Lipase Production by Solid-State Cultivation of Thermomyces Lanuginosus on By-Products from Cold-Pressing Oil Production

 , , ,
, , ,
Abstract
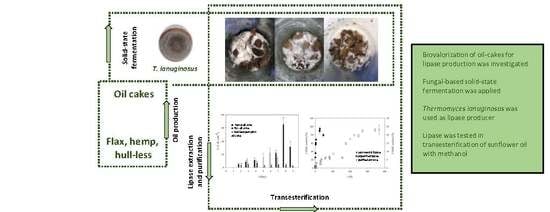
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Substrates
2.2. Microorganism and Commercial Enzyme
2.3. Enzyme Production
2.3.1. Solid-State Fermentation
2.3.2. Enzyme Purification
2.4. Analytical Methods
2.4.1. Measurements of Lipase Activity
2.4.2. Measurement of Fatty Acid Methyl Esters (FAME)
2.4.3. Enzyme Characterization
2.4.4. Biodiesel Production
2.4.5. Data Processing
3. Results
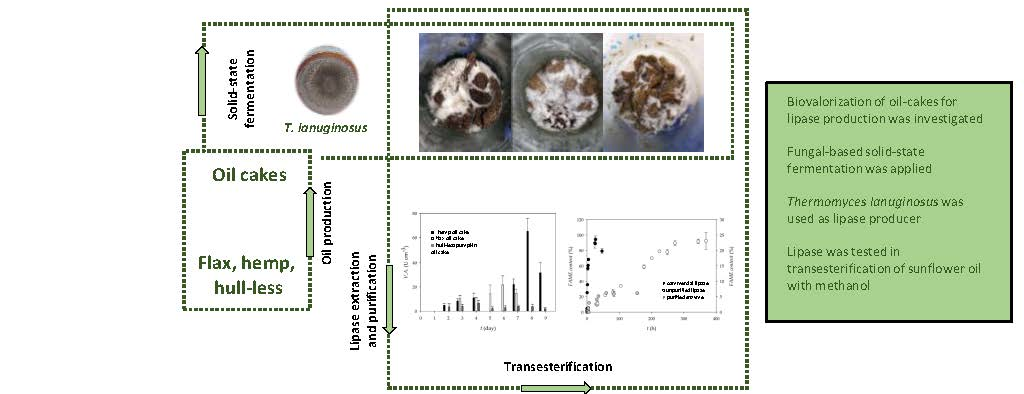
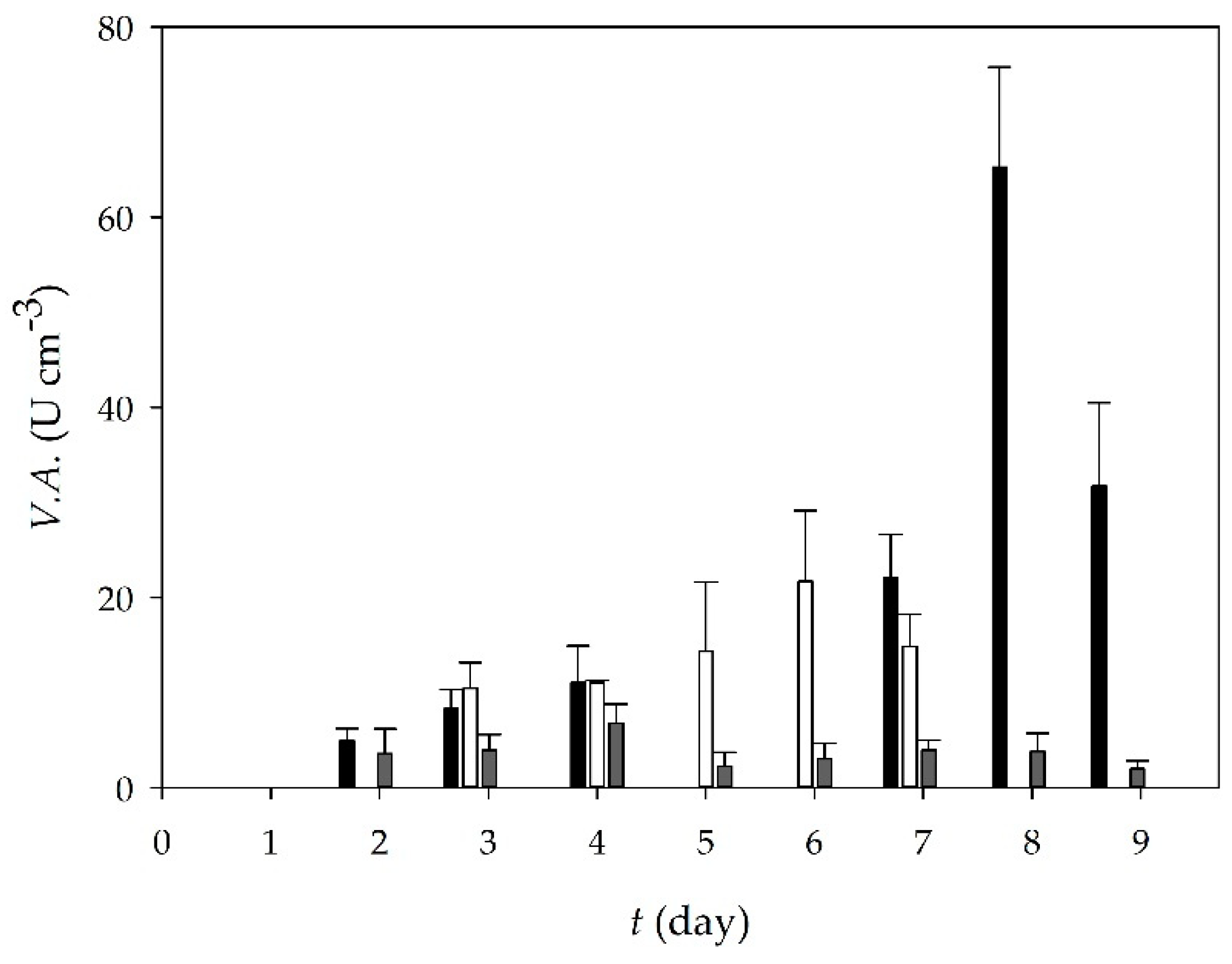
3.1. Lipase Production During Solid-State Cultivation of T. Lanuginosus on Oil Cakes
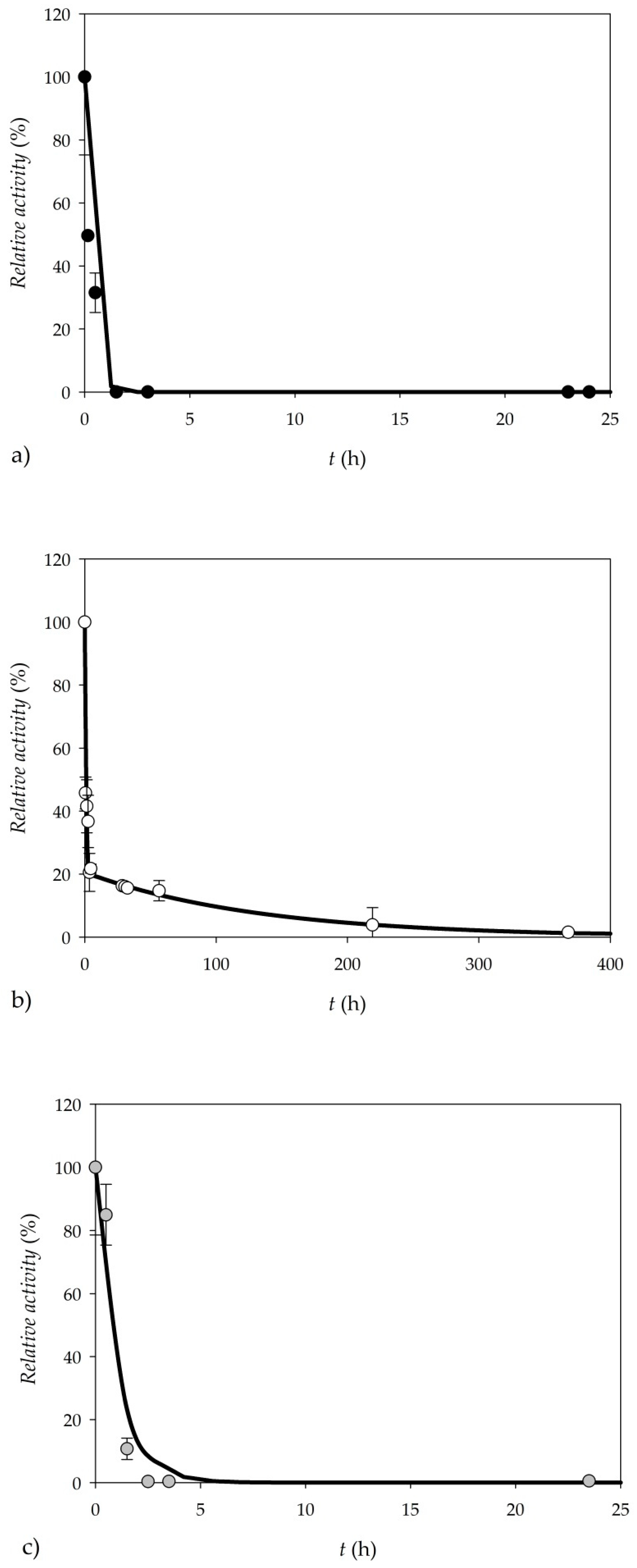
3.2. Enzyme Purification
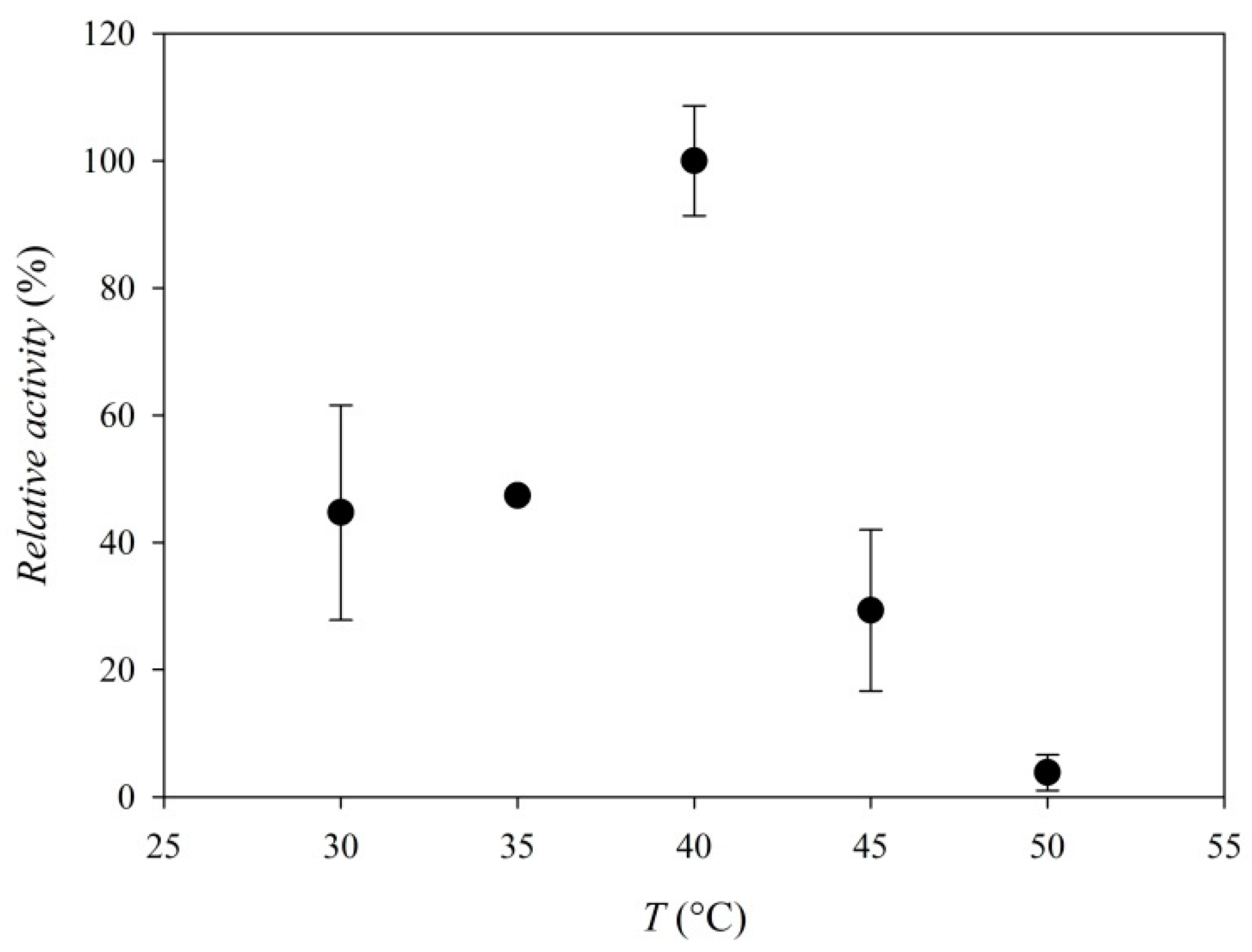
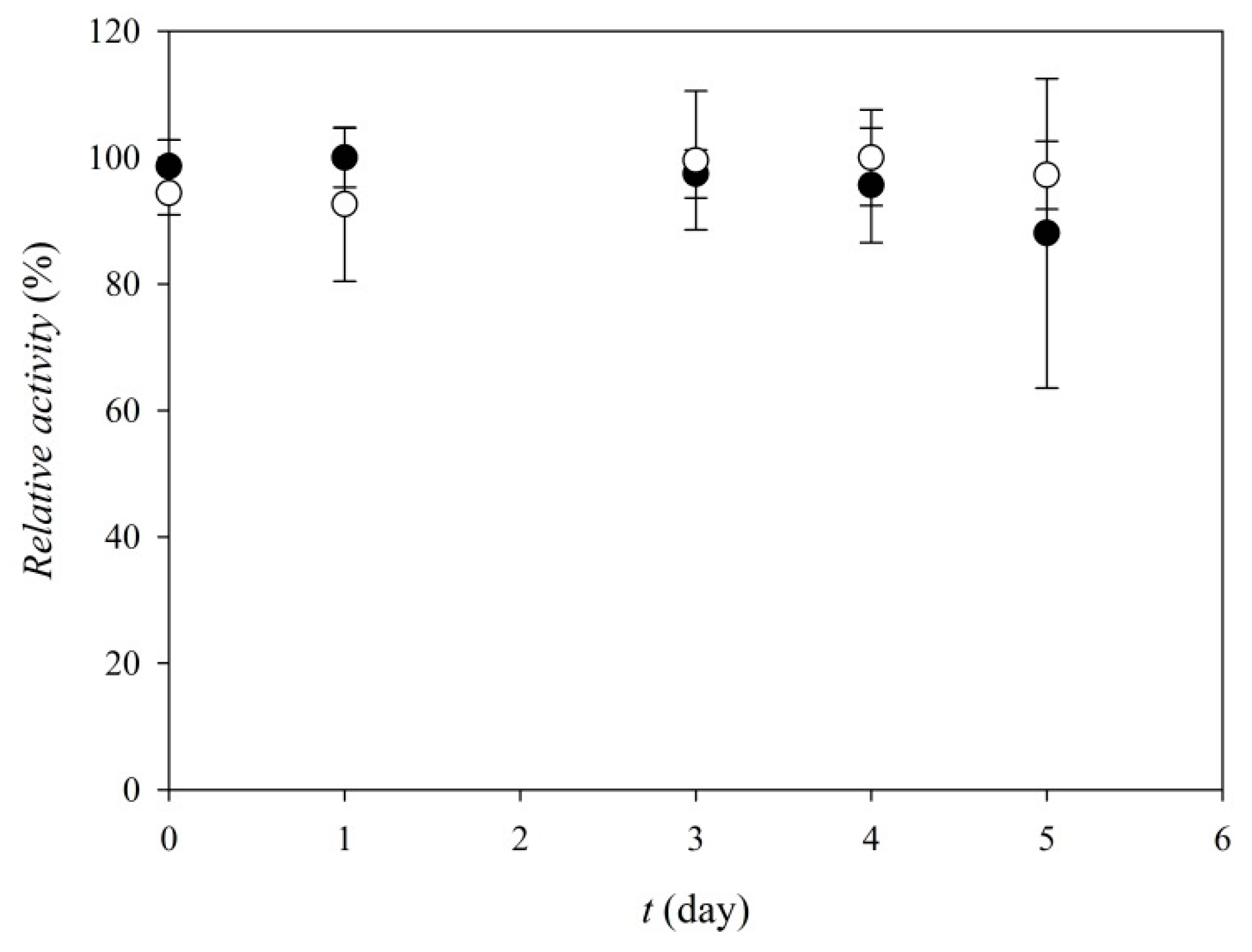
3.3. Enzyme Characterization
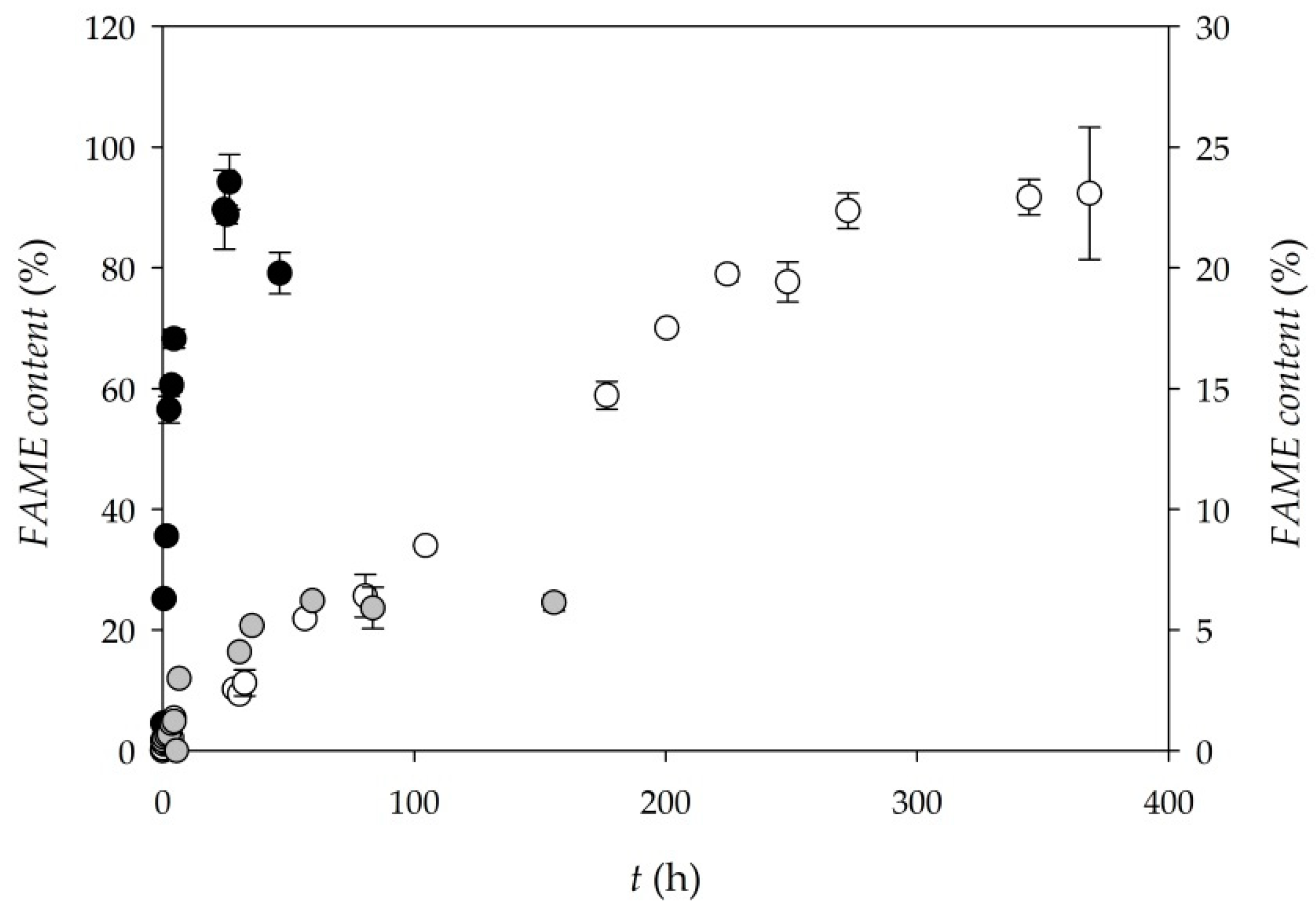
3.4. Biodiesel Production
4. Conclusions and Future Work
Author Contributions
Funding
Conflicts of Interest
References
- Liu, Y.; Huang, L.; Zheng, D.; Fu, Y.; Shan, M.; Xu, Z.; Ma, J.; Lu, F. Development of a Pichia pastoris whole-cell biocatalyst with overexpression of mutant lipase I PCL G47I from Penicillium cyclopium for biodiesel production. RSC Adv. 2018, 8, 26161–26168. [Google Scholar] [CrossRef]
- Nooh, H.M.; Masomian, M.; Salleh, A.B.; Mohamad, R.M.; Ali, S.M.; Rahman, R.N.Z.R.A. Production of thermostable T1 lipase using agroindustrial waste medium formulation. Catalysts 2018, 8, 485. [Google Scholar] [CrossRef]
- Norjannah, B.; Ong, H.C.; Masjuki, H.H.; Juan, J.C.; Chong, W.T. Enzymatic transesterification for biodiesel production: A comprehensive review. Rcs Adv. 2016, 6, 60034–60055. [Google Scholar] [CrossRef]
- DiCosimo, R.; McAuliffe, J.; Poulose, A.J.; Bohlmann, G. Industrial use of immobilized enzymes. Chem. Soc. Rev. 2013, 42, 6437–6474. [Google Scholar] [CrossRef] [PubMed]
- Navvabi, A.; Razzaghi, M.; Fernandes, P.; Karami, L.; Homaei, A. Novel lipases discovery specifically from marine organisms for industrial production and practical applications. Process. Biochem. 2018, 70, 61–70. [Google Scholar] [CrossRef]
- Mohseni, S.; Najafpour, G.D.; Vaseghi, Z.; Mahjoub, S. Solid state fermentation of agricultural residues for lipase production in a tray-bioreactor. World Appl. Sci. J. 2012, 16, 1034–1039. [Google Scholar]
- Aliyah, A.N.; Edelweisis, E.D.; Sahlan, M.; Wijanarko, A.; Hermansyah, H. Solid state fermentation using agroindustrial wastes to produce Aspergillus niger lipase as a biocatalyst immobilized by an adsorption-crosslinking method for biodiesel synthesis. Int. J. Technol. 2016, 8, 1393–1404. [Google Scholar] [CrossRef]
- Silveira, E.A.; Tardioli, P.W.; Farinas, C.S. Valorization of palm oil industrial waste as feedstock for lipase production. Appl. Biochem. Biotechnol. 2016, 179, 558–571. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, E.A.L.; Lima, A.S.; Soares, C.M.F.; de Aquino Santana, L.C.L. Lipase from Aspergillus niger obtained from mangaba residue fermentation: Biochemical characterization of free and immobilized enzymes on a sol-gel matrix. Acta Sci. Technol. 2017, 39, 1–8. [Google Scholar] [CrossRef]
- Vaseghi, Z.; Najafpour, G.D.; Mohseni, S.; Mahjoub, S. Production of active lipase by Rhisopus oryzae from sugarcane bagasse: Solid state fermentation in a tray bioreactor. Int. J. Food Sci. Technol. 2013, 48, 283–289. [Google Scholar] [CrossRef]
- Oliveira, F.; Moreira, C.; Salgado, J.M.; Abrunhosa, L.; Venâncio, A.; Belo, I. Olive pomace valorization by Aspergillus species: Lipase production using solid-state fermentation. J. Sci. Food Agric. 2016, 96, 3583–3589. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.; Salgado, J.M.; Abrunhosa, L.; Prerz-Rodriguez, N.; Dominguez, J.M.; Venancio, A.; Belo, I. Optimization of lipase production by solid-state fermentation of olive pomace: From flask to laboratory-scale packed-bed bioreactor. Bioprocess. Biosyst. Eng. 2017, 40, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Musa, H.; Han, P.C.; Kasim, F.H.; Gopinath, S.C.B.; Ahmad, M.A. Turning oil palm empty fruit bunch waste into substrate for optimal lipase secretion on solid state fermentation by Trichoderma strains. Process. Biochem. 2017, 63, 35–41. [Google Scholar] [CrossRef]
- Malilas, W.; Kang, S.W.; Kim, S.B.; Yoo, H.Y.; Chulalaksananukul, W.; Kim, S.W. Lipase from Penicillium camemberii KCCM 11268: Optimization of solid state fermentation and application to biodiesel production. Korean, J. Chem. Eng 2013, 30, 405–412. [Google Scholar] [CrossRef]
- Singh, M.K.; Singh, J.; Kumar, M.; Thakur, I.S. Novel lipase from basidiomycetes Schizophyllum commune ISTL04 produced by solid state fermentation of Leucaena leucocephala seeds. J. Mol. Catal. B Enzym 2014, 440, 92–99. [Google Scholar] [CrossRef]
- Fernandez-Laufente, R. Lipase from Thermomyces lanuginosus: Uses and prospects as an industrial biocatalyst. J. Mol. Catal. B Enzym. 2010, 62, 197–212. [Google Scholar] [CrossRef]
- Singh, S.; Madlala, A.M.; Prior, B.A. Thermomyces lanuginosus: Properties of strains and their hemicellulases. FEMS Microbiol. Rev. 2003, 27, 3–16. [Google Scholar] [CrossRef]
- Budžaki, S.; Strelec, I.; Krnić, M.; Alilović, K.; Tišma, M.; Zelić, B. Proximate analysis of cold-press oil cakes after biological treatment with Trametes versicolor and Humicola grisea. Eng. Life. Sci. 2018, 18, 924–931. [Google Scholar] [CrossRef]
- Budžaki, S.; Šalić, A.; Zelić, B.; Tišma, M. Enzyme-catalysed biodiesel production from edible and waste cooking oils. Chem Biochem Eng Q. 2015, 29, 329–333. [Google Scholar] [CrossRef]
- Brzozowski, A.M.; Savage, H.; Verma, C.S.; Turkenburg, J.P.; Lawson, D.M.; Svendsen, A. Structural origins of the interfacial activation in Thermomyces (Humicola) lanuginosa lipase. Biochemistry 2000, 39, 15071–15082. [Google Scholar] [CrossRef]
- Ilmi, M.; Hidayaqt, C.; Hastuti, P.; Heeres, H.J.; van der Maarel, M.J.E.C. Utilisation of Jatropha press cake as substrate in biomass and lipase production from Aspergillus niger 6516 and Rhizomucor miehei CBS 360.62. Biocatal. Agric. Biotechnol. 2017, 9, 103–107. [Google Scholar] [CrossRef]
- Castilho, L.R.; Polato, C.M.S.; Baruque, E.A.; Sant’Anna, G.L., Jr.; Freire, D.M.G. Economic analysis of lipase production by Penicillium restrictum in solid-state and submerged fermentations. Biochem. Eng. J. 2000, 4, 239–247. [Google Scholar] [CrossRef]
- Massadeh, M.; Sabra, F.; Dajani, R.; Arafat, A. Purification of lipase enzyme produced by Bacillus stearothermophilus HU1. In Proceedings of the International Conference on Eco-systems and Biological Sciences (ICEBS’2012), Penang, Malaysia, 19–20 May 2012. [Google Scholar]
- Tripathi, R.; Singh, Y.; Bharti, R.K.; Thakur, I.S. Isolation, purification and characterization of lipase from Microbacterium sp. and its application in biodiesel production. Energy Procedia 2014, 54, 518–529. [Google Scholar] [CrossRef]
- Ugur, A.; Sarac, N.; Boran, R.; Ayaz, B.; Ceylan, O.; Okmen, G. New lipase for biodiesel production: Partial purification and characterization of LipSB 25-4. Biochemistry 2015, 14, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Ogundero, V.W. Partial purification and activities of an extracellular lipase from Thermomces lanuginosus from Nigerian palm produce. Mycopathologia 1987, 97, 105–109. [Google Scholar] [CrossRef]
- Zhao, X.; Qi, F.; Yuan, C.; Du, W.; Liu, D. Lipase-catalyzed process for biodiesel production: Enzyme immobilization: Process simulation and optimization. Renew. Sust. Energy Rev. 2014, 44, 182–197. [Google Scholar] [CrossRef]
- Arumugam, A.; Ponnusami, V. Production of biodiesel by enzymatic transesterification of waste sardine oil and evaluation of its engine performance. Heliyon 2017, 3. [Google Scholar] [CrossRef]
- Stergiou, P.Y.; Foukis, A.; Filippou, M.; Koukouritaki, M.; Parapouli, M.; Theodorou, L.G.; Papamichael, E.M. Advances in lipase-catalyzed esterification reactions. Biotechnol. Adv. 2013, 31, 1846–1859. [Google Scholar] [CrossRef]
- Hama, S.; Kondo, A. Enzymatic biodiesel production: An overview of potential feedstocks and process development. Bioresour. Technol. 2013, 135, 386–395. [Google Scholar] [CrossRef]
- Verma, P.; Sharma, M.P. Review of process parameters for biodiesel production from different feedstocks. Renew. Sust. Energ. Rev. 2016, 62, 1063–1071. [Google Scholar] [CrossRef]
- Christopher, L.P.; Kumar, H.; Zambare, V.P. Enzymatic biodiesel: Challenges and opportunities. Appl. Energy. 2014, 119, 497–520. [Google Scholar] [CrossRef]
- Robles-Medina, A.; Gonzalez-Moreno, P.A.; Esteban-Cerdán, L.; Molina-Grima, E. Biocatalysis: Towards ever greener biodiesel production. Biotechnol. Adv. 2009, 27, 398–408. [Google Scholar] [CrossRef]
- Marchaetti, J.M.; Miguel, V.U.; Errazu, A.F. Possible methods for biodiesel production. Renew. Sust. Energ. Rev. 2007, 11, 1300–1311. [Google Scholar] [CrossRef]
- Torres, C.F.; Fornari, T.; Tenllado, D.; Senoráns, F.J.; Reglero, G. A predictive kinetic study of lipase-catalyzed ethanolysis reactions for the optimal reutilization of the biocatalyst. Biochem. Eng. J 2008, 42, 105–110. [Google Scholar] [CrossRef]
- Ranganathan, S.V.; Narasimhan, S.L.; Muthukumar, K. An overview of enzymatic production of biodiesel. Bioresour. Technol. 2008, 99, 3975–3981. [Google Scholar] [CrossRef]
- Kamal, M.d.Z.; Yedavalli, P.; Deshmukh, M.V.; Rao, N.-M. Lipase in aqueous-polar organic solvents: Activity, structure, and stability. Protein Sci. 2013, 22, 904–915. [Google Scholar] [CrossRef] [Green Version]
- Maceiras, R.; Vega, M.; Costa, C.; Ramos, P.; Márquez, M.C. Enzyme deactivation during biodiesel production. Chem. Eng. J. 2011, 166, 358–361. [Google Scholar] [CrossRef]
- Salis, A.; Svensson, I.; Monduzzi, M.; Solinas, V.; Adlercreutz, P. The atypical lipase B from Candida antarctica is better adapted for organic media than the typical lipase from Thermomyces lanuginose. Biochim. Biophys. Acta 2003, 1646, 145–151. [Google Scholar] [CrossRef]
- Šalić, A.; Jurinjak Tušek, A.; Sander, A.; Zelić, B. Lipase catalysed biodiesel synthesis with integrated glycerol separation in continuously operated microchips connected in series. N. Biotechnol. 2018, 47, 80–88. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tišma, M.; Tadić, T.; Budžaki, S.; Ostojčić, M.; Šalić, A.; Zelić, B.; Tran, N.N.; Ngothai, Y.; Hessel, V. Lipase Production by Solid-State Cultivation of Thermomyces Lanuginosus on By-Products from Cold-Pressing Oil Production. Processes 2019, 7, 465. https://doi.org/10.3390/pr7070465
Tišma M, Tadić T, Budžaki S, Ostojčić M, Šalić A, Zelić B, Tran NN, Ngothai Y, Hessel V. Lipase Production by Solid-State Cultivation of Thermomyces Lanuginosus on By-Products from Cold-Pressing Oil Production. Processes. 2019; 7(7):465. https://doi.org/10.3390/pr7070465
Chicago/Turabian StyleTišma, Marina, Toma Tadić, Sandra Budžaki, Marta Ostojčić, Anita Šalić, Bruno Zelić, Nghiep Nam Tran, Yung Ngothai, and Volker Hessel. 2019. "Lipase Production by Solid-State Cultivation of Thermomyces Lanuginosus on By-Products from Cold-Pressing Oil Production" Processes 7, no. 7: 465. https://doi.org/10.3390/pr7070465