

HIV Dynamics with a Trilinear Antibody Growth Function and Saturated Infection Rate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Positivity and Boundedness of Solutions
3. Analysis of the Model
3.1. Stability of the Disease-Free Equilibrium
3.2. Infection Steady States
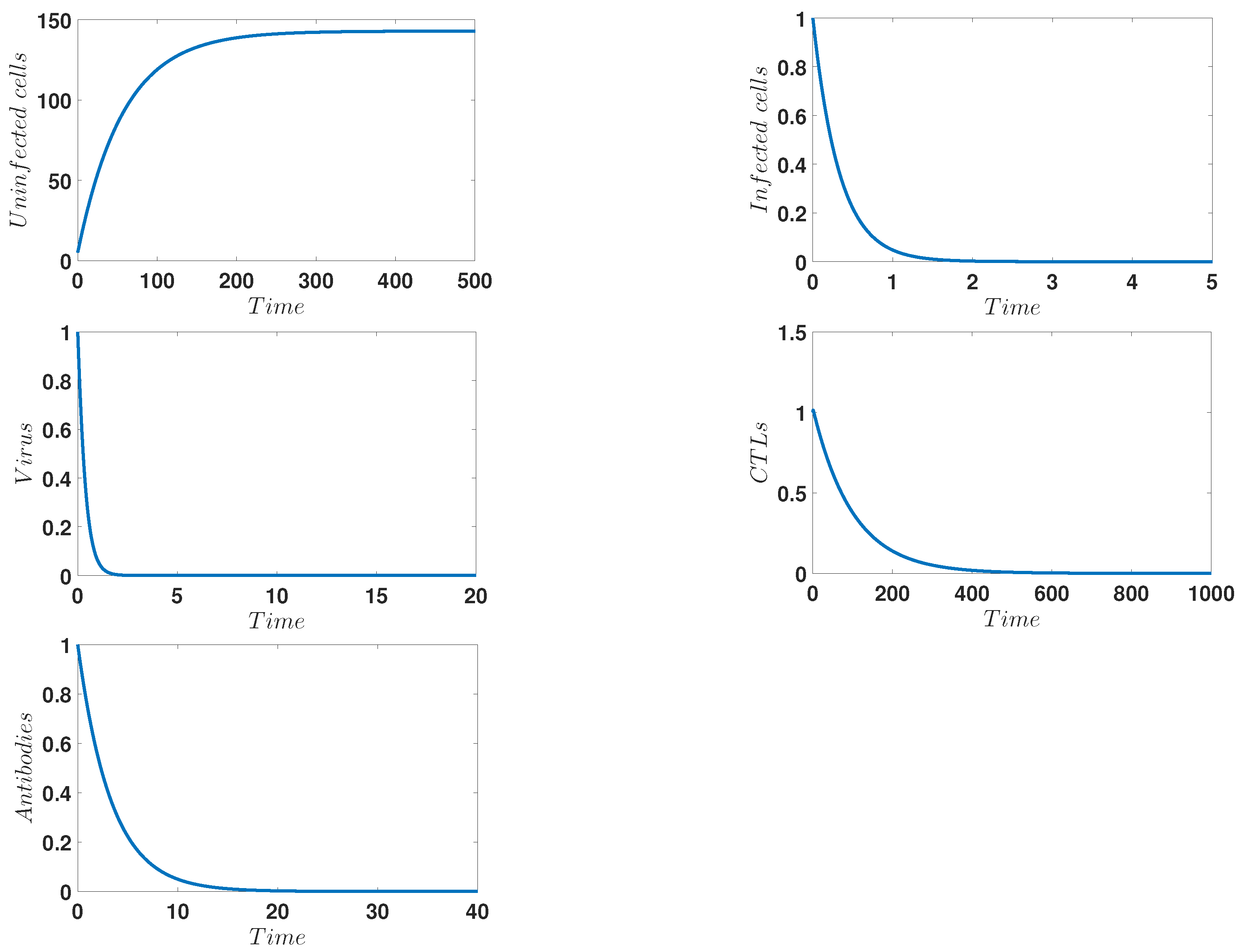
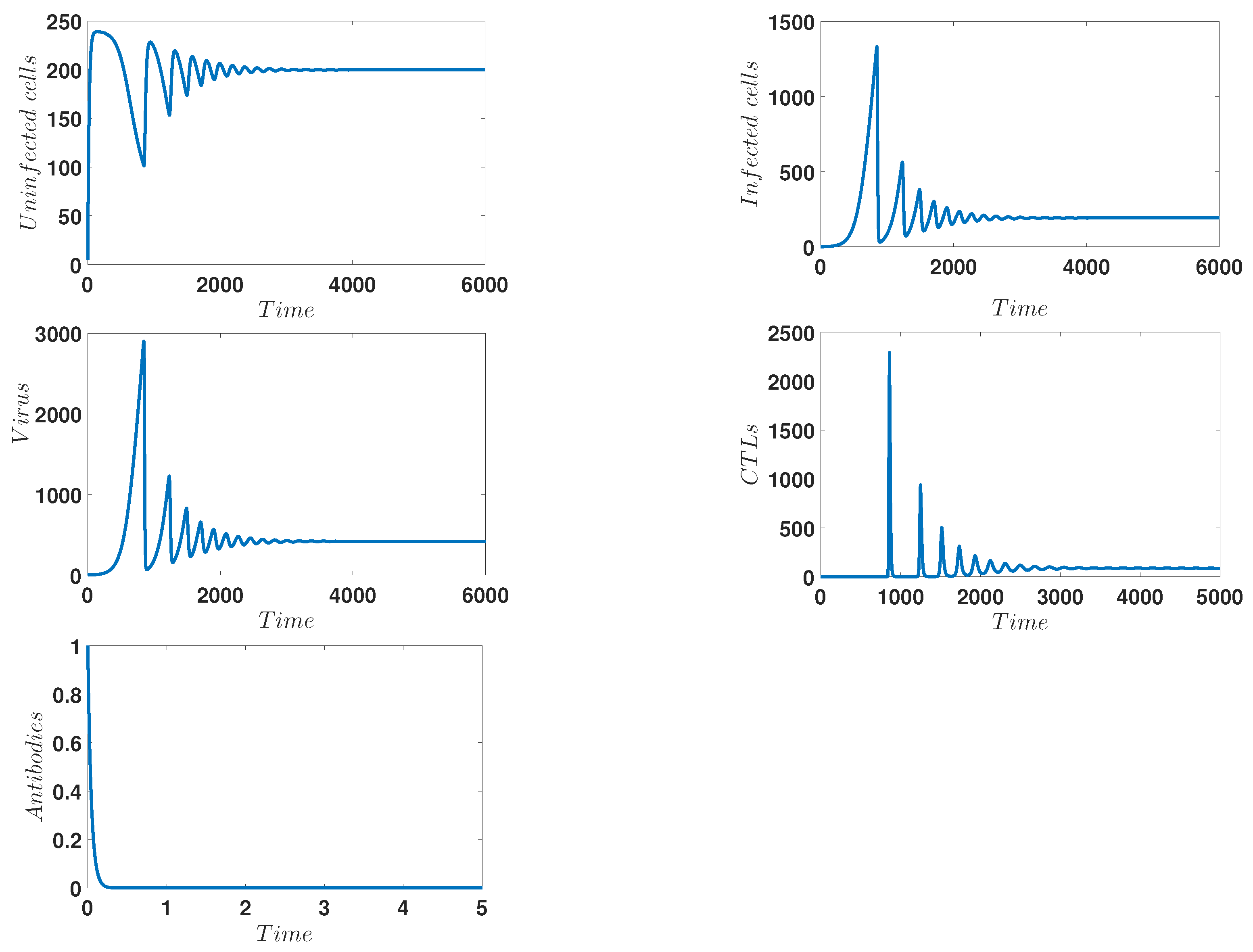
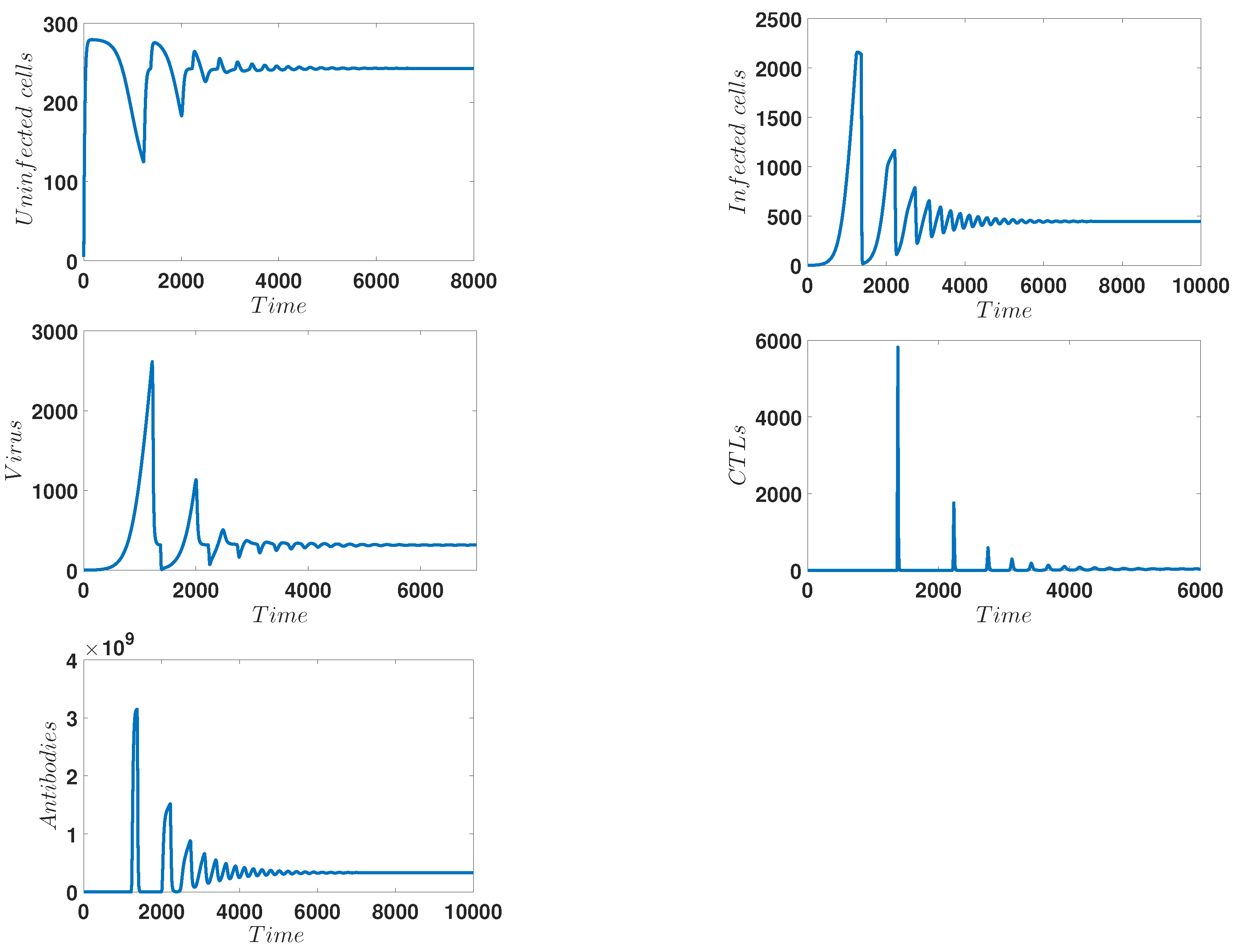
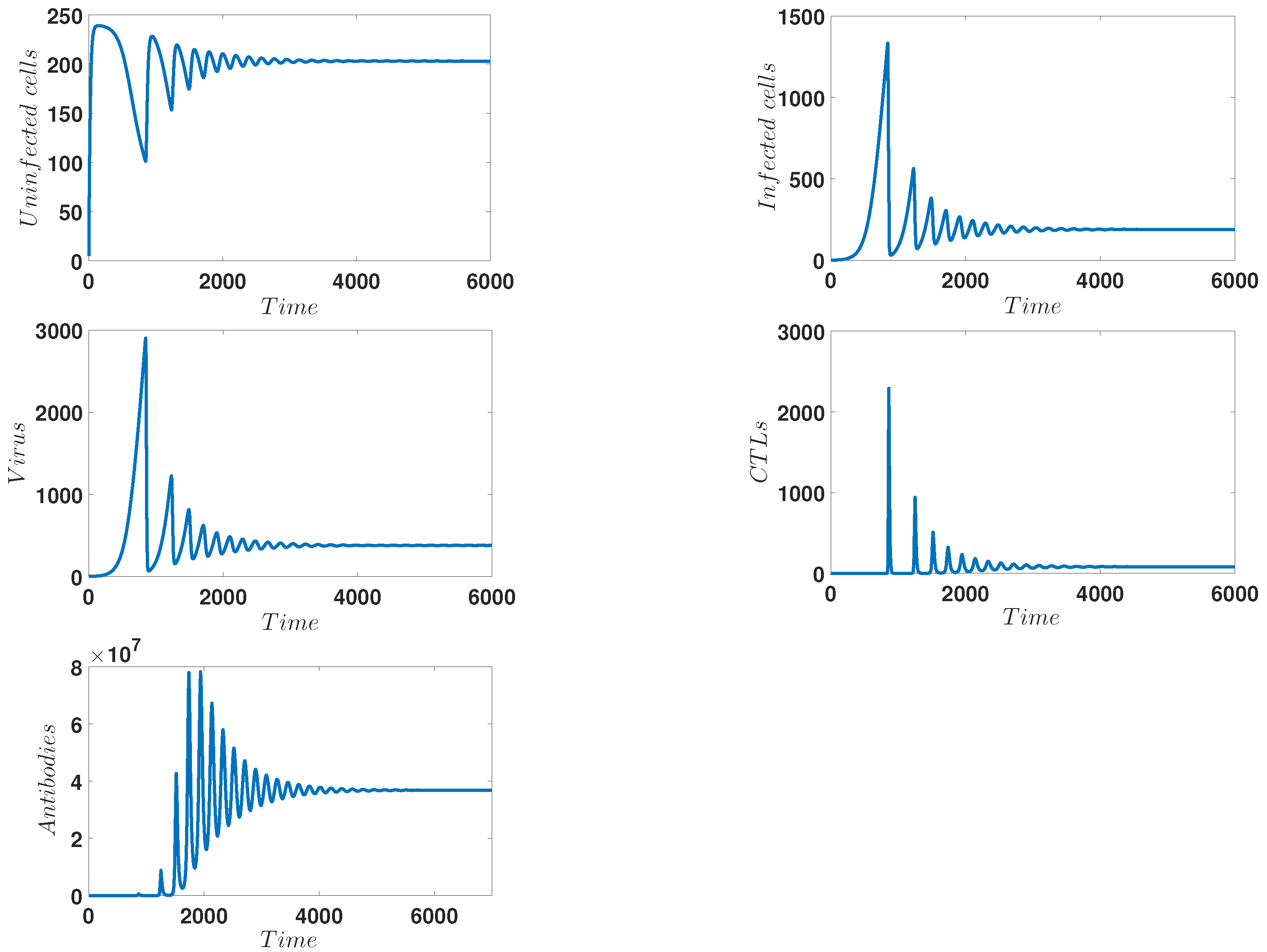
4. Numerical Simulations
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Weiss, R. How does HIV cause AIDS? Science 1993, 260, 1273–1279. [Google Scholar] [CrossRef]
- Blanttner, W.; Gallo, R.; Temin, H. HIV causes aids. Science 1988, 241, 515–516. [Google Scholar] [CrossRef]
- Bairagi, N.; Adak, D. Global analysis of HIV-1 dynamics with Hill type infection rate and intracellular delay. Appl. Math. Model. 2014, 38, 5047–5066. [Google Scholar] [CrossRef]
- Balasubramaniam, P.; Prakash, M.; Rihan, F.A.; Lakshmanan, S. Hopf bifurcation and stability of periodic solutions for delay differential model of HIV infection of CD4+ T-cells. Abstr. Appl. Anal. 2014, 2014, 838396. [Google Scholar] [CrossRef] [Green Version]
- Rihan, F.A.; Rahman, D.H.A. Delay differential model for tumour–immune dynamics with HIV infection of CD4+ T-cells. Int. J. Comput. Math. 2013, 90, 594–614. [Google Scholar] [CrossRef]
- Nowak, M.; May, R. Mathematical biology of HIV infection: Antigenic variation and diversity threshold. Math. Biosci. 1991, 106, 1–21. [Google Scholar] [CrossRef]
- Perelson, A.; Nelson, P. Mathematical analysis of HIV-1 dynamics in vivo. SIAM Rev. 1999, 41, 3–44. [Google Scholar] [CrossRef] [Green Version]
- Tabit, Y.; Hattaf, K.; Yousfi, N. Dynamics of an HIV pathogenesis model with CTL immune response and two saturated rates. World J. Model. Simul. 2014, 10, 215–223. [Google Scholar]
- Chun, T.W.; Murray, D.; Justement, J.S.; Blazkova, J.; Hallahan, C.W.; Fankuchen, O.; Gittens, K.; Benko, E.; Kovacs, C.; Moir, S.; et al. Broadly neutralizing antibodies suppress HIV in the persistent viral reservoir. Proc. Natl. Acad. Sci. USA 2014, 111, 13151–13156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geib, Y.; Dietrich, U. The race between HIV and neutralizing antibodies. AIDS Rev. 2015, 17, 107–113. [Google Scholar]
- West, A.P., Jr.; Scharf, L.; Scheid, J.F.; Klein, F.; Bjorkman, P.J.; Nussenzweig, M.C. Structural insights on the role of antibodies in HIV-1 vaccine and therapy. Cell 2014, 156, 633–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Yang, K.; Zhou, K.; Wang, C. Optimal Treatment Strategies for HIV with Antibody Response. J. Appl. Math. 2014, 2014, 685289. [Google Scholar] [CrossRef]
- Allali, K.; Harroudi, S.; Torres, D.F.M. Optimal control of HIV model with a trilinear antibody growth function. arXiv 2021, arXiv:2105.10291. [Google Scholar] [CrossRef]
- Wodarz, D. Helper-dependent vs. helper-independent CTL responses in HIV infection: Implications for drug therapy and resistance. J. Theor. Biol. 2001, 213, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, Y.; Brauer, F.; Heffernan, J.M. Viral dynamics model with CTL immune response incorporating antiretroviral therapy. J. Math. Biol. 2013, 67, 901–934. [Google Scholar] [CrossRef]
- Zhu, H.; Zou, X. Dynamics of a HIV-1 infection model with cell-mediated immune response and intracellular delay. Discrete Contin. Dyn. Syst. Ser. B 2009, 12, 511–524. [Google Scholar] [CrossRef]
- Stafford, M.A.; Corey, L.; Cao, Y.; Daar, E.S.; Ho, D.D.; Perelson, A.S. Modeling plasma virus concentration during primary HIV infection. J. Theor. Biol. 2000, 203, 285–301. [Google Scholar] [CrossRef] [Green Version]
- Danane, J.; Allali, K. Mathematical analysis and clinical implications of an HIV Model with adaptive immunity. Comput. Math. Methods Med. 2019, 2019, 7673212. [Google Scholar] [CrossRef]
- Gradshteyn, I.S.; Ryzhik, I.M. Table of integrals, series, and products. Math. Comput. 1982, 39, 747–757. [Google Scholar]
- van den Driessche, P. Reproduction numbers of infectious disease models. Infect. Dis. Model. 2017, 2, 288–303. [Google Scholar] [CrossRef]
- Tabit, Y.; Meskaf, A.; Allali, K. Mathematical analysis of HIV model with two saturated rates, CTL and antibody responses. World J. Model. Simul. 2016, 12, 137–146. [Google Scholar]
- Lim, S.C.; Eab, C.H.; Mak, K.H.; Li, M.; Chen, S.Y. Solving linear coupled fractional differential equations by direct operational method and some applications. Math. Probl. Eng. 2012, 2012, 653939. [Google Scholar] [CrossRef] [Green Version]
- Kartal, S.; Gurcan, F. Discretization of conformable fractional differential equations by a piecewise constant approximation. Int. J. Comput. Math. 2019, 96, 1849–1860. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fikri, F.E.; Allali, K. HIV Dynamics with a Trilinear Antibody Growth Function and Saturated Infection Rate. Math. Comput. Appl. 2022, 27, 85. https://doi.org/10.3390/mca27050085
Fikri FE, Allali K. HIV Dynamics with a Trilinear Antibody Growth Function and Saturated Infection Rate. Mathematical and Computational Applications. 2022; 27(5):85. https://doi.org/10.3390/mca27050085
Chicago/Turabian StyleFikri, Fatima Ezzahra, and Karam Allali. 2022. "HIV Dynamics with a Trilinear Antibody Growth Function and Saturated Infection Rate" Mathematical and Computational Applications 27, no. 5: 85. https://doi.org/10.3390/mca27050085