet-rich plasma with beta-lactams to mitigate MRSA-infected skin wounds. Biomed. Pharmacother. 2023, 165, 115017. [Google Scholar] [CrossRef]
- Leisi, S.; Farahpour, M.R. Effectiveness of topical administration of platelet-rich plasma on the healing of methicillin-resistant Staphylococcus aureus-infected full-thickness wound model. J. Plast. Reconstr. Aesthet. Surg. 2023, 77, 416–429. [Google Scholar] [CrossRef]
- Feuerherm, A.J.; Jorgensen, K.M.; Sommerfelt, R.M.; Eidem, L.E.; Laegreid, A.; Johansen, B. Platelet-activating factor induces proliferation in differentiated keratinocytes. Mol. Cell. Biochem. 2013, 384, 83–94. [Google Scholar] [CrossRef]
- White, S.R.; Fischer, B.M.; Marroquin, B.A.; Stern, R. Interleukin-1beta mediates human airway epithelial cell migration via NF-kappaB. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 295, L1018–L1027. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.C.; Lee, I.T.; Lin, C.C.; Chi, P.L.; Cheng, S.E.; Shih, R.H.; Hsiao, L.D.; Yang, C.M. IL-1beta promotes corneal epithelial cell migration by increasing MMP-9 expression through NF-kappaB- and AP-1-dependent pathways. PLoS ONE 2013, 8, e57955. [Google Scholar] [CrossRef]
- Masola, V.; Carraro, A.; Granata, S.; Signorini, L.; Bellin, G.; Violi, P.; Lupo, A.; Tedeschi, U.; Onisto, M.; Gambaro, G.; et al. In vitro effects of interleukin (IL)-1 beta inhibition on the epithelial-to-mesenchymal transition (EMT) of renal tubular and hepatic stellate cells. J. Transl. Med. 2019, 17, 12. [Google Scholar] [CrossRef] [PubMed]
- Momose, F.; Araida, T.; Negishi, A.; Ichijo, H.; Shioda, S.; Sasaki, S. Variant sublines with different metastatic potentials selected in nude mice from human oral squamous cell carcinomas. J. Oral Pathol. Med. 1989, 18, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Ichijo, H.; Momose, F.; Miyazono, K. Biological effects and binding properties of transforming growth factor-beta on human oral squamous cell carcinoma cells. Exp. Cell Res. 1990, 187, 263–269. [Google Scholar] [CrossRef]
- Lalla, R.V.; Saunders, D.P.; Peterson, D.E. Chemotherapy or radiation-induced oral mucositis. Dent. Clin. N. Am. 2014, 58, 341–349. [Google Scholar] [CrossRef]
- Sundaram, G.M.; Quah, S.; Sampath, P. Cancer: The dark side of wound healing. FEBS J. 2018, 285, 4516–4534. [Google Scholar] [CrossRef]
- Boyd, F.T.; Massague, J. Transforming growth factor-beta inhibition of epithelial cell proliferation linked to the expression of a 53-kDa membrane receptor. J. Biol. Chem. 1989, 264, 2272–2278. [Google Scholar] [CrossRef]
- Kargarpour, Z.; Panahipour, L.; Mildner, M.; Miron, R.J.; Gruber, R. Lipids of Platelet-Rich Fibrin Reduce the Inflammatory Response in Mesenchymal Cells and Macrophages. Cells 2023, 12, 634. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Sequence | Reverse Sequence |
|---|---|---|
| CDC45 | GAAGCGCACACGGTTAGAA | GTTCACTCCCAGAGCCACTC |
| CyclinD1 | TCGGTGTCCTACTTCAAATGTGT | GGGATGGTCTCCTTCATCTTAG |
| E2F1 | GGACCTGGAAACTGACCATCAG | CAGTGAGGTCTCATAGCGTGAC |
| FAM111B | GGTTCTGGGGAAATCCAGTC | AAGATGGAGAAACAAGGATTGAA |
| HIST1H1A | GGCAAAGAAACCTGCTAAGGCTG | TAAGAGCTGCCAACGACACACC |
| HIST1H1D | GCTTATCACCAAGGCAGTGGCA | CCAGAGTACCTTTGCTCACCAAG |
| HIST1H2AJ | CGACAACAAGAAGACTCGCATCA | TGTGCGATGGTGACTTTGCCCA |
| IL1b | ATGATGGCTTATTACAGTGGCAA | GTCGGAGATTCGTAGCTGGA |
| Ki-67 | ATAAACACCCCAACACACACAA | GCCACTTCTTCATTCCAGTTACA |
| MCM10 | CATGAAGCCCAAGGATGG | GACCTTCTGAGGATGATCGATAG |
| PCNA | TGGAGAACTTGGAAATGGAAAC | GAACTGGTTCATTCATCTCTATGG |
| SULT1B1 | CCAACTACAGTGATGGATCATAGC | GCGGAATTGAAGTGCAGTTTTGG |
| Upregulated Genes | Downregulated Genes | ||
|---|---|---|---|
| HIST1H1A | 83.0 | FOXA1 | 21.4 |
| IL1B | 59.5 | KCNH4 | 19.6 |
| E2F1 | 56.1 | SAMD11 | 18.9 |
| HIST1H1D | 43.3 | DPP7 | 18.7 |
| SULT1B1 | 35.9 | TCHHL1 | 18.4 |
| HIST1H2AJ | 33.7 | IFI44L | 16.9 |
| XYLT1 | 24.7 | RGS16 | 16.5 |
| MMP9 | 24.7 | ROM1 | 16.0 |
| NAV3 | 23.6 | SH2D3C | 14.9 |
| OTUB2 | 22.4 | PIFO | 14.3 |
| ZNF367 | 21.6 | BLNK | 14.3 |
| ORC1 | 21.3 | CACNA1A | 14.3 |
| LRRC75B | 20.2 | KRTAP19-1 | 13.4 |
| EXO1 | 19.3 | ATOH8 | 12.5 |
| CAMP | 19.1 | S1PR1 | 12.5 |
| CDCA7 | 18.2 | CAMK2A | 12.0 |
| CLCF1 | 16.8 | C10orf99 | 11.8 |
| CCNE2 | 16.8 | TMPRSS11E | 11.6 |
| F2R | 16.5 | TTLL3 | 11.6 |
| SLCO4A1 | 16.3 | PDE3A | 11.3 |
| ETV5 | 16.3 | TMEM86A | 11.1 |
| FAM72D | 15.7 | SMAD9 | 10.9 |
| IL1R2 | 14.9 | ZNF691 | 10.7 |
| MCM10 | 14.8 | RCAN2 | 10.7 |
| RAET1L | 14.6 | ACAA2 | 10.7 |
| HIST1H2BF | 14.6 | CCL26 | 10.7 |
| HIST2H2AC | 14.6 | WDR93 | 10.7 |
| FAM111B | 13.6 | RBBP8NL | 10.7 |
| IL1A | 13.5 | SLC41A2 | 10.7 |
| GPSM3 | 13.5 | TM7SF2 | 10.6 |
| HIST1H4C | 12.9 | NUPR1 | 10.6 |
| PLXNA4 | 12.8 | GBP2 | 10.4 |
| MAMDC2 | 12.3 | PAPLN | 10.1 |
| CDH11 | 12.3 | SERPINI1 | 10.1 |
| SPRED1 | 12.3 | ||
| AMIGO2 | 12.2 | ||
| AREG | 11.7 | ||
| VEPH1 | 11.5 | ||
| VAX1 | 11.2 | ||
| MYBPHL | 11.2 | ||
| KLF14 | 11.2 | ||
| GLI2 | 11.2 | ||
| ANKRD44 | 11.2 | ||
| IL1RN | 10.9 | ||
| CLMP | 10.7 | ||
| APCDD1L | 10.4 | ||
| CDC45 | 10.3 | ||
| GPR68 | 10.1 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panahipour, L.; Croci, R.; Guarnieri, S.; Gruber, R. PRF Lysates Enhance the Proliferation and Migration of Oral Squamous Carcinoma Cell Lines. Dent. J. 2023, 11, 242. https://doi.org/10.3390/dj11100242
Panahipour L, Croci R, Guarnieri S, Gruber R. PRF Lysates Enhance the Proliferation and Migration of Oral Squamous Carcinoma Cell Lines. Dentistry Journal. 2023; 11(10):242. https://doi.org/10.3390/dj11100242
Chicago/Turabian StylePanahipour, Layla, Rebecca Croci, Sara Guarnieri, and Reinhard Gruber. 2023. "PRF Lysates Enhance the Proliferation and Migration of Oral Squamous Carcinoma Cell Lines" Dentistry Journal 11, no. 10: 242. https://doi.org/10.3390/dj11100242