
Oral Microbiome and Dental Caries Development




Abstract
:1. Introduction
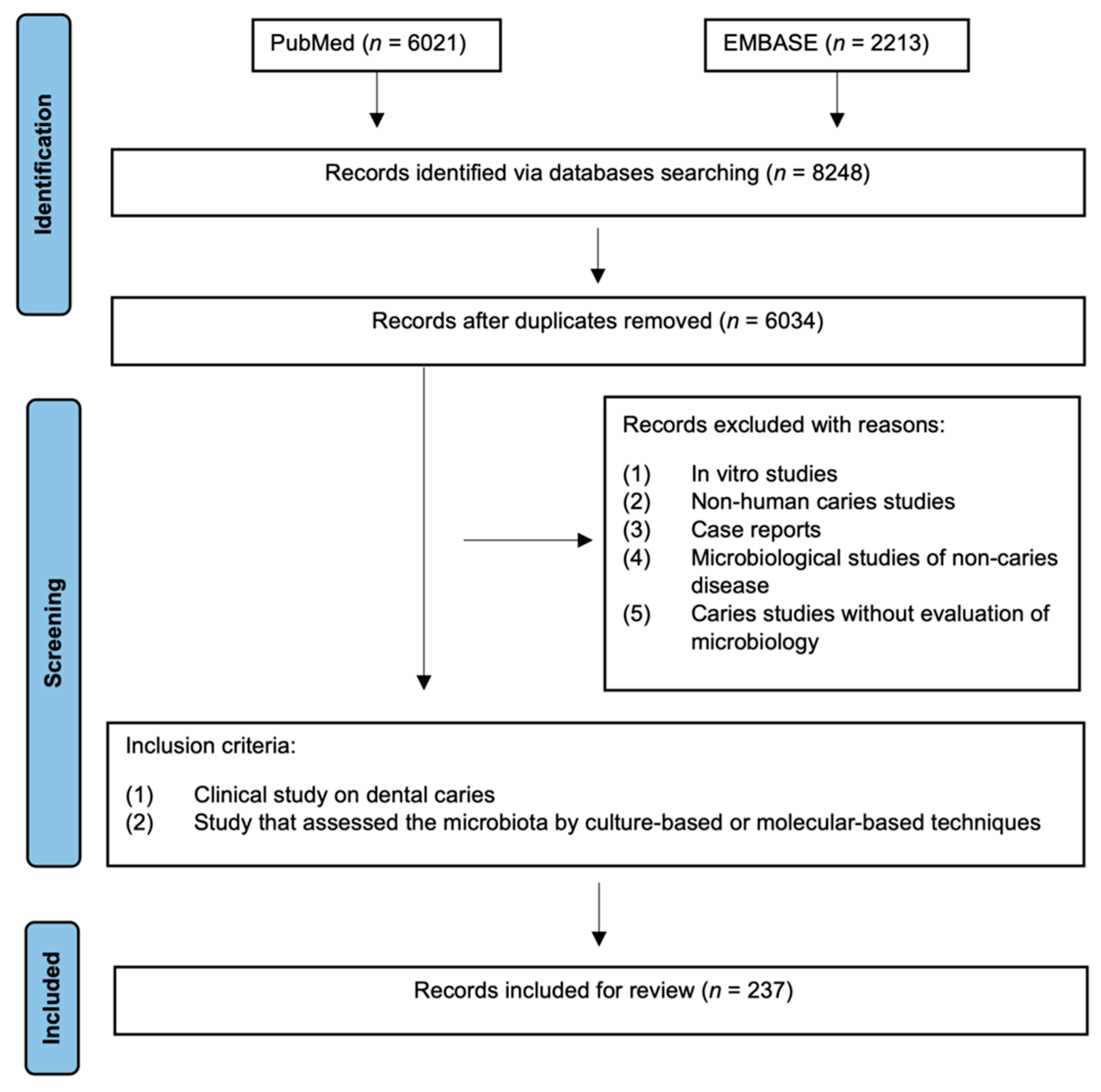
2. Materials and Methods
3. Results
3.1. Oral Microbial Communities
3.2. The Microbiological Hypothesis of Dental Caries Etiology
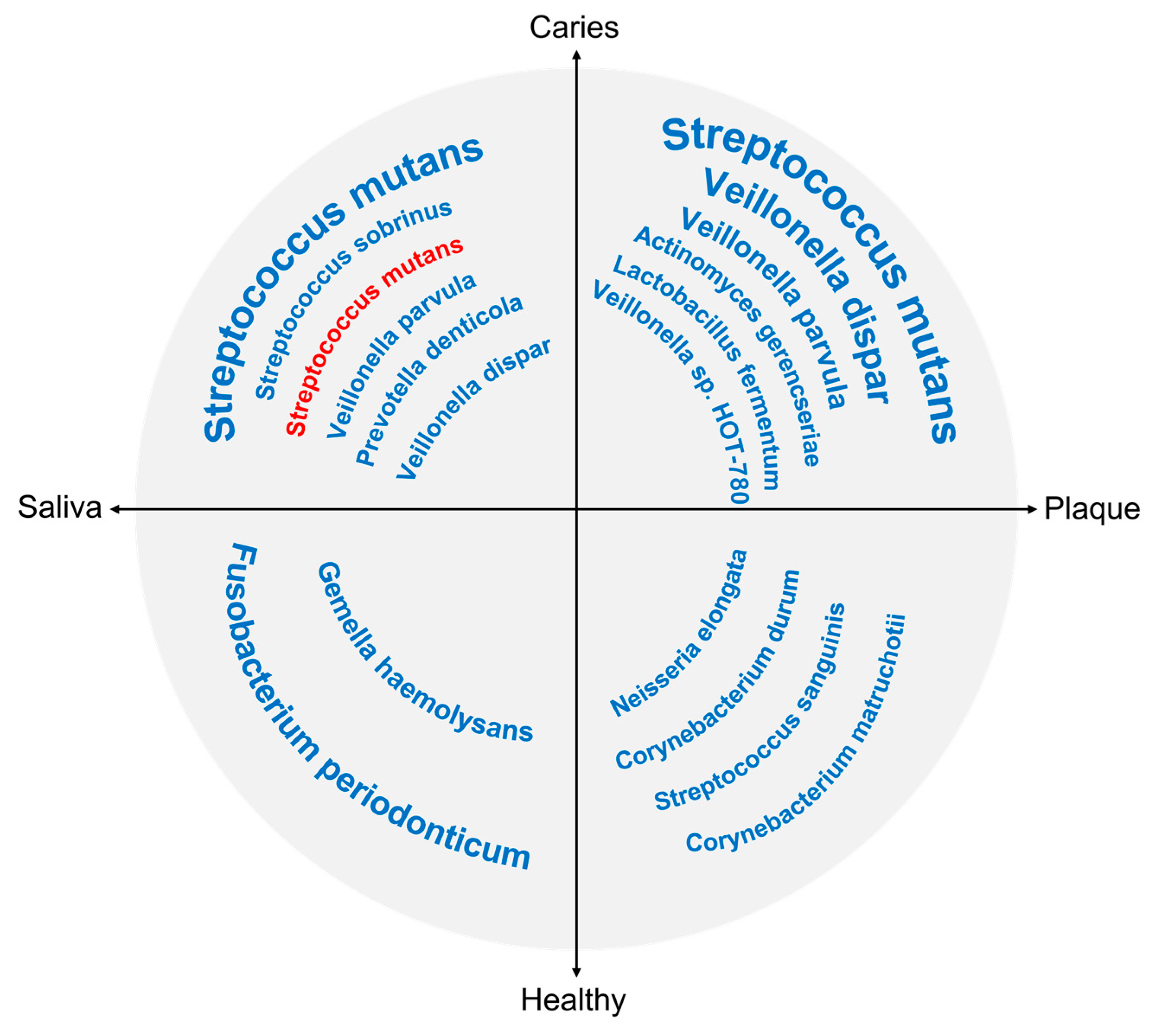
3.3. Microorganisms Associated with Caries Development
3.4. Shifts in the Oral Microbiome in Dental Caries
3.4.1. Microbiome Shifts in Caries of Different Stages
3.4.2. Microbiome Shifts in Caries with Different Activities
3.4.3. Microbiome Shifts in Caries in Different Locations
4. Discussion and Future Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kassebaum, N.; Smith, A. Global, Regional, and National Prevalence, Incidence, and Disability-Adjusted Life Years for Oral Conditions for 195 Countries, 1990–2015: A Systematic Analysis for the Global Burden of Diseases, Injuries, and Risk Factors. J. Dent. Res. 2017, 96, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Yu, O.Y.; Lam, W.Y.; Wong, A.W.; Duangthip, D.; Chu, C.H. Nonrestorative Management of Dental Caries. Dent. J. 2021, 9, 121. [Google Scholar] [CrossRef] [PubMed]
- De Soet, J.J.; van Gemert-Schriks, M.C. Host and microbiological factors related to dental caries development. Caries Res. 2008, 42, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Tanner, A.; Kressirer, C. The Caries Microbiome: Implications for Reversing Dysbiosis. Adv. Dent. Res. 2018, 29, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Schüpbach, P.; Osterwalder, V. Human root caries: Microbiota in plaque covering sound, carious and arrested carious root surfaces. Caries Res. 1995, 29, 382–395. [Google Scholar] [CrossRef] [PubMed]
- Percival, R.; Challacombe, S. Age-related microbiological changes in the salivary and plaque microflora of healthy adults. J. Med. Microbiol. 1991, 35, 5–11. [Google Scholar] [CrossRef]
- Vartoukian, S.R.; Palmer, R.M.; Wade, W.G. Strategies for culture of ‘unculturable’ bacteria. FEMS Microbiol. Lett. 2010, 309, 1–7. [Google Scholar] [CrossRef]
- Bleiweis, A.S.; Oyston, P.C.; Brady, L.J. Molecular, immunological and functional characterization of the major surface adhesin of Streptococcus mutans. Adv. Exp. Med. Biol. 1992, 327, 229–241. [Google Scholar]
- Colby, S.; Harrington, D. Identification and genetic characterisation of melibiose-negative isolates of Streptococcus mutans. Caries Res. 1995, 29, 407–412. [Google Scholar] [CrossRef]
- Socransky, S.; Smith, C. “Checkerboard” DNA-DNA hybridization. BioTechniques 1994, 17, 788–792. [Google Scholar]
- Woo, P.C.; Lau, S.K. Then and now: Use of 16S rDNA gene sequencing for bacterial identification and discovery of novel bacteria in clinical microbiology laboratories. Clin. Microbiol. Infect. 2008, 14, 908–934. [Google Scholar] [CrossRef]
- Johnson, J.S.; Spakowicz, D.J. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [Green Version]
- Van Rossum, T.; Ferretti, P.; Maistrenko, O.M.; Bork, P. Diversity within species: Interpreting strains in microbiomes. Nat. Rev. Microbiol. 2020, 18, 491–506. [Google Scholar] [CrossRef]
- Simón-Soro, A.; Mira, A. Solving the etiology of dental caries. Trends Microbiol. 2015, 23, 76–82. [Google Scholar] [CrossRef]
- Tong, X.; Hou, S. The integration of transcriptome-wide association study and mRNA expression profiling data to identify candidate genes and gene sets associated with dental caries. Arch. Oral Biol. 2020, 118, 104863. [Google Scholar] [CrossRef]
- Edlund, A.; Yang, Y. Meta-omics uncover temporal regulation of pathways across oral microbiome genera during in vitro sugar metabolism. ISME J. 2015, 9, 2605–2619. [Google Scholar] [CrossRef]
- Verma, D.; Garg, P.K.; Dubey, A.K. Insights into the human oral microbiome. Arch. Microbiol. 2018, 200, 525–540. [Google Scholar] [CrossRef]
- Horz, H. Archaeal Lineages within the Human Microbiome: Absent, Rare or Elusive? Life 2015, 5, 1333–1345. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X. Human oral microbiota and its modulation for oral health. Biomed. Pharmacother. 2018, 99, 883–893. [Google Scholar] [CrossRef]
- Beighton, D.; Lynch, E. Relationships between yeasts and primary root-caries lesions. Gerodontology 1993, 10, 105–108. [Google Scholar] [CrossRef]
- Bandara, H.; Panduwawala, C. Biodiversity of the human oral mycobiome in health and disease. Oral Dis. 2019, 25, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Diaz, P.I.; Dongari-Bagtzoglou, A. Critically Appraising the Significance of the Oral Mycobiome. J. Dent. Res. 2021, 100, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.; Hoare, A. The Salivary Mycobiome Contains 2 Ecologically Distinct Mycotypes. J. Dent. Res. 2020, 99, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.; Jurevic, R. Characterization of the oral fungal microbiome (mycobiome) in healthy individuals. PLoS Pathog. 2010, 6, e1000713. [Google Scholar] [CrossRef]
- Pereira, D.; Seneviratne, C. Is the oral fungal pathogen Candida albicans a cariogen? Oral Dis. 2018, 24, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Canabarro, A.; Valle, C. Association of subgingival colonization of Candida albicans and other yeasts with severity of chronic periodontitis. J. Periodontal Res. 2013, 48, 428–432. [Google Scholar] [CrossRef]
- Brusa, T.; Conca, R. The presence of methanobacteria in human subgingival plaque. J. Clin. Periodontol. 1987, 14, 470–471. [Google Scholar] [CrossRef]
- Belay, N.; Johnson, R. Methanogenic bacteria from human dental plaque. Appl. Environ. Microbiol. 1988, 54, 600–603. [Google Scholar] [CrossRef]
- Lepp, P.; Brinig, M. Methanogenic Archaea and human periodontal disease. Proc. Natl. Acad. Sci. USA 2004, 101, 6176–6181. [Google Scholar] [CrossRef]
- Faveri, M.; Gonçalves, L. Prevalence and microbiological diversity of Archaea in peri-implantitis subjects by 16S ribosomal RNA clonal analysis. J. Periodontal Res. 2011, 46, 338–344. [Google Scholar] [CrossRef]
- Horz, H.; Robertz, N. Relationship between methanogenic archaea and subgingival microbial complexes in human periodontitis. Anaerobe 2015, 35, 10–12. [Google Scholar] [CrossRef]
- Horz, H.; Seyfarth, I. McrA and 16S rRNA gene analysis suggests a novel lineage of Archaea phylogenetically affiliated with Thermoplasmatales in human subgingival plaque. Anaerobe 2012, 18, 373–377. [Google Scholar] [CrossRef]
- Kulik, E.; Sandmeier, H. Identification of archaeal rDNA from subgingival dental plaque by PCR amplification and sequence analysis. FEMS Microbiol. Lett. 2001, 196, 129–133. [Google Scholar] [CrossRef]
- Brzezińska-Błaszczyk, E.; Pawłowska, E. Presence of archaea and selected bacteria in infected root canal systems. Can. J. Microbiol. 2018, 64, 317–326. [Google Scholar] [CrossRef]
- Efenberger, M.; Agier, J. Archaea prevalence in inflamed pulp tissues. Cent. Eur. J. Immunol. 2015, 40, 194–200. [Google Scholar] [CrossRef]
- Dame-Teixeira, N.; de Cena, J. Presence of Archaea in dental caries biofilms. Arch. Oral Biol. 2020, 110, 104606. [Google Scholar] [CrossRef]
- Guindo, C.; Terrer, E. Culture of salivary methanogens assisted by chemically produced hydrogen. Anaerobe 2020, 61, 102128. [Google Scholar] [CrossRef]
- Göhler, A.; Samietz, S. Comparison of Oral Microbe Quantities from Tongue Samples and Subgingival Pockets. Int. J. Dent. 2018, 2018, 2048390. [Google Scholar]
- Belmok, A.; de Cena, J. The Oral Archaeome: A Scoping Review. J. Dent. Res. 2020, 99, 630–643. [Google Scholar] [CrossRef]
- Vianna, M.; Conrads, G. Identification and quantification of archaea involved in primary endodontic infections. J. Clin. Microbiol. 2006, 44, 1274–1282. [Google Scholar] [CrossRef]
- Baker, J.; Bor, B. Ecology of the Oral Microbiome: Beyond Bacteria. Trends Microbiol. 2017, 25, 362–374. [Google Scholar] [CrossRef]
- Abeles, S.; Robles-Sikisaka, R. Human oral viruses are personal, persistent and gender-consistent. ISME J. 2014, 8, 1753–1767. [Google Scholar] [CrossRef]
- Naidu, M.; Robles-Sikisaka, R. Characterization of bacteriophage communities and CRISPR profiles from dental plaque. BMC Microbiol. 2014, 14, 175. [Google Scholar] [CrossRef] [Green Version]
- Ly, M.; Abeles, S. Altered oral viral ecology in association with periodontal disease. mBio 2014, 5, e01133-14. [Google Scholar] [CrossRef]
- Pride, D.; Salzman, J. Evidence of a robust resident bacteriophage population revealed through analysis of the human salivary virome. ISME J. 2012, 6, 915–926. [Google Scholar] [CrossRef]
- Doel, J.; Benjamin, N. Evaluation of bacterial nitrate reduction in the human oral cavity. Eur. J. Oral Sci. 2005, 113, 14–19. [Google Scholar] [CrossRef]
- Harriott, M.; Noverr, M. Importance of Candida-bacterial polymicrobial biofilms in disease. Trends Microbiol. 2011, 19, 557–563. [Google Scholar] [CrossRef]
- Miller, W.D. The Micro-Organisms of the Human Mouth; The S.S. White Dental MFG. Co.: Philadelphia, PA, USA, 1890. [Google Scholar]
- Gibbons, R.; van Houte, J. Selective bacterial adherence to oral epithelial surfaces and its role as an ecological determinant. Infect. Immun. 1971, 3, 567–573. [Google Scholar] [CrossRef]
- JAY, P. The reduction of oral Lactobacillus acidophilus counts by the periodic restriction of carbohydrate. Am. J. Orthod. 1947, 33, 162–184. [Google Scholar] [CrossRef]
- Loesche, W.J. Role of Streptococcus mutans in human dental decay. Microbiol. Rev. 1986, 50, 353–380. [Google Scholar] [CrossRef]
- Zickert, I.; Emilson, C.G. Streptococcus mutans, lactobacilli and dental health in 13–14-year-old Swedish children. Community Dent. Oral Epidemiol. 1982, 10, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Loesche, W.; Bradbury, D. Reduction of dental decay in rampant caries individuals following short-term kanamycin treatment. J. Dent. Res. 1977, 56, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Loesche, W. Chemotherapy of dental plaque infections. Oral Sci. Rev. 1976, 9, 65–107. [Google Scholar] [PubMed]
- Lindquist, B.; Emilson, C. Dental location of Streptococcus mutans and Streptococcus sobrinus in humans harboring both species. Caries Res. 1991, 25, 146–152. [Google Scholar] [CrossRef]
- Marsh, P. Sugar, fluoride, pH and microbial homeostasis in dental plaque. Proc. Finn. Dent. Soc. Suom. Hammaslaak. Toim. 1991, 87, 515–525. [Google Scholar]
- Marsh, P. Microbial ecology of dental plaque and its significance in health and disease. Adv. Dent. Res. 1994, 8, 263–271. [Google Scholar] [CrossRef]
- Aas, J.; Paster, B. Defining the normal bacterial flora of the oral cavity. J. Clin. Microbiol. 2005, 43, 5721–5732. [Google Scholar] [CrossRef]
- Benítez-Páez, A.; Belda-Ferre, P.; Simón-Soro, A.; Mira, A. Microbiota diversity and gene expression dynamics in human oral biofilms. BMC Genom. 2014, 15, 311. [Google Scholar] [CrossRef]
- Takahashi, N.; Nyvad, B. Caries ecology revisited: Microbial dynamics and the caries process. Caries Res. 2008, 42, 409–418. [Google Scholar] [CrossRef]
- Zickert, I.; Emilson, C.G. Correlation of level and duration of Streptococcus mutans infection with incidence of dental caries. Infect. Immun. 1983, 39, 982–985. [Google Scholar] [CrossRef]
- Kressirer, C.A.; Chen, T. Functional profiles of coronal and dentin caries in children. J. Oral Microbiol. 2018, 10, 1495976. [Google Scholar] [CrossRef]
- Krzyściak, W.; Jurczak, A. The virulence of Streptococcus mutans and the ability to form biofilms. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 499–515. [Google Scholar] [CrossRef]
- Loesche, W.; Eklund, S. Longitudinal investigation of bacteriology of human fissure decay: Epidemiological studies in molars shortly after eruption. Infect. Immun. 1984, 46, 765–772. [Google Scholar] [CrossRef]
- Marchant, S.; Brailsford, S.R. The predominant microflora of nursing caries lesions. Caries Res. 2001, 35, 397–406. [Google Scholar] [CrossRef]
- Choi, E.J.; Lee, S.H. Quantitative real-time polymerase chain reaction for Streptococcus mutans and Streptococcus sobrinus in dental plaque samples and its association with early childhood caries. Int. J. Paediatr. Dent. 2009, 19, 141–147. [Google Scholar] [CrossRef]
- Byun, R.; Nadkarni, M.A. Quantitative analysis of diverse Lactobacillus species present in advanced dental caries. J. Clin. Microbiol. 2004, 42, 3128–3136. [Google Scholar] [CrossRef]
- Kianoush, N.; Adler, C. Bacterial profile of dentine caries and the impact of pH on bacterial population diversity. PLoS ONE 2014, 9, e92940. [Google Scholar] [CrossRef]
- Richards, V.P.; Alvarez, A.J.; Luce, A.R.; Bedenbaugh, M.; Mitchell, M.L.; Burne, R.A.; Nascimento, M.M. Microbiomes of Site-Specific Dental Plaques from Children with Different Caries Status. Infect. Immun. 2017, 85, e00106-17. [Google Scholar] [CrossRef] [PubMed]
- Caufield, P.W.; Schön, C.N. Oral Lactobacilli and Dental Caries: A Model for Niche Adaptation in Humans. J. Dent. Res. 2015, 94 (Suppl. 9), 110S–118S. [Google Scholar] [CrossRef] [PubMed]
- Aas, J.A.; Griffen, A.L. Bacteria of dental caries in primary and permanent teeth in children and young adults. J. Clin. Microbiol. 2008, 46, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Brailsford, S.R.; Shah, B. The predominant aciduric microflora of root-caries lesions. J. Dent. Res. 2001, 80, 1828–1833. [Google Scholar] [CrossRef]
- Brailsford, S.R.; Tregaskis, R.B. The predominant Actinomyces spisolated from infected dentin of active root caries lesions. J. Dent. Res. 1999, 78, 1525–1534. [Google Scholar] [CrossRef]
- Piwat, S.; Teanpaisan, R. Lactobacillus species and genotypes associated with dental caries in Thai preschool children. Mol. Oral Microbiol. 2010, 25, 157–164. [Google Scholar] [CrossRef]
- Mantzourani, M.; Fenlon, M. Association between Bifidobacteriaceae and the clinical severity of root caries lesions. Oral Microbiol. Immunol. 2009, 24, 32–37. [Google Scholar] [CrossRef]
- Tanner, A.; Mathney, J. Cultivable anaerobic microbiota of severe early childhood caries. J. Clin. Microbiol. 2011, 49, 1464–1474. [Google Scholar] [CrossRef]
- Nakajo, K.; Takahashi, N. Resistance to acidic environments of caries-associated bacteria: Bifidobacterium dentium and Bifidobacterium longum. Caries Res. 2010, 44, 431–437. [Google Scholar] [CrossRef]
- Kaur, R.; Gilbert, S. Salivary levels of Bifidobacteria in caries-free and caries-active children. Int. J. Paediatr. Dent. 2013, 23, 32–38. [Google Scholar] [CrossRef]
- Qudeimat, M.A.; Alyahya, A. Dental plaque microbiota profiles of children with caries-free and caries-active dentition. J. Dent. 2021, 104, 103539. [Google Scholar] [CrossRef]
- Dame-Teixeira, N.; de Lima, A.K.A. Meta-Analysis Using NGS Data: The Veillonella Species in Dental Caries. Front. Oral Health 2021, 2, 770917. [Google Scholar] [CrossRef]
- Corralo, D.J.; Ev, L.D. Functionally Active Microbiome in Supragingival Biofilms in Health and Caries. Caries Res. 2021, 55, 603–616. [Google Scholar] [CrossRef]
- Zhou, P.; Manoil, D. Veillonellae: Beyond Bridging Species in Oral Biofilm Ecology. Front. Oral Health 2021, 2, 774115. [Google Scholar] [CrossRef]
- Arif, N.; Sheehy, E. Diversity of Veillonella spfrom sound and carious sites in children. J. Dent. Res. 2008, 87, 278–282. [Google Scholar] [CrossRef]
- Falsetta, M.L.; Klein, M.I. Symbiotic relationship between Streptococcus mutans and Candida albicans synergizes virulence of plaque biofilms in vivo. Infect. Immun. 2014, 82, 1968–1981. [Google Scholar] [CrossRef]
- Ellepola, K.; Truong, T.; Liu, Y.; Lin, Q.; Lim, T.K.; Lee, Y.M.; Cao, T.; Koo, H.; Seneviratne, C.J. Multi-omics Analyses Reveal Synergistic Carbohydrate Metabolism in Streptococcus mutans-Candida albicans Mixed-Species Biofilms. Infect. Immun. 2019, 87, e00339-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.; Huang, X. Candida albicans and Early Childhood Caries: A Systematic Review and Meta-Analysis. Caries Res. 2018, 52, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Duangthip, D. Early childhood caries and candida albicans. Evid. Based Dent. 2018, 19, 100–101. [Google Scholar] [PubMed]
- Fechney, J.M.; Browne, G.V. Preliminary study of the oral mycobiome of children with and without dental caries. J. Oral Microbiol. 2019, 11, 1536182. [Google Scholar] [CrossRef]
- O’Connell, L.M.; Santos, R. Site-Specific Profiling of the Dental Mycobiome Reveals Strong Taxonomic Shifts during Progression of Early-Childhood Caries. Appl. Environ. Microbiol. 2020, 86, e02825-19. [Google Scholar] [CrossRef]
- Bustamante, M.; Oomah, B.D. Probiotics as an Adjunct Therapy for the Treatment of Halitosis, Dental Caries and Periodontitis. Probiotics Antimicrob. Proteins 2020, 12, 325–334. [Google Scholar] [CrossRef]
- Zaura, E.; Twetman, S. Critical Appraisal of Oral Pre- and Probiotics for Caries Prevention and Care. Caries Res. 2019, 53, 514–526. [Google Scholar] [CrossRef]
- Havsed, K.; Stensson, M. Bacterial Composition and Metabolomics of Dental Plaque From Adolescents. Front. Cell Infect. Microbiol. 2021, 11, 716493. [Google Scholar] [CrossRef]
- Camelo-Castillo, A.; Benítez-Páez, A. Streptococcus dentisani sp. nov., a novel member of the mitis group. Int. J. Syst. Evol. Microbiol. 2014, 64, 60–65. [Google Scholar] [CrossRef]
- Xu, H.; Tian, J. Oral Microbiome Shifts From Caries-Free to Caries-Affected Status in 3-Year-Old Chinese Children: A Longitudinal Study. Front. Microbiol. 2018, 9, 2009. [Google Scholar] [CrossRef]
- Kim, B.; Han, D. Association of Salivary Microbiota with Dental Caries Incidence with Dentine Involvement after 4 Years. J. Microbiol. Biotechnol. 2018, 28, 454–464. [Google Scholar] [CrossRef] [Green Version]
- Hurley, E.; Barrett, M.P.J. Comparison of the salivary and dentinal microbiome of children with severe-early childhood caries to the salivary microbiome of caries-free children. BMC Oral Health 2019, 19, 13. [Google Scholar] [CrossRef]
- Simón-Soro, A.; Belda-Ferre, P.; Cabrera-Rubio, R.; Alcaraz, L.D.; Mira, A. A tissue-dependent hypothesis of dental caries. Caries Res. 2013, 47, 591–600. [Google Scholar] [CrossRef]
- Jiang, S.; Gao, X.; Jin, L.; Lo, E.C. Salivary Microbiome Diversity in Caries-Free and Caries-Affected Children. Int. J. Mol. Sci. 2016, 17, 1978. [Google Scholar] [CrossRef]
- Kalpana, B.; Prabhu, P.; Bhat, A.H.; Senthilkumar, A.; Arun, R.P.; Asokan, S.; Gunthe, S.S.; Verma, R.S. Bacterial diversity and functional analysis of severe early childhood caries and recurrence in India. Sci. Rep. 2020, 10, 21248. [Google Scholar] [CrossRef]
- Pang, L.; Wang, Y. Metagenomic Analysis of Dental Plaque on Pit and Fissure Sites With and Without Caries Among Adolescents. Front. Cell Infect. Microbiol. 2021, 11, 740981. [Google Scholar] [CrossRef]
- Jiang, Q.; Liu, J. The Oral Microbiome in the Elderly With Dental Caries and Health. Front. Cell Infect. Microbiol. 2018, 8, 442. [Google Scholar] [CrossRef]
- Bhaumik, D.; Manikandan, D. Cariogenic and oral health taxa in the oral cavity among children and adults: A scoping review. Arch. Oral Biol. 2021, 129, 105204. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; He, J. Oral cavity contains distinct niches with dynamic microbial communities. Environ. Microbiol. 2015, 17, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Simón-Soro, A.; Guillen-Navarro, M. Metatranscriptomics reveals overall active bacterial composition in caries lesions. J. Oral Microbiol. 2014, 6, 25443. [Google Scholar] [CrossRef] [PubMed]
- Mei, M.L.; Yan, Z. Effect of silver diamine fluoride on plaque microbiome in children. J. Dent. 2020, 102, 103479. [Google Scholar] [CrossRef] [PubMed]
- Mitwalli, H.; Mourao, M.D.A. Effect of Silver Diamine Fluoride Treatment on Microbial Profiles of Plaque Biofilms from Root/Cervical Caries Lesions. Caries Res. 2019, 53, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Da Costa Rosa, T.; de Almeida Neves, A. The bacterial microbiome and metabolome in caries progression and arrest. J. Oral Microbiol. 2021, 13, 1886748. [Google Scholar] [CrossRef]
- Do, T.; Damé-Teixeira, N. Root Surface Biofilms and Caries. Monogr. Oral Sci. 2017, 26, 26–34. [Google Scholar]
- Syed, S.A.; Loesche, W.J. Predominant cultivable flora isolated from human root surface caries plaque. Infect. Immun. 1975, 11, 727–731. [Google Scholar] [CrossRef]
- Akenaka, S.; Edanami, N.; Komatsu, Y.; Nagata, R.; Naksagoon, T.; Sotozono, M.; Ida, T.; Noiri, Y. Periodontal Pathogens Inhabit Root Caries Lesions Extending beyond the Gingival Margin: A Next-Generation Sequencing Analysis. Microorganisms 2021, 9, 2349. [Google Scholar] [CrossRef]
- Baker, J.L.; Morton, J.T. Deep metagenomics examines the oral microbiome during dental caries, revealing novel taxa and co-occurrences with host molecules. Genome Res. 2021, 31, 64–74. [Google Scholar] [CrossRef]
- Chawhuaveang, D.D.; Yu, O.Y. Acquired salivary pellicle and oral diseases: A literature review. J. Dent. Sci. 2021, 16, 523–529. [Google Scholar] [CrossRef]
- Cugini, C.; Ramasubbu, N. Dysbiosis From a Microbial and Host Perspective Relative to Oral Health and Disease. Front. Microbiol. 2021, 12, 617485. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Microbial Species | Dentition | Location | Infected Tissue | Role in Caries | References |
|---|---|---|---|---|---|
| Bacteria | |||||
| S. mutans | Primary | Coronal | Enamel and dentine | Biofilm formation Caries initiation and progression | [62,63,64,65,66] |
| S. sobrinus | Primary | Coronal | N/A | N/A | [64,67,68,69,70,71] |
| L. salivarius | Primary and permanent | Coronal and root | Dentine | Caries progression | |
| L. gasseri | Primary and permanent | Coronal and root | Dentine | Caries progression | |
| L. fermentum | Primary and permanent | Coronal and root | Dentine | Caries progression | |
| L. casei | Primary and permanent | Coronal and root | Dentine | Caries progression | |
| A. israelii | Permanent | Root | N/A | Caries initiation and progression | [5,72,73] |
| A. gerencseriae | Primary and permanent | Coronal and root | N/A | Caries initiation and progression | |
| A. naeslundii | Permanent | Root | N/A | Caries initiation and progression | |
| S. wiggsiae | Primary | Coronal | Dentine | Caries progression | [67,69,74,75,76] |
| P. denticolens | Primary and permanent | Coronal and root | Dentine | Caries progression | [69,75] |
| B. dentium | Permanent | Coronal and root | Dentine | Caries progression | [74,75,76,77,78] |
| B. longum | Permanent | Coronal and root | Dentine | Caries progression | |
| B. breve | Permanent | Root | Dentine | Caries progression | |
| L. shahii | Primary | Coronal | N/A | Caries progression | [71,77,78,79,80,81] |
| L. HOT 498 | Primary | Coronal | N/A | Caries progression | |
| P. melaninogenica | Primary | Coronal | Enamel and dentine | Caries progression | [77,78,79] |
| V. dispar | Primary | Coronal | Enamel and dentine | Biofilm formation Caries initiation and progression | [71,79,82,83] |
| V. parvula | Primary and permanent | Coronal and root | Enamel and dentine | Caries initiation and progression | |
| V. denticariosi | Primary | Coronal | Enamel and dentine | Caries initiation and progression | |
| Fungi | |||||
| C. albicans | Primary | Coronal | Enamel | Symbiotic with S. mutans Caries initiation | [79,84,85,86,87,88,89] |
| C. dubliniensis | Primary | Coronal | N/A | Caries progression | |
| N. oryzae | Primary | Coronal | N/A | N/A | [89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.S.; Chu, C.-H.; Yu, O.Y. Oral Microbiome and Dental Caries Development. Dent. J. 2022, 10, 184. https://doi.org/10.3390/dj10100184
Zhang JS, Chu C-H, Yu OY. Oral Microbiome and Dental Caries Development. Dentistry Journal. 2022; 10(10):184. https://doi.org/10.3390/dj10100184
Chicago/Turabian StyleZhang, Josie Shizhen, Chun-Hung Chu, and Ollie Yiru Yu. 2022. "Oral Microbiome and Dental Caries Development" Dentistry Journal 10, no. 10: 184. https://doi.org/10.3390/dj10100184