
Effects of Boron-Containing Compounds on Liposoluble Hormone Functions
 , , ,
, , ,
Abstract
:1. Introduction
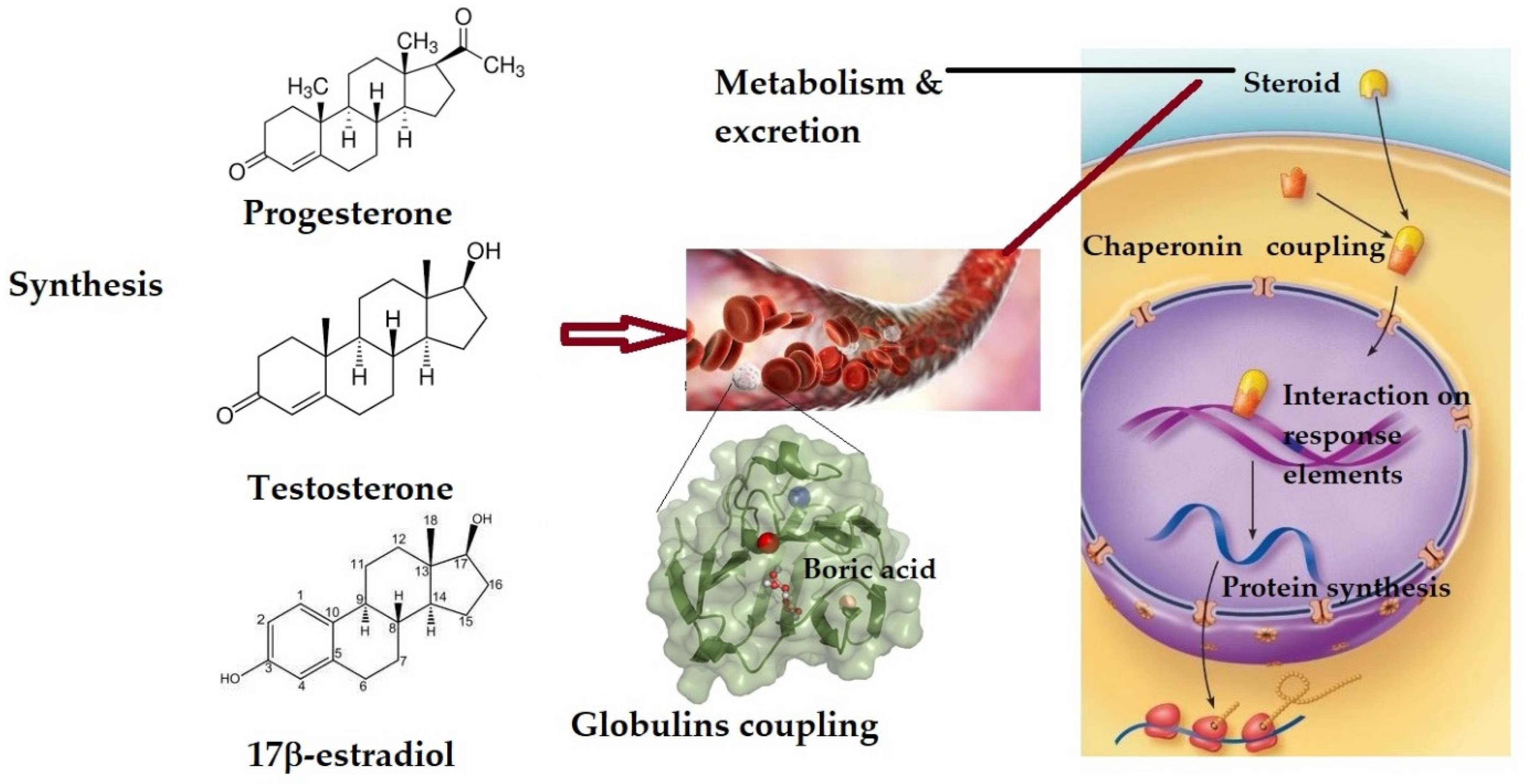
2. The Effect on Sex Hormones and Their Receptors
2.1. Changes in Sex Hormones’ Plasma Level
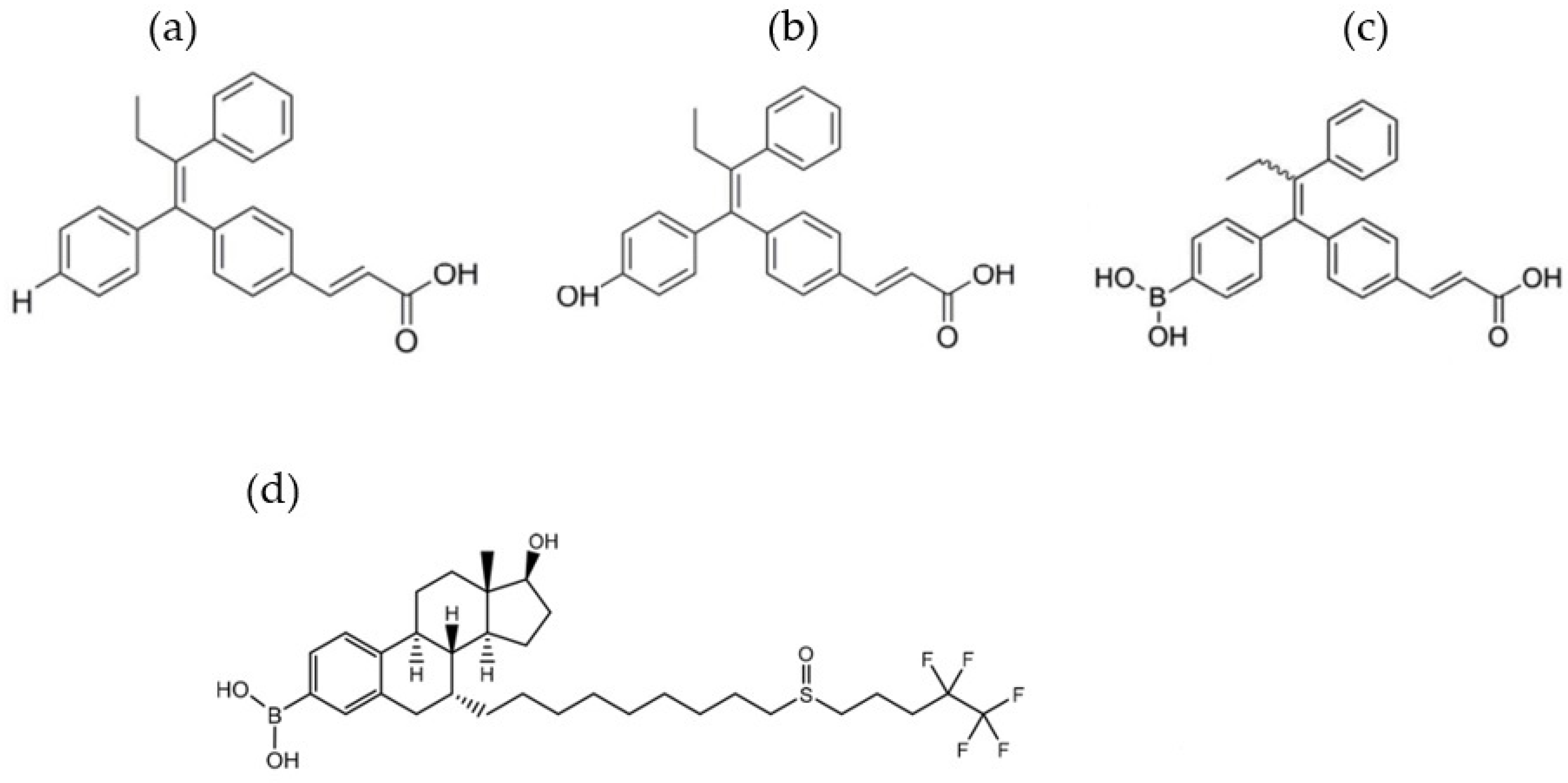
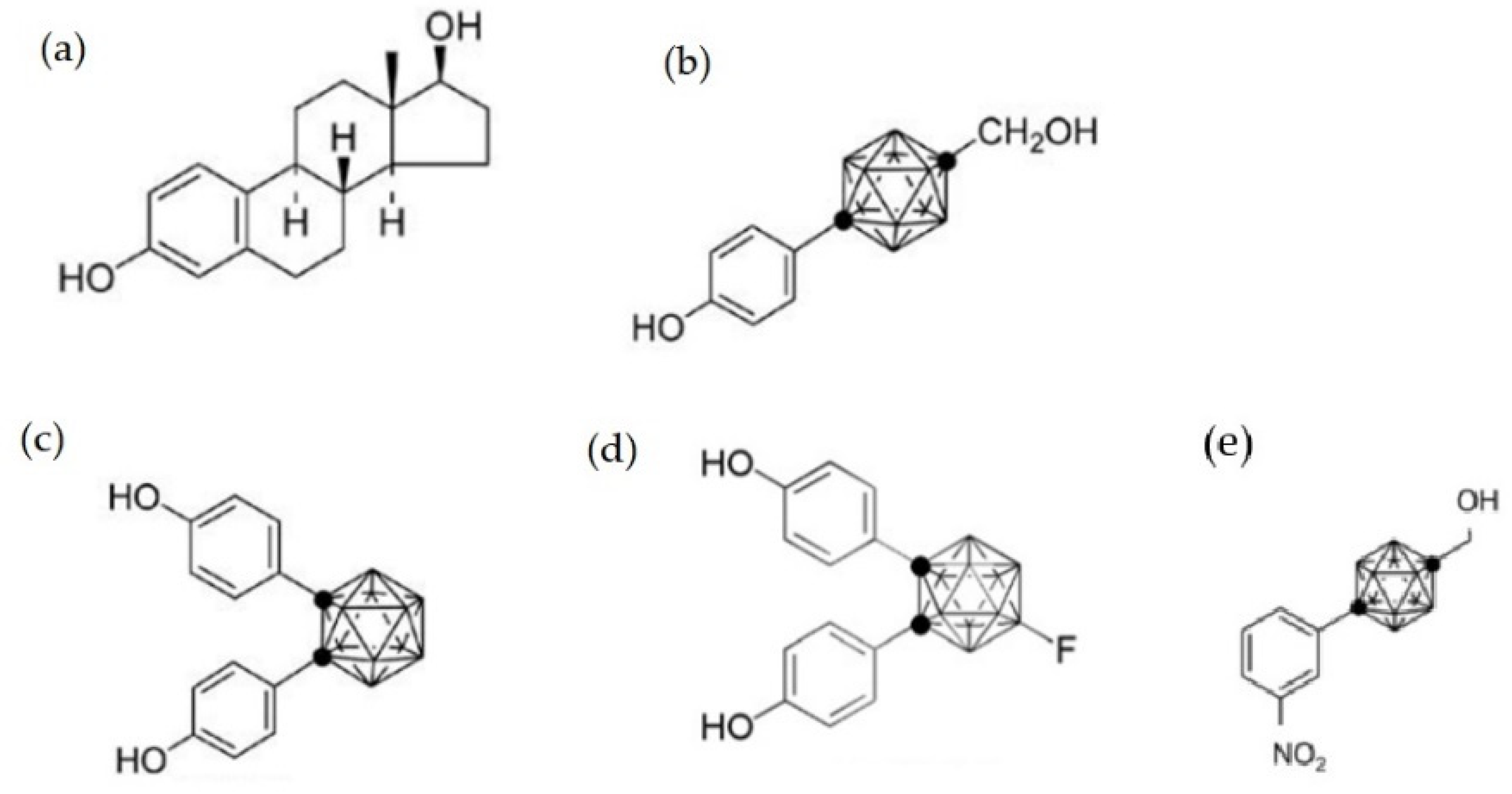
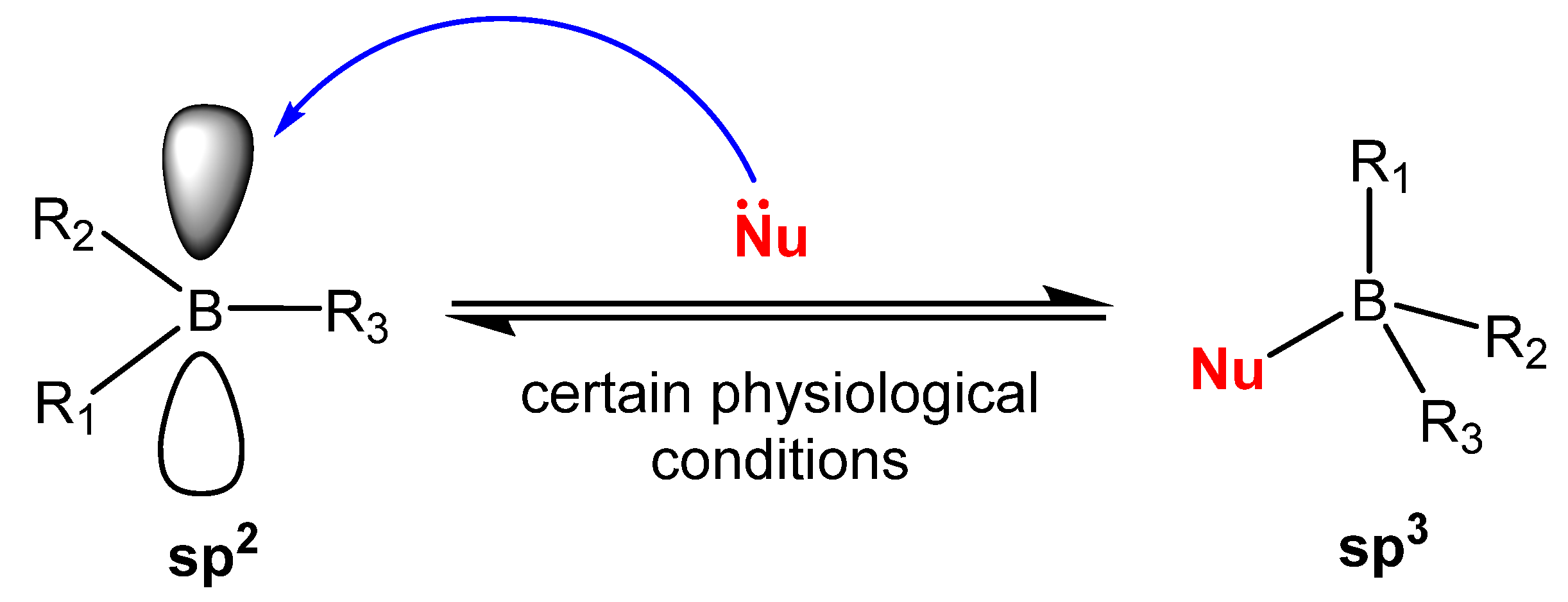
2.2. The Effect on Sex Hormone Receptors
2.3. The Effect on Sex Hormone-Dependent Human Metabolism
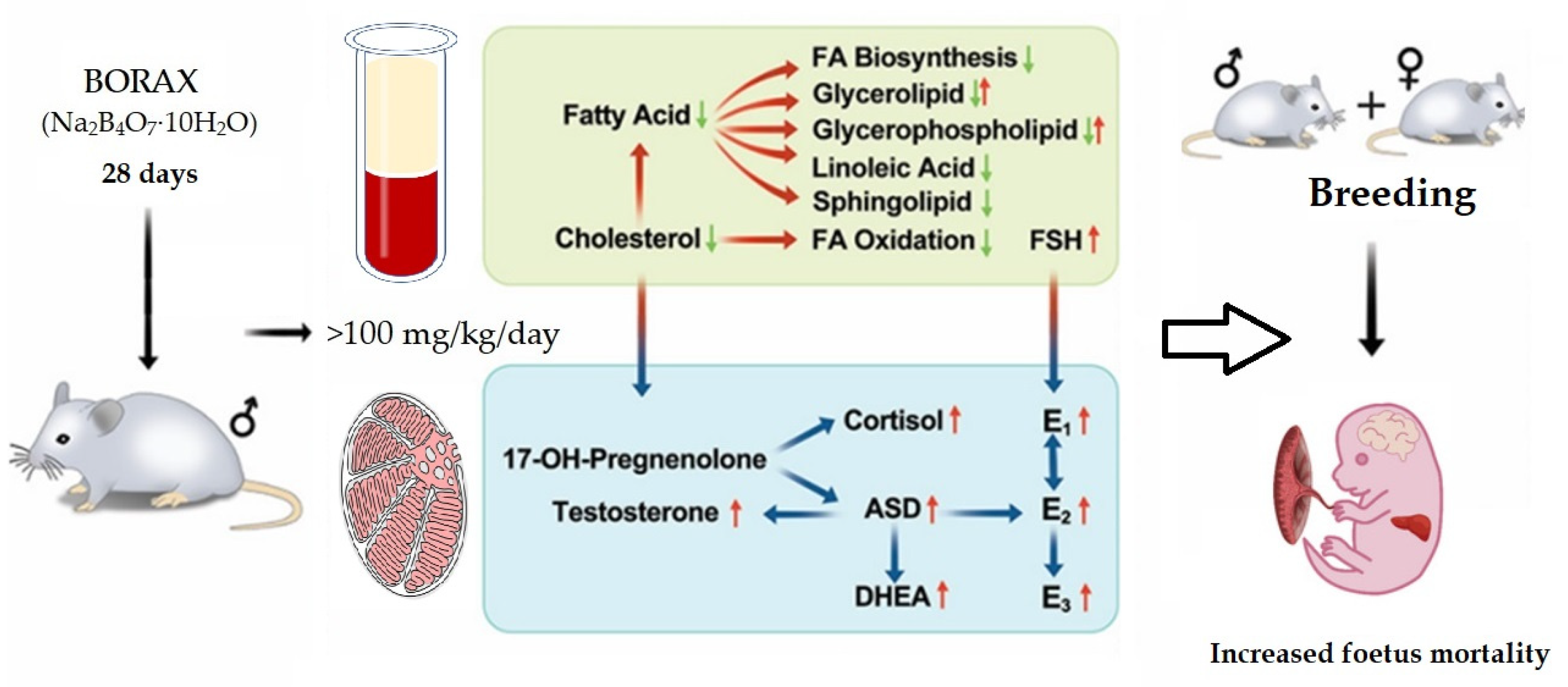
2.4. Regarding Fertility
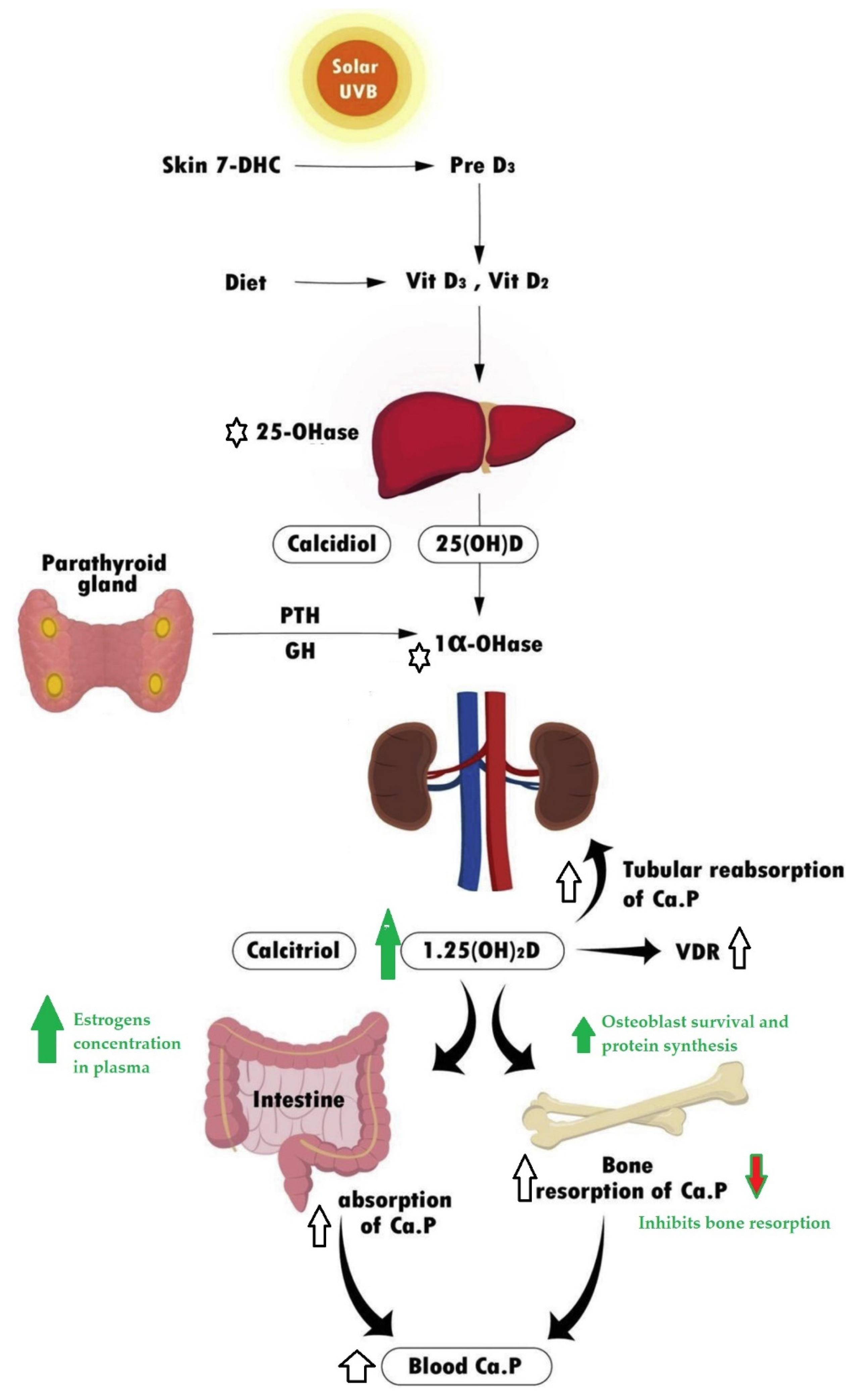
3. The Effect in Vitamin D Plasma Levels and Actions
4. Effects in Thyroid Hormones
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, J.; Wang, M.; Pallarés, N.; Ferrer, E.; Berrada, H.; Barba, F.J. Sterols and fat-soluble vitamins. In Food Lipids; Academic Press: Cambridge, MA, USA, 2022; pp. 323–348. [Google Scholar]
- Cole, T.J.; Short, K.L.; Hooper, S.B. The science of steroids. In Proceedings of the Seminars in Fetal and Neonatal Medicine; Elsevier: Amsterdam, The Netherlands, 2019; Volume 24, pp. 170–175. [Google Scholar]
- Rafeeq, H.; Ahmad, S.; Tareen, M.B.K.; Shahzad, K.A.; Bashir, A.; Jabeen, R.; Shehzadi, I. Biochemistry of Fat Soluble Vitamins, Sources, Biochemical Functions and Toxicity. Haya Saudi J. Life Sci. 2020, 5, 188–196. [Google Scholar] [CrossRef]
- Imig, J.D. Eicosanoid blood vessel regulation in physiological and pathological states. Clin. Sci. 2020, 134, 2707–2727. [Google Scholar] [CrossRef] [PubMed]
- Soriano-Ursúa, M.A.; Farfán-García, E.D.; Geninatti-Crich, S. Turning Fear of Boron Toxicity into Boron-containing Drug Design. Curr. Med. Chem. 2019, 26, 5005–5018. [Google Scholar] [CrossRef] [PubMed]
- Hosmane, N.S. Boron Science: New Technologies and Applications; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Topnikova, A.P.; Belokoneva, E.L. The structure and classification of complex borates. Russ. Chem. Rev. 2019, 88, 204. [Google Scholar] [CrossRef]
- Muetterties, E. Boron Hydride Chemistry; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 032314649X. [Google Scholar]
- Hey-Hawkins, E.; Teixidor, C.V. Boron-Based Compounds: Potential and Emerging Applications in Medicine; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Grimes, R.N. Carboranes; Academic Press: Cambridge, MA, USA, 2016; ISBN 0128019050. [Google Scholar]
- Mogoşanu, G.D.; Biţă, A.; Bejenaru, L.E.; Bejenaru, C.; Croitoru, O.; Rău, G.; Rogoveanu, O.-C.; Florescu, D.N.; Neamţu, J.; Scorei, I.D.; et al. Calcium Fructoborate for Bone and Cardiovascular Health. Biol. Trace Elem. Res. 2016, 172, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Gizer, M.; Köse, S.; Karaosmanoglu, B.; Taskiran, E.Z.; Berkkan, A.; Timuçin, M.; Korkusuz, F.; Korkusuz, P. The effect of boron-containing nano-hydroxyapatite on bone cells. Biol. Trace Elem. Res. 2020, 193, 364–376. [Google Scholar] [CrossRef]
- Sizmaz, O.; Koksal, B.; Tekeli, A.; Yildiz, G. Effects of boron supplementation alone or in combination with different vitamin D-3 levels on laying performance, eggshell quality, and mineral content and fatty acid composition of egg yolk in laying hens. J. Anim. Feed Sci. 2021, 30, 288–294. [Google Scholar] [CrossRef]
- Naghii, M.R.; Mofid, M.; Asgari, A.R.; Hedayati, M.; Daneshpour, M.S. Comparative effects of daily and weekly boron supplementation on plasma steroid hormones and proinflammatory cytokines. J. Trace Elem. Med. Biol. 2011, 25, 54–58. [Google Scholar] [CrossRef]
- Ri, C.-C.; Mf, C.-R.; IR, S.; MA, S.-U. Boron-Containing Compounds for Prevention, Diagnosis, and Treatment of Human Metabolic Disorders. Biol. Trace Elem. Res. 2022, online ahead of print. [Google Scholar] [CrossRef]
- Kan, F.; Kucukkurt, I. Investigation of the effect of boron on thyroid functions and biochemical parameters in hypothyroid induced-rats. J. Biochem. Mol. Toxicol. 2022, 36, e23186. [Google Scholar] [CrossRef]
- Rondanelli, M.; Faliva, M.A.; Peroni, G.; Infantino, V.; Gasparri, C.; Iannello, G.; Perna, S.; Riva, A.; Petrangolini, G.; Tartara, A. Pivotal role of boron supplementation on bone health: A narrative review. J. Trace Elem. Med. Biol. 2020, 62, 126577. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Kong, Z.; Duan, L.; Deng, F.; Chen, Y.; Quan, S.; Liu, X.; Cha, Y.; Gong, Y.; Wang, C. Reproductive toxicity and metabolic perturbations in male rats exposed to boron. Sci. Total Environ. 2021, 785, 147370. [Google Scholar] [CrossRef]
- Pizzorno, L. Nothing boring about boron. Integr. Med. 2015, 14, 35–48. [Google Scholar]
- Ghanizadeh, G.; Babaei, M.; Naghii, M.R.; Mofid, M.; Torkaman, G.; Hedayati, M. The effect of supplementation of calcium, vitamin D, boron, and increased fluoride intake on bone mechanical properties and metabolic hormones in rat. Toxicol. Ind. Health 2014, 30, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Samman, S.; Naghii, M.R.; Lyons Wall, P.M.; Verus, A.P. The nutritional and metabolic effects of Boron in humans and animals. Biol. Trace Elem. Res. 1998, 66, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Naghii, M.R.; Samman, S. The effect of boron supplementation on the distribution of boron in selected tissues and on testosterone synthesis in rats. J. Nutr. Biochem. 1996, 7, 507–512. [Google Scholar] [CrossRef]
- Naghii, M.R.; Samman, S. The effect of boron supplementation on its urinary excretion and selected cardiovascular risk factors in healthy male subjects. Biol. Trace Elem. Res. 1997, 56, 273–286. [Google Scholar] [CrossRef]
- Lee, I.P.; Sherins, R.J.; Dixon, R.L. Evidence for induction of germinal aplasia in male rats by environmental exposure to boron. Toxicol. Appl. Pharmacol. 1978, 45, 577–590. [Google Scholar] [CrossRef]
- Nielsen, F.H.; Gallagher, S.K.; Johnson, L.K.; Nielsen, E.J. Boron enhaces and mimics some effects of estrogen therapy in psotmenopausal women. J. Trace Elem. Exp. Med. 1992, 5, 237–246. [Google Scholar]
- Bello, M.; Guadarrama-García, C.; Velasco-Silveyra, L.M.; Farfán-García, E.D.; Soriano-Ursúa, M.A. Several effects of boron are induced by uncoupling steroid hormones from their transporters in blood. Med. Hypotheses 2018, 118, 78–83. [Google Scholar] [CrossRef]
- Avvakumov, G.V.; Grishkovskaya, I.; Muller, Y.A.; Hammond, G.L. Resolution of the human sex hormone-binding globulin dimer interface and evidence for two steroidbinding sites per homodimer. J. Biol. Chem. 2001, 276, 34453–34457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, G.L. Plasma steroid-binding proteins: Primary gatekeepers of steroid hormone action. J. Endocrinol. 2016, 230, R13–R25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breuner, C.W.; Orchinik, M. Plasma binding proteins as mediators of corticosteroid action in vertebrates. J. Endocrinol. 2002, 175, 99–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artem, C.; Zheng, S.; Magid, F.; Hammond, G.L. Successful in silico discovery of novelnonsteroidal ligands for human sex hormone binding globulin. J. Med. Chem. 2005, 48, 3203–3213. [Google Scholar]
- Simó, R.; Sáez-López, C.; Barbosa-Desongles, A.; Hernández, C.; Selva, D.M. Novel insights in SHBG regulation and clinical implications. Trends Endocrinol. Metab. 2015, 26, 376–383. [Google Scholar] [CrossRef]
- Gardill, B.R.; Vogl, M.R.; Lin, H.Y.; Hammond, G.L.; Muller, Y.A. Corticosteroid-binding globulin: Structure-function implications from species differences. PLoS ONE 2012, 7, e52759. [Google Scholar] [CrossRef] [Green Version]
- Esther, M.Y.; Subramaniyan, V.; Kumar, A.P.; Subramanian, M.; Palani, M. Molecular docking, ADMET analysis and dynamics approach to potent natural inhibitors against sex hormone binding globulin in male infertility. Pharmacogn. J. 2017, 9, s35–s43. [Google Scholar] [CrossRef] [Green Version]
- Başaran, N.; Duydu, Y.; Bolt, H.M. Reproductive toxicity in boron exposed workers in Bandirma, Turkey. J. Trace Elem. Med. Biol. 2012, 26, 165–167. [Google Scholar] [CrossRef]
- Green, N.R.; Ferrando, A.A. Plasma boron and the effects of boron supplementation in males. Environ. Health Perspect. 1994, 102, 73–77. [Google Scholar]
- Leifke, E.; Gorenoi, V.; Wichers, C.; Von Zur Mühlen, A.; Von Büren, E.; Brabant, G. Age-related changes of serum sex hormones, insulin-like growth factor-1 and sex-hormone binding globulin levels in men: Cross-sectional data from a healthy male cohort. Clin. Endocrinol. (Oxf.) 2000, 53, 689–695. [Google Scholar] [CrossRef]
- Hunt, C.D. Dietary Boron: An Overview of the Evidence for Its Role in Immune Function. J. Trace Elem. Exp. Med. 2003, 16, 291–306. [Google Scholar] [CrossRef]
- Beattie, J.H.; Peace, H.S. The influence of a low-boron diet and boron supplementation on bone, major mineral and sex steroid metabolism in postmenopausal women. Br. J. Nutr. 1993, 69, 871–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyacioglu, O.; Orenay-Boyacioglu, S.; Yildirim, H.; Korkmaz, M. Boron intake, osteocalcin polymorphism and serum level in postmenopausal osteoporosis. J. Trace Elem. Med. Biol. 2018, 48, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Orenay-Boyacioglu, S.; Korkmaz, M.; Kahraman, E.; Yildirim, H.; Bora, S.; Ataman, O.Y. Biological effects of tolerable level chronic boron intake on transcription factors. J. Trace Elem. Med. Biol. 2017, 39, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, T.A.; Flowers, W.L.; Spears, J.W.; Nielsent, F.H. Long-term effects of boron supplementation on reproductive characteristics and bone mechanical properties in gilts. J. Anim. Sci. 2002, 80, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, T.A.; Spears, J.W. Effect of boron supplementation of pig diets on the production of tumor necrosis factor- α and interferon- γ 1, 2. Am. Soc. Anim. Sci. 2003, 81, 2552–2561. [Google Scholar] [CrossRef]
- Hunt, C.D.; Herbel, J.L.; Nielsen, F.H. Metabolic responses of postmenopausal women to supplemental dietary boron and aluminum during usual and low magnesium intake: Boron, calcium, and magnesium absorption and retention and blood mineral concentrations. Am. J. Clin. Nutr. 1997, 65, 803–813. [Google Scholar] [CrossRef] [Green Version]
- Khaliq, H.; Juming, Z.; Ke-Mei, P. The Physiological Role of Boron on Health. Biol. Trace Elem. Res. 2018, 186, 31–51. [Google Scholar] [CrossRef]
- Liu, J.; Zheng, S.; Akerstrom, V.L.; Yuan, C.; Ma, Y.; Zhong, Q.; Zhang, C.; Zhang, Q.; Guo, S.; Ma, P. Fulvestrant-3 boronic acid (ZB716): An orally bioavailable selective estrogen receptor downregulator (SERD). J. Med. Chem. 2016, 59, 8134–8140. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zheng, S.; Guo, S.; Zhang, C.; Zhong, Q.; Zhang, Q.; Ma, P.; Skripnikova, E.V.; Bratton, M.R.; Wiese, T.E. Rational design of a boron-modified triphenylethylene (GLL398) as an oral selective estrogen receptor downregulator. ACS Med. Chem. Lett. 2017, 8, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Raghava, N.; Das, B.C.; Ray, S.K. Neuroprotective effects of estrogen in CNS injuries: Insights from animal models. Neurosci. Neuroeconomics 2017, 6, 15–29. [Google Scholar] [CrossRef] [Green Version]
- Fink, K.; Uchman, M. Boron cluster compounds as new chemical leads for antimicrobial therapy. Coord. Chem. Rev. 2021, 431, 213684. [Google Scholar] [CrossRef]
- Scholz, M.; Hey-Hawkins, E. Carbaboranes as pharmacophores: Properties, synthesis, and application strategies. Chem. Rev. 2011, 111, 7035–7062. [Google Scholar] [CrossRef] [PubMed]
- Avdeeva, V.V.; Garaev, T.M.; Malinina, E.A.; Zhizhin, K.Y.; Kuznetsov, N.T. Physiologically Active Compounds Based on Membranotropic Cage Carriers–Derivatives of Adamantane and Polyhedral Boron Clusters. Russ. J. Inorg. Chem. 2022, 67, 28–47. [Google Scholar] [CrossRef]
- Soriano-Ursúa, M.A.; Das, B.C.; Trujillo-Ferrara, J.G. Boron-containing compounds: Chemico-biological properties and expanding medicinal potential in prevention, diagnosis and therapy. Expert Opin. Ther. Pat. 2014, 24, 485–500. [Google Scholar] [CrossRef]
- Thirumamagal, B.T.S.; Zhao, X.B.; Bandyopadhyaya, A.K.; Narayanasamy, S.; Johnsamuel, J.; Tiwari, R.; Golightly, D.W.; Patel, V.; Jehning, B.T.; Backer, M.V. Receptor-targeted liposomal delivery of boron-containing cholesterol mimics for boron neutron capture therapy (BNCT). Bioconjug. Chem. 2006, 17, 1141–1150. [Google Scholar] [CrossRef]
- Messner, K.; Vuong, B.; Tranmer, G.K. The Boron Advantage: The Evolution and Diversification of Boron’s Applications in Medicinal Chemistry. Pharmaceuticals 2022, 15, 264. [Google Scholar] [CrossRef]
- Sedlak, D.; Wilson, T.A.; Tjarks, W.; Radomska, H.S.; Wang, H.; Kolla, J.N.; Lesnikowski, Z.J.; Spicakova, A.; Ali, T.; Ishita, K. Structure–Activity Relationship of para-Carborane Selective Estrogen Receptor β Agonists. J. Med. Chem. 2021, 64, 9330–9353. [Google Scholar] [CrossRef]
- Machuki, J.O.; Zhang, H.Y.; Harding, S.E.; Sun, H. Molecular pathways of oestrogen receptors and β-adrenergic receptors in cardiac cells: Recognition of their similarities, interactions and therapeutic value. Acta Physiol. 2018, 222, e12978. [Google Scholar] [CrossRef]
- Ohta, K.; Ogawa, T.; Oda, A.; Kaise, A.; Endo, Y. Design and synthesis of carborane-containing estrogen receptor-beta (ERβ)-selective ligands. Bioorganic Med. Chem. Lett. 2015, 25, 4174–4178. [Google Scholar] [CrossRef]
- Endo, Y.; Iijima, T.; Yamakoshi, Y.; Fukasawa, H.; Miyaura, C.; Inada, M.; Kubo, A.; Itai, A. Potent estrogen agonists based on carborane as a hydrophobic skeletal structure: A new medicinal application of boron clusters. Chem. Biol. 2001, 8, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, F.H.; Hunt, C.D.; Mullen, L.M.; Hunt, J.R. Effect of dietary boron on mineral, estrogen, and testosterone metabolism in postmenopausal women 1. FASEB J. 1987, 1, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Hakki, S.S.; Bozkurt, B.S.; Hakki, E.E. Boron regulates mineralized tissue-associated proteins in osteoblasts (MC3T3-E1). J. Trace Elem. Med. Biol. 2010, 24, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Hirata, M.; Inada, M.; Matsumoto, C.; Takita, M.; Ogawa, T.; Endo, Y.; Miyaura, C. A novel carborane analog, BE360, with a carbon-containing polyhedral boron-cluster is a new selective estrogen receptor modulator for bone. Biochem. Biophys. Res. Commun. 2009, 380, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Ogawa, T.; Kaise, A.; Endo, Y. Enhanced estrogen receptor beta (ERβ) selectivity of fluorinated carborane-containing ER modulators. Bioorg. Med. Chem. Lett. 2013, 23, 6555–6558. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Ogawa, T.; Endo, Y. Estrogenic activity of B-fluorinated o-carborane-1,2-bisphenol synthesized via SNAr reaction. Bioorg. Med. Chem. Lett. 2012, 22, 4728–4730. [Google Scholar] [CrossRef]
- Paterni, I.; Granchi, C.; Katzenellenbogen, J.A.; Minutolo, F. Estrogen receptors alpha (ERα) and beta (ERβ): Subtype-selective ligands and clinical potential. Steroids 2014, 90, 13–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.; Gustafsson, J.-Å. The different roles of ER subtypes in cancer biology and therapy. Nat. Rev. Cancer 2011, 11, 597–608. [Google Scholar] [CrossRef]
- Dall, G.V.; Hawthorne, S.; Seyed-Razavi, Y.; Vieusseux, J.; Wu, W.; Gustafsson, J.-A.; Byrne, D.; Murphy, L.; Risbridger, G.P.; Britt, K.L. Estrogen receptor subtypes dictate the proliferative nature of the mammary gland. J. Endocrinol. 2018, 237, 323–336. [Google Scholar] [CrossRef] [Green Version]
- Murphy, N.; McCarthy, E.; Dwyer, R.; Farràs, P. Boron clusters as breast cancer therapeutics. J. Inorg. Biochem. 2021, 218, 111412. [Google Scholar] [CrossRef]
- Watanabe, K.; Hirata, M.; Tominari, T.; Matsumoto, C.; Endo, Y.; Murphy, G.; Nagase, H.; Inada, M.; Miyaura, C. BA321, a novel carborane analog that binds to androgen and estrogen receptors, acts as a new selective androgen receptor modulator of bone in male mice. Biochem. Biophys. Res. Commun. 2016, 478, 279–285. [Google Scholar] [CrossRef]
- Romero-Aguilar, K.S.; Arciniega-Martínez, I.M.; Farfán-García, E.D.; Campos-Rodríguez, R.; Reséndiz-Albor, A.A.; Soriano-Ursúa, M.A. Effects of boron-containing compounds on immune responses: Review and patenting trends. Expert Opin. Ther. Pat. 2019, 29, 339–351. [Google Scholar] [CrossRef]
- Jin, E.; Pei, Y.; Liu, T.; Ren, M.; Hu, Q.; Gu, Y.; Li, S. Effects of boron on the proliferation, apoptosis and immune function of splenic lymphocytes through ERα and ERβ. Food Agric. Immunol. 2019, 30, 743–761. [Google Scholar] [CrossRef] [Green Version]
- Mori, S.; Tsuemoto, N.; Kasagawa, T.; Nakano, E.; Fujii, S.; Kagechika, H. Development of Boron-Cluster-Based Progesterone Receptor Antagonists Bearing a Pentafluorosulfanyl (SF5) Group. Chem. Pharm. Bull. 2019, 67, 1278–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, S.; Takagaki, R.; Fujii, S.; Urushibara, K.; Tanatani, A.; Kagechika, H. Novel Non-steroidal Progesterone Receptor Ligands Based on m-Carborane Containing a Secondary Alcohol: Effect of Chirality on Ligand Activity. Chem. Pharm. Bull. 2017, 65, 1051–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, S.; Nakano, E.; Yanagida, N.; Mori, S.; Masuno, H.; Kagechika, H. Development of p-carborane-based nonsteroidal progesterone receptor antagonists. Bioorg. Med. Chem. 2014, 22, 5329–5337. [Google Scholar] [CrossRef] [PubMed]
- Zargham, E.O.; Mason, C.A.; Lee, M.W., Jr. The use of carboranes in cancer drug development. Int. J. Cancer Clin. Res 2019, 6, 110–113. [Google Scholar]
- Goto, T.; Ohta, K.; Fujii, S.; Ohta, S.; Endo, Y. Design and synthesis of androgen receptor full antagonists bearing ap-carborane cage: Promising ligands for anti-androgen withdrawal syndrome. J. Med. Chem. 2010, 53, 4917–4926. [Google Scholar] [CrossRef]
- Azad, N.; Sakla, N.; Bahn, G. The effect of testosterone replacement therapy on glycemic control in hypogonadal men with type 2 diabetes mellitus. J. Clin. Diabetes 2018, 1, 1–5. [Google Scholar]
- Gambineri, A.; Pelusi, C. Sex hormones, obesity and type 2 diabetes: Is there a link? Endocr. Connect. 2019, 8, R1–R9. [Google Scholar] [CrossRef]
- Schiffer, L.; Kempegowda, P.; Arlt, W.; O’Reilly, M.W. Mechanisms in endocrinology: The sexually dimorphic role of androgens in human metabolic disease. Eur. J. Endocrinol. 2017, 177, R125–R143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, G.F.S.; Denny, W.A.; Dos Santos, J.L. Boron in drug design: Recent advances in the development of new therapeutic agents. Eur. J. Med. Chem. 2019, 179, 791–804. [Google Scholar] [CrossRef]
- Morgentaler, A. Testosterone for Life: Recharge Your Vitality, Sex Drive, Muscle Mass, and Overall Health; McGraw-Hill: New York, NY, USA, 2009; ISBN 9780071642514. [Google Scholar]
- Nielsen, F.H. Is boron nutritionally relevant? Nutr. Rev. 2008, 66, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Farfán-García, E.D.; Castillo-Mendieta, N.T.; Ciprés-Flores, F.J.; Padilla-Martínez, I.I.; Trujillo-Ferrara, J.G.; Soriano-Ursúa, M.A. Current data regarding the structure-toxicity relationship of boron-containing compounds. Toxicol. Lett. 2016, 258, 115–125. [Google Scholar] [CrossRef]
- Moreira, W.; Aziz, D.B.; Dick, T. Boromycin kills mycobacterial persisters without detectable resistance. Front Microbiol. 2016, 7, 199. [Google Scholar] [CrossRef] [Green Version]
- Rogoveanu, O.C.; Mogoşanu, G.D.; Bejenaru, C.; Bejenaru, L.E.; Croitoru, O.; Neamţu, J.; Pietrzkowski, Z.; Reyes-Izquierdo, T.; Biţă, A.; Scorei, I.D.; et al. Effects of calcium fructoborate on levels of C-reactive protein, total cholesterol, low-density lipoprotein, triglycerides, IL-1β, IL-6, and MCP-1: A double-blind, placebo-controlled clinical study. Biol. Trace Elem. Res. 2015, 163, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Bita, A.; Mogosanu, G.D.; Bejenaru, L.E.; Oancea, C.N.; Bejenaru, C.; Croitoru, O.; Rau, G.; Neamtu, J.; Scorei, I.D.; Scorei, I.R.; et al. Simultaneous quantitation of boric acid and calcium fructoborate in dietary supplements by HPTLC–densitometry. Anal. Chem. Res. 2017, 33, 743–746. [Google Scholar]
- Naghii, M.R.; Mofid, M. Elevation of biosynthesis of endogenous 17-B oestradiol by boron supplementation: One possible role of dietary boron consumption in humans. J. Nutr. Environ. Med. 2008, 17, 127–135. [Google Scholar] [CrossRef]
- Naghii, M.R.; Samman, S. Role of boron in nutrition and metabolism. Prog. Fd. Nutr. Sci. 1993, 17, 331–349. [Google Scholar]
- Brewster, J.H.; Negishi, E. Brown: Passes through the mountains. Science 1980, 207, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Granner, D.K. Hormones of the Gonads, 21st ed.; Murray, R.K., Granner, D.K., Mayes, P.A., Rodwell, V.W., Eds.; Appleton & Lange: New York, NY, USA, 1988. [Google Scholar]
- Beattie, J.H.; Weersink, E. Borate and molybdate inhibitory of catechol estrogen and pyrocatechol methylation by catechol-o-methyltransferase. J. Inorg. Biochem. 1992, 46, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Naghii, M.R.; Samman, S. The effect of boron on plasma testosterone and plasma lipids in rats. Nutr. Res. 1997, 17, 523–532. [Google Scholar] [CrossRef]
- Nielsen, F.H.; Meacham, S.L. Growing Evidence for Human Health Benefits of Boron. J. Evid. Based. Complementary Altern. Med. 2011, 16, 169–180. [Google Scholar] [CrossRef]
- Kilic, A.; Savci, A.; Alan, Y.; Beyazsakal, L. The synthesis of novel boronate esters and N-Heterocyclic carbene (NHC)-stabilized boronate esters: Spectroscopy, antimicrobial and antioxidant studies. J. Organomet. Chem. 2020, 917, 121268. [Google Scholar] [CrossRef]
- Barrón-González, M.; Montes-Aparicio, A.V.; Cuevas-Galindo, M.E.; Orozco-Suárez, S.; Barrientos, R.; Alatorre, A.; Querejeta, E.; Trujillo-Ferrara, J.G.; Farfán-García, E.D.; Soriano-Ursúa, M.A. Boron-containing compounds on neurons: Actions and potential applications for treating neurodegenerative diseases. J. Inorg. Biochem. 2022, 238, 112027. [Google Scholar] [CrossRef]
- Kilic, A.; Söylemez, R.; Okumuş, V. Design, spectroscopic properties and effects of novel catechol spiroborates derived from Schiff bases in the antioxidant, antibacterial and DNA binding activity. J. Organomet. Chem. 2022, 960, 122228. [Google Scholar] [CrossRef]
- Kilic, A.; Savci, A.; Alan, Y.; Birsen, H. Synthesis and spectroscopic properties of 4, 4′-bipyridine linker bioactive macrocycle boronate esters: Photophysical properties and antimicrobial with antioxidant studies. J. Organomet. Chem. 2021, 941, 121807. [Google Scholar] [CrossRef]
- Korkmaz, M. Boron. In Boron and Human Health; Korkmaz, M., Ed.; Nobel Akademik Yayıncılık: Çankaya, Türkiye, 2020; pp. 47–66. ISBN 978-625-402-341-5. [Google Scholar]
- Sayli, B.S.; Tüccar, E.; Elhan, A.H. An assessment of fertility in boron-exposed Turkish subpopulations. Reprod. Toxicol. 1998, 12, 297–304. [Google Scholar] [CrossRef]
- Bolt, H.M.; Başaran, N.; Duydu, Y. Effects of boron compounds on human reproduction. Arch. Toxicol. 2020, 94, 717–724. [Google Scholar] [CrossRef] [Green Version]
- Smallwood, C. International Program in Chemical Safety. In Boron; World Health Organization: Geneva, Switzerland, 1998; pp. 192–201. [Google Scholar]
- Robbins, W.; Xun, L.; Jia, J.; Kennedy, N.; Elashoff, D.; Ping, L. Chronic boron exposure and human semen parameters. Reprod. Toxicol. 2010, 29, 184–190. [Google Scholar] [CrossRef]
- Weir, R.J.; Fisher, R.S. Toxicologic studies on borax and boric acid. Toxicol. Appl. Pharmacol. 1972, 23, 351–364. [Google Scholar] [CrossRef]
- Scialli, A.R.; Bonde, J.P.; Brüske-Hohlfeld, I.; Culver, B.D.; Li, Y.; Sullivan, F.M. An overview of male reproductive studies of boron with an emphasis on studies of highly exposed Chinese workers. Reprod. Toxicol. 2010, 29, 10–24. [Google Scholar] [CrossRef]
- European Commission. OPINION ON Boron Compounds; European Commission: Brussels, Belgium, 2010. [Google Scholar]
- Whorton, M.D.; Haas, J.L.; Trent, L.; Wong, O. Reproductive effects of sodium borates on male employees: Birth rate assessment. Occup. Environ. Med. 1994, 51, 761–767. [Google Scholar] [CrossRef] [Green Version]
- Marat, I.; Arstan, M.; Galymzhan, Y.; Timur, J.; Yerbolat, I.; Almasbek, Y. Impact of chromium and boron compounds on the reproductive function in rats. Toxicol. Ind. Health 2018, 34, 365–374. [Google Scholar] [CrossRef]
- Medicines Adverse Reactions Comitte. Boron-Containing Excipients and Fertility Concerns; Medsafe Pharmacovigilance Team: Wellington, New Zealand, 2021. Available online: https://www.medsafe.govt.nz/committees/MARC/reports/186-3.2.1-Boron.pdf (accessed on 14 February 2023).
- Duydu, Y.; Başaran, N.; Yalçın, C.Ö.; Üstündağ, A.; Aydın, S.; Anlar, H.G.; Bacanlı, M.; Aydos, K.; Atabekoğlu, C.S.; Golka, K. Boron-exposed male workers in Turkey: No change in sperm Y: X chromosome ratio and in offspring’s sex ratio. Arch. Toxicol. 2019, 93, 743–751. [Google Scholar] [CrossRef]
- Hadrup, N.; Frederiksen, M.; Sharma, A.K. Toxicity of boric acid, borax and other boron containing compounds: A review. Regul. Toxicol. Pharmacol. 2021, 121, 104873. [Google Scholar] [CrossRef]
- El-Dakdoky, M.H.; Abd El-Wahab, H.M.F. Impact of boric acid exposure at different concentrations on testicular DNA and male rats fertility. Toxicol. Mech. Methods 2013, 23, 360–367. [Google Scholar] [CrossRef]
- Hunt, C.D.; Nielsen, F.H. Interaction between boron and cholecalciferol in the chick. In Proceedings of the Trace Element Metabolism in Man and Animals, Canberra, Australia, 11–15 May 1981; pp. 597–600. [Google Scholar]
- Hunt, C.D. The biochemical effects of physiologic amounts of dietary boron in animal nutrition models. Env. Health Perspect. 1994, 102, 35–43. [Google Scholar]
- Rosen, V.; Wozney, J.M. Bone morphogenetic proteins. In Principles of Bone Biology; Academic Press: Cambridge, MA, USA, 2002; Volume 2, pp. 919–928. [Google Scholar]
- Naghii, M.R.; Torkaman, G.; Mofid, M. Effects of boron and calcium supplementation on mechanical properties of bone in rats. BioFactors 2006, 28, 195–201. [Google Scholar] [CrossRef]
- Hunter, J.M.; Nemzer, B.V.; Rangavajla, N.; Biţă, A.; Rogoveanu, O.C.; Neamţu, J.; Scorei, I.R.; Bejenaru, L.E.; Rău, G.; Bejenaru, C.; et al. The fructoborates: Part of a family of naturally occurring sugar–borate complexes—Biochemistry, physiology, and impact on human health: A review. Biol. Trace Elem. Res. 2019, 188, 11–25. [Google Scholar] [CrossRef] [Green Version]
- Sheng, M.H.; Taper, L.J.; Veit, H.; Qian, H.; Ritchey, S.J.; Lau, K.H. Dietary boron supplementation enhanced the action of estrogen, but not that of parathyroid hormone, to improve trabecular bone quality in ovariectomized rats. Biol. Trace Elem. Res. 2001, 82, 109–123. [Google Scholar] [CrossRef]
- Dupre, J.N.; Keenan, M.J.; Hegsted, M.; Brudevold, A.M. Effects of dietary boron in rats fed a vitamin D-deficient diet. Environ. Health Perspect. 1994, 102, 55–58. [Google Scholar]
- Kameda, T.; Mano, H.; Yuasa, T.; Mori, Y.; Miyazawa, K.; Shiokawa, M.; Nakamaru, Y.; Hiroi, E.; Hiura, K.; Kameda, A.; et al. Estrogen inhibits bone resorption by directly inducing apoptosis of the bone-resorbing osteoclasts. J. Exp. Med. 1997, 186, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Cauley, J.A.; Robbins, J.; Chen, Z. Effect of boron supplementation of pig diets on the production of tumor necrosis factor-alpha and interferon-gamma. Jama 2003, 290, 1729–1738. [Google Scholar] [CrossRef] [Green Version]
- Horowitz, M.C. Cytokines and estrogen in bone: Anti-osteoporotic effects. Science 1993, 260, 626–628. [Google Scholar] [CrossRef]
- Holick, M.F. Sunlight and vitamin D for bone health and prevention of autoimmune diseases, cancers, and cardiovascular disease. Am. J. Clin Nutr. 2004, 80, 1678S–1688S. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Lingxia, Z.; Schrauzer, G.N.; Xiong, G. Selenium, boron, and germanium deficiency in the etiology of Keshan-Beck disease. Biol. Trace Elem. Res. 2000, 77, 193–197. [Google Scholar] [CrossRef]
- Fang, W.; Wu, P.; Hu, R.; Huang, Z. Environmental Se-Mo-B deficiency and its possible effects on crops and Keshan-Beck disease (KBD) in the Chousang area, Yao County, Shaanxi Province, China. Environ. Geochem. Health 2003, 25, 267–280. [Google Scholar] [CrossRef]
- Hunt, C.D.; Idso, J.P. Dietary boron as a physiological regulator of the normal inflammatory response: A review and current research progress. J. Trace Elem. Exp. Med. 1999, 12, 221–233. [Google Scholar] [CrossRef]
- Volpe, S.L.; Taper, L.J.; Meacham, S. The relationship between boron and magnesium status and bone mineral density in the human: A review. Magnes Res. 1993, 6, 291–296. [Google Scholar]
- Nielsen, F.H.; Mullen, L.M.; Gallagher, S.K. Effect of boron depletion and repletion on blood indicators of calcium status in humans fed a magnesium-low diet. J. Trace Elem. Exp. Med. 1990, 3, 45–54. [Google Scholar]
- Miljkovic, D.; Scorei, R.I.; Cimpoiaşu, V.M.; Scorei, I.D. Calcium fructoborate: Plant-based dietary boron for human nutrition. J. Diet Suppl. 2009, 6, 211–226. [Google Scholar] [CrossRef]
- Miljkovic, D.; Miljkovic, N.; McCarty, M.F. Up-regulatory impact of boron on vitamin D function–Does it reflect inhibition of 24-hydroxylase? Med. Hypotheses 2004, 63, 1054–1056. [Google Scholar] [CrossRef]
- Hegsted, M.; Keenan, M.J.; Siver, F.; Wozniak, P. Effect of boron on vitamin D deficient rats. Biol. Trace Elem. Res. 1991, 28, 243–255. [Google Scholar] [CrossRef]
- Hunt, C.D.; Herbel, J.L.; Idso, J.P. Dietary boron modifies the effects of vitamin D3 nutriture on indices of energy substrate utilization and mineral metabolism in the chick. J. Bone Min. Res. 1994, 9, 171–181. [Google Scholar] [CrossRef]
- Franceschi, R.T.; Ge, C.; Xiao, G.; Roca, H.; Jiang, D. Transcriptional regulation of osteoblasts. Cells Tissues Organs. 2009, 189, 144–152. [Google Scholar] [CrossRef]
- Korolev, I.N.; Panova, L.N.; Bobkova, A.S.; Korovkina, E.G. Morphofunctional characteristics of the thyroid and a change in the level of thyroid hormones in the blood from the internal use of boron-containing waters. Vopr. Kurortol. Fizioter. Lech. Fiz. Kult. 1989, 3, 28–31. [Google Scholar]
- Popova, E.V.; Tinkov, A.A.; Ajsuvakova, O.P.; Skalnaya, M.G.; Skalny, A.V. Boron–A potential goiterogen? Med. Hypotheses 2017, 104, 63–67. [Google Scholar] [CrossRef]
- Kucukkurt, I.; Akbel, E.; Karabag, F.; Ince, S. The effects of dietary boron compounds in supplemented diet on hormonal activity and some biochemical parameters in rats. Toxicol. Ind. Health 2015, 31, 255–260. [Google Scholar] [CrossRef]
- Armstrong, T.A.; Spears, J.W.; Lloyd, K.E. Inflammatory response, growth, and thyroid hormone concentrations are affected by long-term boron supplementation in gilts. J. Anim. Sci. 2001, 79, 1549–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Wahab, A.; Ibrahim, S.S.; El-Anwar, A.H.; Mabrook, E.A.; Ibrahim, T.B.; Abdel-Razik, A.-R.H. Effects of dietary boron supplementation on the testicular function and thyroid activity in male goats: Involvement of CYP17A1 gene. Reprod. Domest. Anim. 2022, 57, 1353–1362. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, T.B.; Abdel-Wahab, A.; Aziz, R.L.A.; El-Anwar, A.H.; Ibrahim, S.S. Dietary boron supplementation and its impact on testicular function, thyroid activity and serum calcium in rams. Small Rumin. Res. 2019, 174, 156–162. [Google Scholar] [CrossRef]
- Mirzaei, H.R.; Sahebkar, A.; Salehi, R.; Nahand, J.S.; Karimi, E.; Jaafari, M.R.; Mirzaei, H. Boron neutron capture therapy: Moving toward targeted cancer therapy. J. Cancer Res. Ther. 2016, 12, 520. [Google Scholar] [CrossRef] [PubMed]
- Pisarev, M.A.; Dagrosa, M.A.; Thomasz, L.; Juvenal, G. Boron neutron capture therapy applied to undifferentiated thyroid carcinoma. Medicina (B. Aires) 2006, 66, 569–573. [Google Scholar] [PubMed]
- Luca, E.; Fici, L.; Ronchi, A.; Marandino, F.; Rossi, E.D.; Caristo, M.E.; Malandrino, P.; Russo, M.; Pontecorvi, A.; Vigneri, R. Intake of Boron, Cadmium, and Molybdenum enhances rat thyroid cell transformation. J. Exp. Clin. Cancer Res. 2017, 36, 73. [Google Scholar] [CrossRef]
- Rodriguez, C.; Carpano, M.; Curotto, P.; Thorp, S.; Casal, M.; Juvenal, G.; Pisarev, M.; Dagrosa, M.A. In vitro studies of DNA damage and repair mechanisms induced by BNCT in a poorly differentiated thyroid carcinoma cell line. Radiat. Environ. Biophys. 2018, 57, 143–152. [Google Scholar] [CrossRef]
- Pan, Y.-Y.; Yao, S.-F.; Lin, K.-H.; Chou, F.-I.; Lee, J.-C.; Tai, S.-K.; Huang, W.-S.; Lan, K.-L.; Chao, Y.; Chen, Y.-W. Boron neutron capture therapy as salvage treatment for recurrent papillary thyroid carcinoma—A case report. Ther. Radiol. Oncol. 2020, 4, 21. [Google Scholar] [CrossRef]
- Perona, M.; Majdalani, M.E.; Rodríguez, C.; Nievas, S.; Carpano, M.; Rossini, A.; Longhino, J.M.; Cabrini, R.; Pisarev, M.A.; Juvenal, G.J.; et al. Experimental studies of boron neutron capture therapy (BNCT) using histone deacetylase inhibitor (HDACI) sodium butyrate, as a complementary drug for the treatment of poorly differentiated thyroid cancer (PDTC). Appl. Radiat. Isot. 2020, 164, 109297. [Google Scholar] [CrossRef]
- Pulagam, K.R.; Gona, K.B.; Gómez-Vallejo, V.; Meijer, J.; Zilberfain, C.; Estrela-Lopis, I.; Baz, Z.; Cossío, U.; Llop, J. Gold nanoparticles as boron carriers for boron neutron capture therapy: Synthesis, radiolabelling and in vivo evaluation. Molecules 2019, 24, 3609. [Google Scholar] [CrossRef] [Green Version]
- Hosseinzadeh, B.; Salimi Beni, A.; Eskandari, R.; Karami, M.; Khorram, M. Interaction of propylthiouracil, an anti-thyroid drug with boron nitride nanotube: A DFT study. Adsorption 2020, 26, 1385–1396. [Google Scholar] [CrossRef]
- Ou, M.; Wang, X.; Yu, L.; Liu, C.; Tao, W.; Ji, X.; Mei, L. The emergence and evolution of borophene. Adv. Sci. 2021, 8, 2001801. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Sadique, M.A.; Kaushik, A.; Ranjan, P.; Khan, R.; Srivastava, A.K. Borophene as an emerging 2D flatland for biomedical applications: Current challenges and future prospects. J. Mater. Chem. B 2022, 10, 1146–1175. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 (μM) 1 |
|---|---|
| GW5638 | 0.39 |
| GW7604 | 0.0017 |
| GLL398 | 1.14 |
| ZB716 | 4.1 |
| Compound | t ½ (h) | Cmax (μg/mL) | AUC (μg·h/mL) |
|---|---|---|---|
| GLL398 | 3.9 | 3.51 | 36.9 |
| ZB716 | 23.5 | 0.16 | 2.5 |
| Compound | RBA | Selectivity ERβ/ERα | IC50 (μM) | EC50 (μM) | |
|---|---|---|---|---|---|
| ERα | ERβ | ||||
| 17β-E2 | 100 | 100 | 1.0 | 1.0 | 1.0 |
| BE360 | 68 | 46 | 0.68 | 0.58 | 0.024 |
| BE310 | 15 | 16 | 1.1 | 0.63 | 0.85 |
| Control | B-125 | B-250 | B-500 | |
|---|---|---|---|---|
| Final body weight (g) | 299.6 ± 5.36 | 300.5 ± 7.86 | 281.3 ± 8.90 | 267.7 ± 3.88 a,b |
| Body weight gain (g) | 120.5 ± 3.63 | 123.0 ± 6.56 | 102.3 ± 7.43 a,b | 96.3 ± 5.48 a,b |
| R. testis weight (g) | 1.57 ± 0.03 | 1.49 ± 0.04 | 1.38 ± 0.10 a | 0.46 ± 0.02 a,b,c |
| R. epididymis weight (g) | 0.58 ± 0.02 | 0.57 ± 0.02 | 0.52 ± 0.04 | 0.35 ± 0.02 a,b,c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estevez-Fregoso, E.; Kilic, A.; Rodríguez-Vera, D.; Nicanor-Juárez, L.E.; Romero-Rizo, C.E.M.; Farfán-García, E.D.; Soriano-Ursúa, M.A. Effects of Boron-Containing Compounds on Liposoluble Hormone Functions. Inorganics 2023, 11, 84. https://doi.org/10.3390/inorganics11020084
Estevez-Fregoso E, Kilic A, Rodríguez-Vera D, Nicanor-Juárez LE, Romero-Rizo CEM, Farfán-García ED, Soriano-Ursúa MA. Effects of Boron-Containing Compounds on Liposoluble Hormone Functions. Inorganics. 2023; 11(2):84. https://doi.org/10.3390/inorganics11020084
Chicago/Turabian StyleEstevez-Fregoso, Elizabeth, Ahmet Kilic, Diana Rodríguez-Vera, Luis E. Nicanor-Juárez, C. Elena M. Romero-Rizo, Eunice D. Farfán-García, and Marvin A. Soriano-Ursúa. 2023. "Effects of Boron-Containing Compounds on Liposoluble Hormone Functions" Inorganics 11, no. 2: 84. https://doi.org/10.3390/inorganics11020084