Towards an Aspect-Oriented Design and Modelling Framework for Synthetic Biology

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
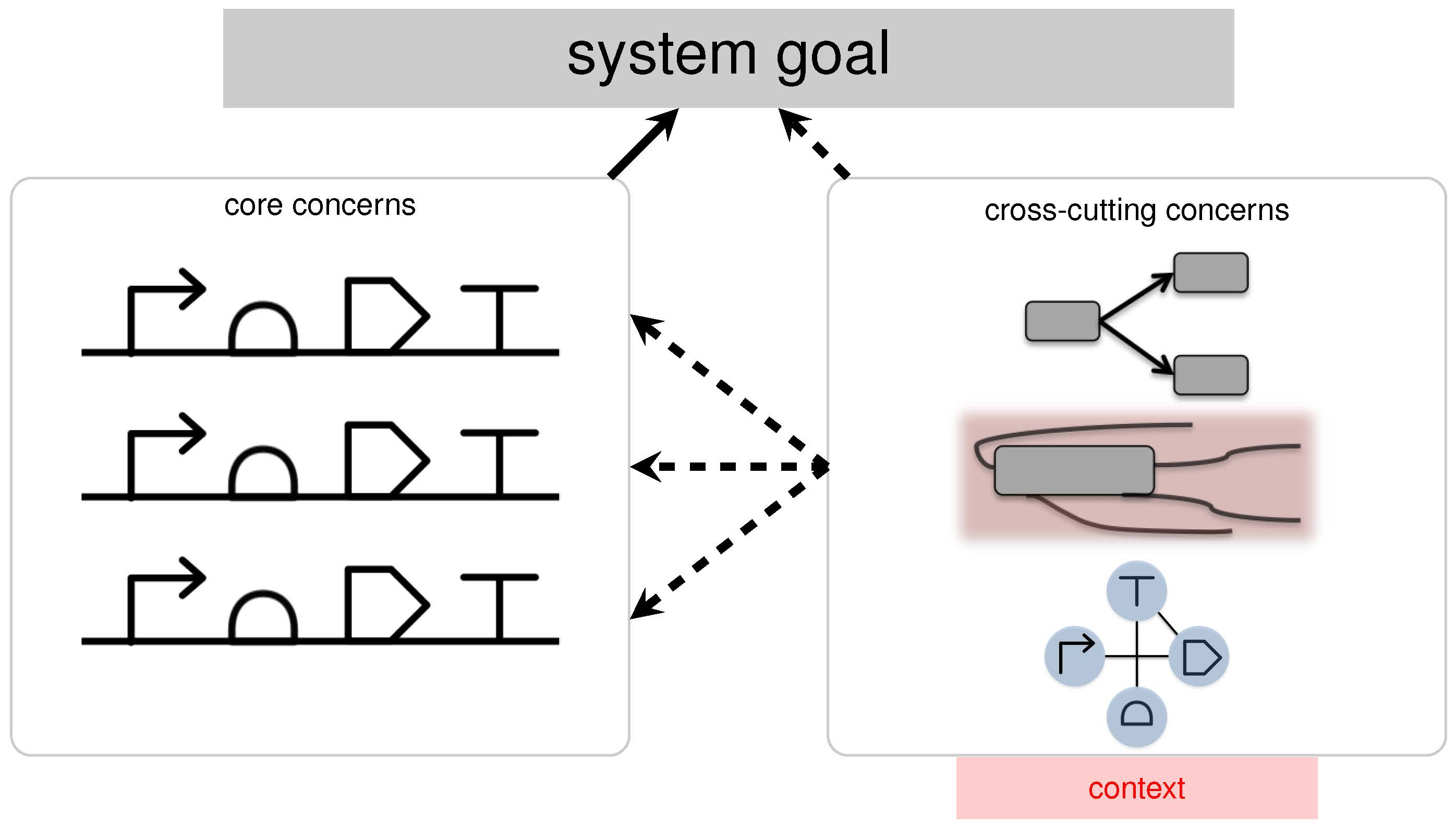
2.1. Concerns: A New Design Paradigm for Synthetic Biology
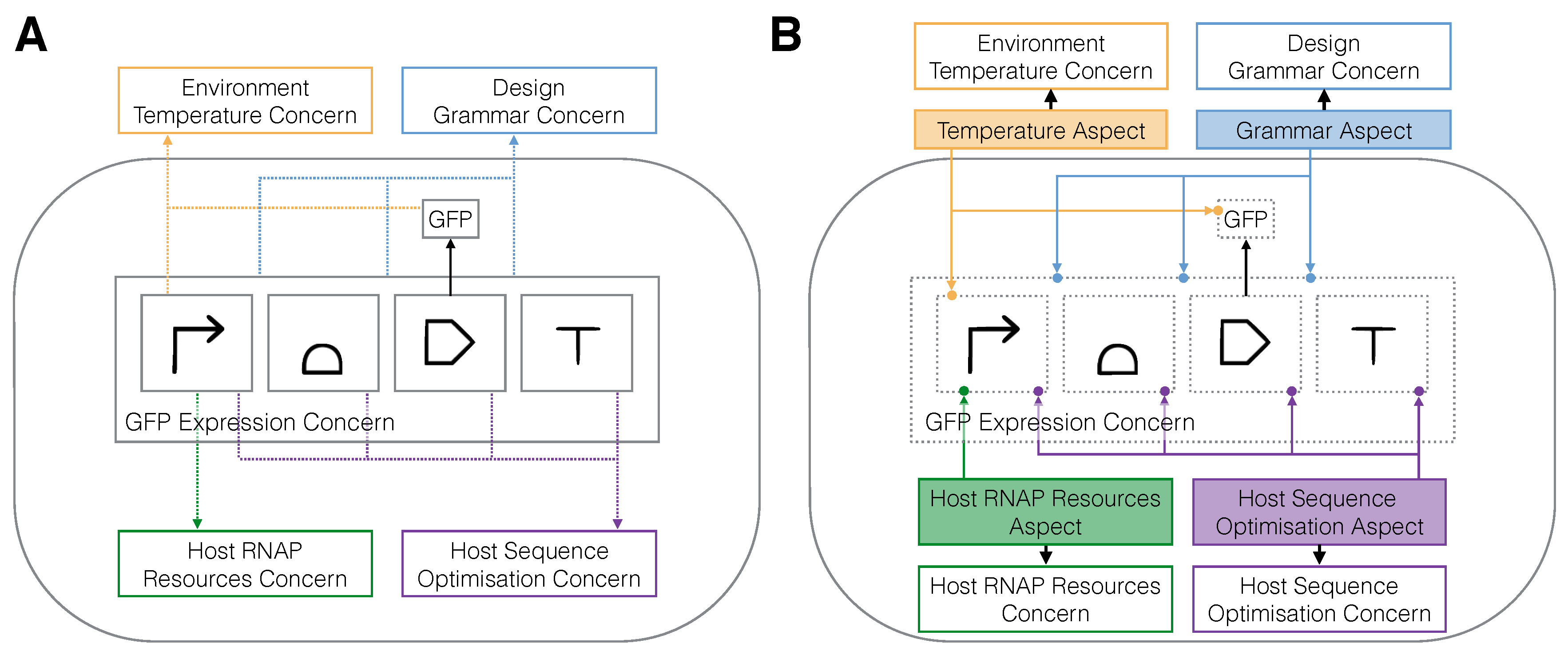
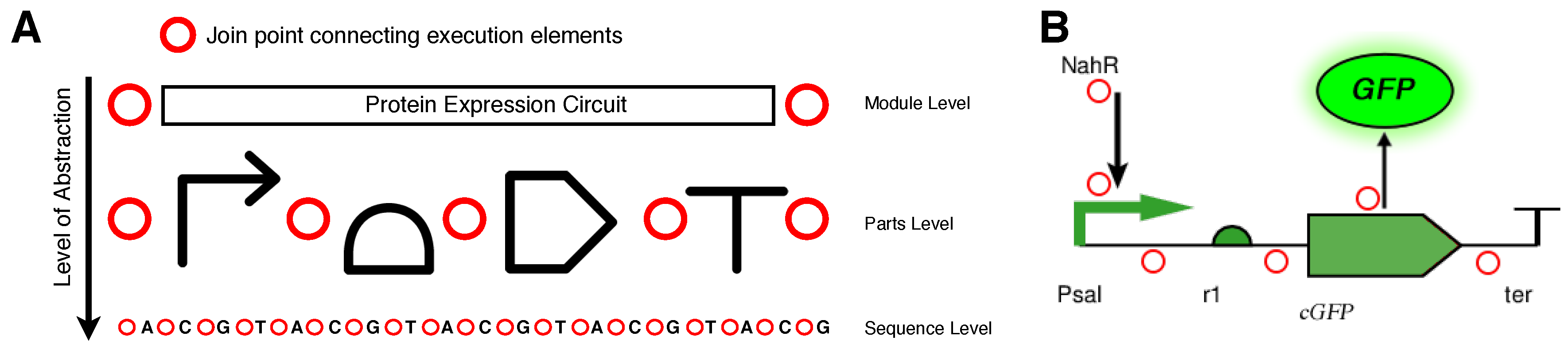
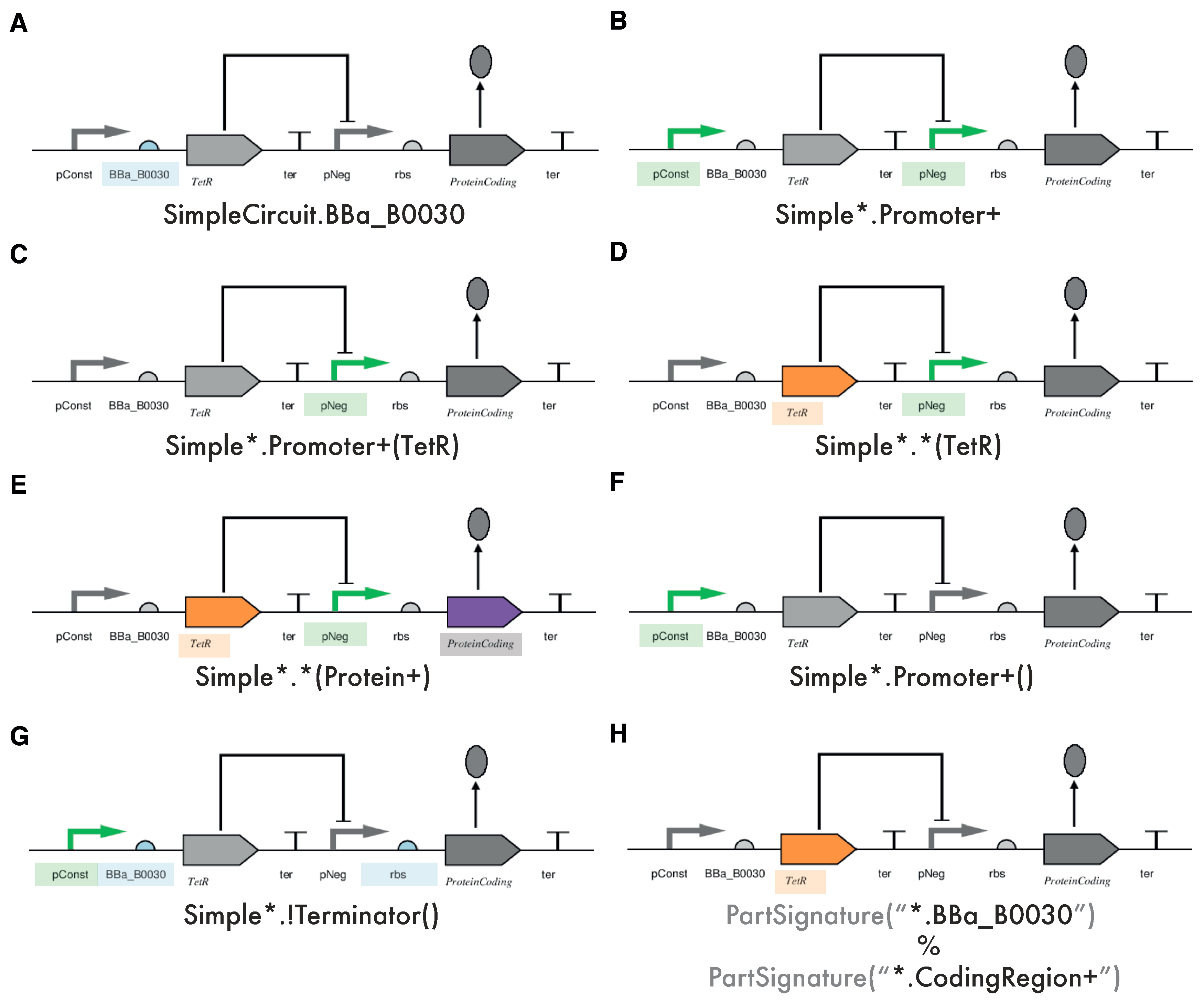
2.2. Aspect-Oriented Synthetic Biology
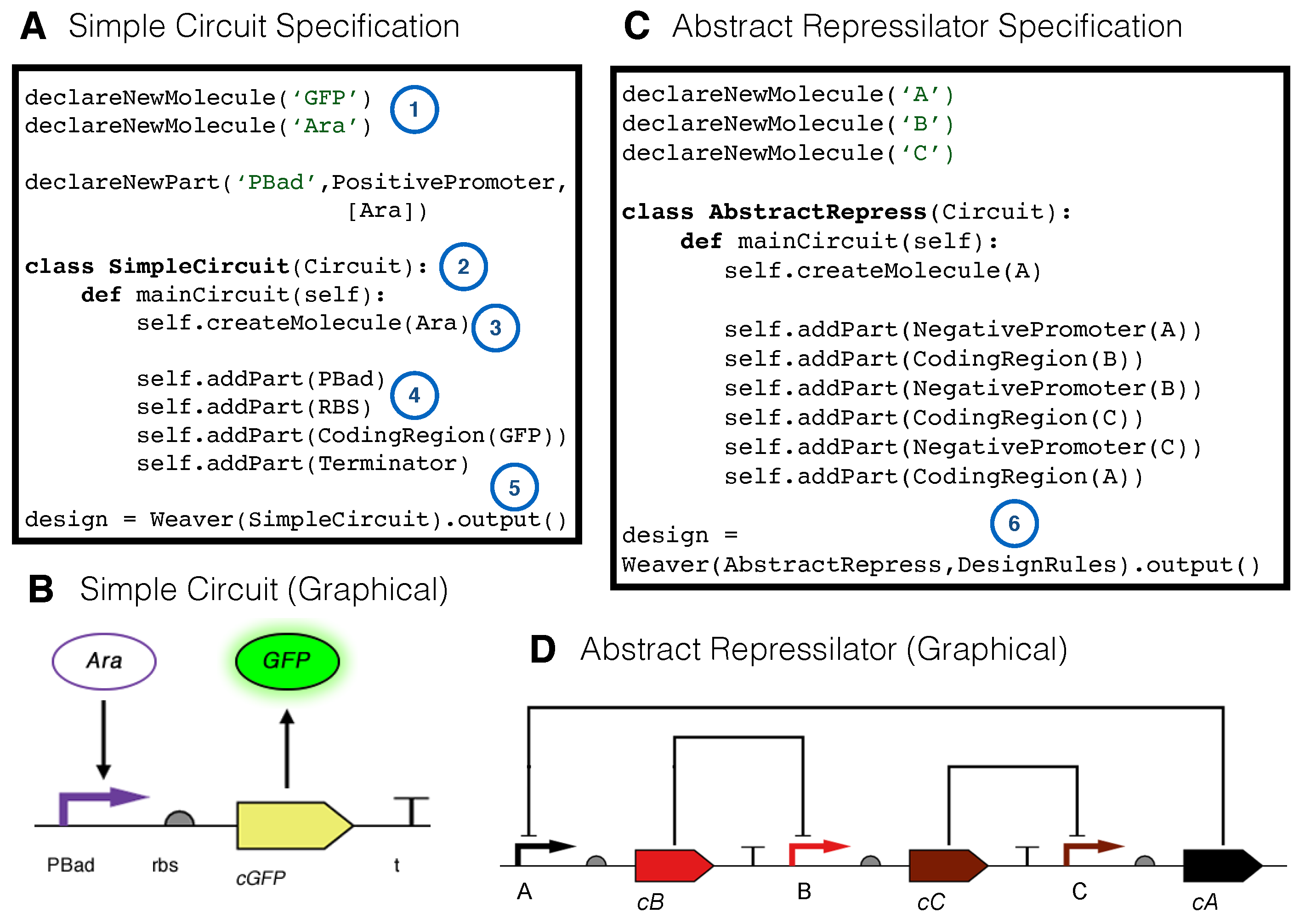
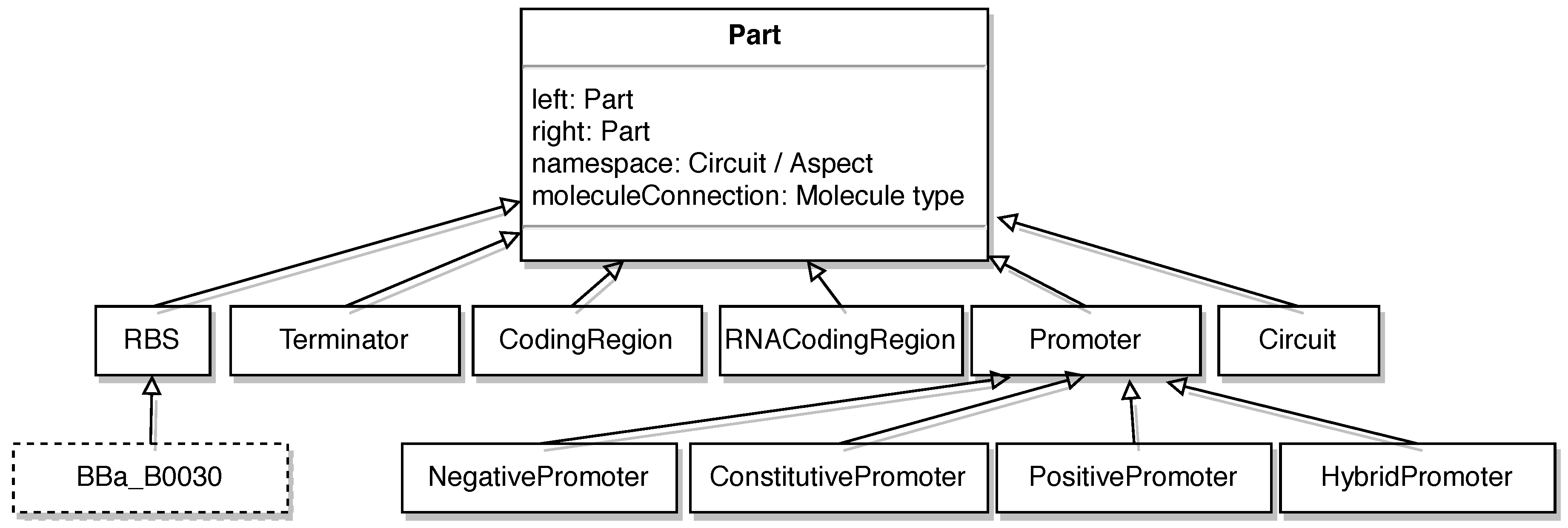
2.3. The SynBioWeaver Framework
2.4. A Simple Design Constraint Example
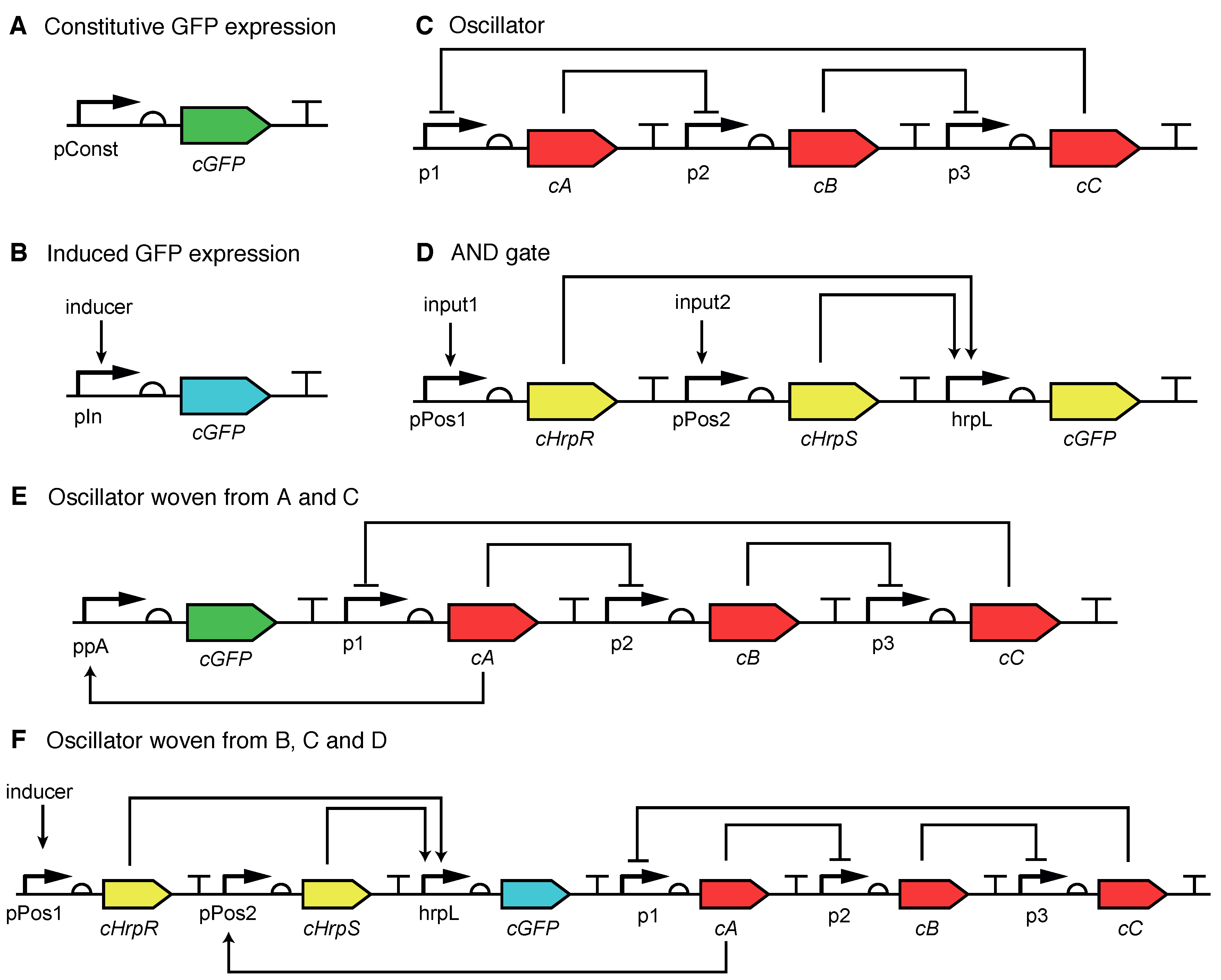
2.5. Designs for Switchable Oscillating Systems Using Concerns at the Part and System Levels
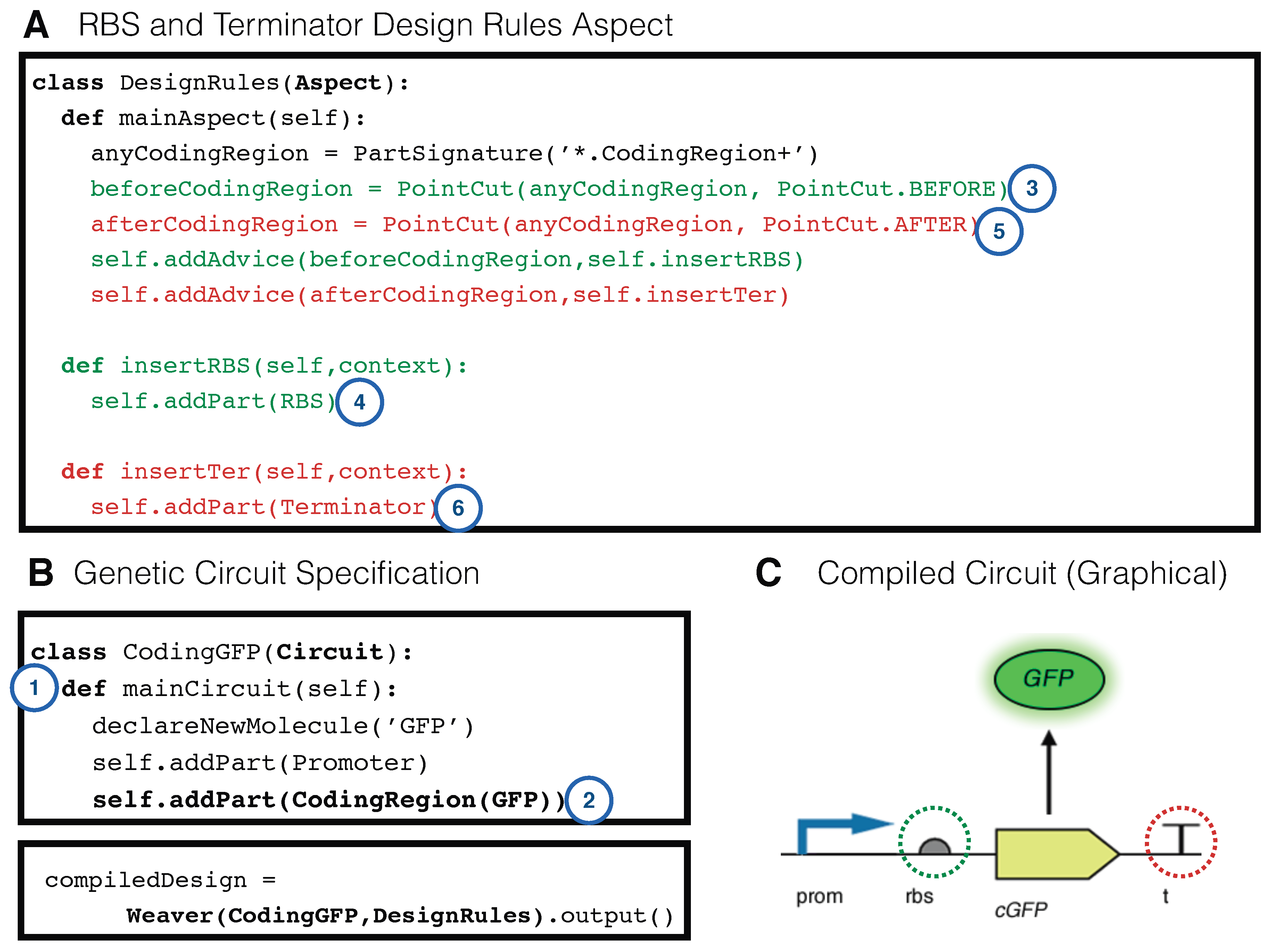
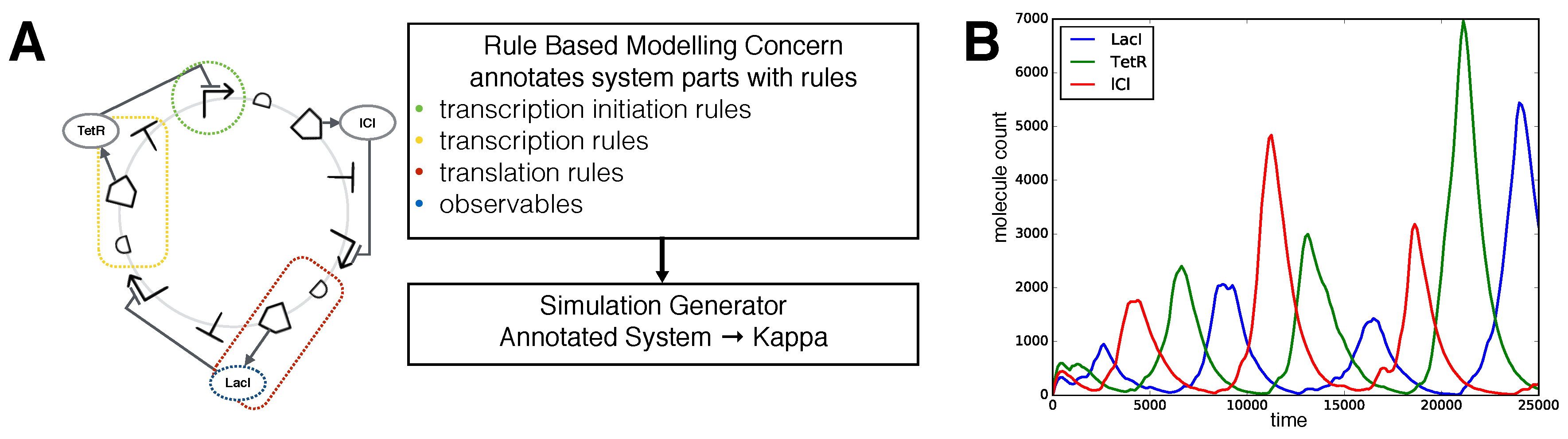
2.6. Rule-Based Modelling as an Aspect
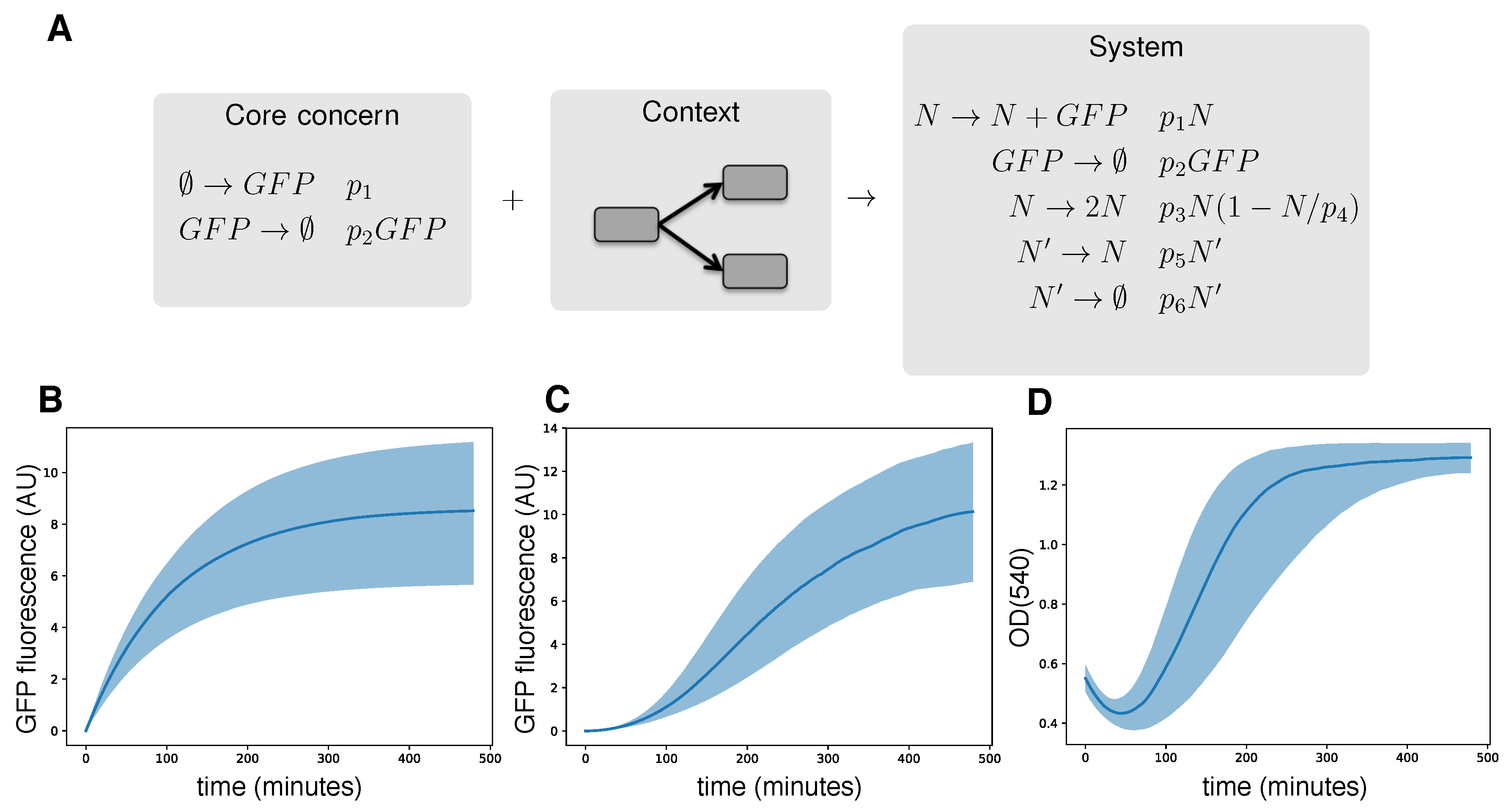
2.7. Type Advice, Abstraction and Cross-Cutting Contextual Model Generation
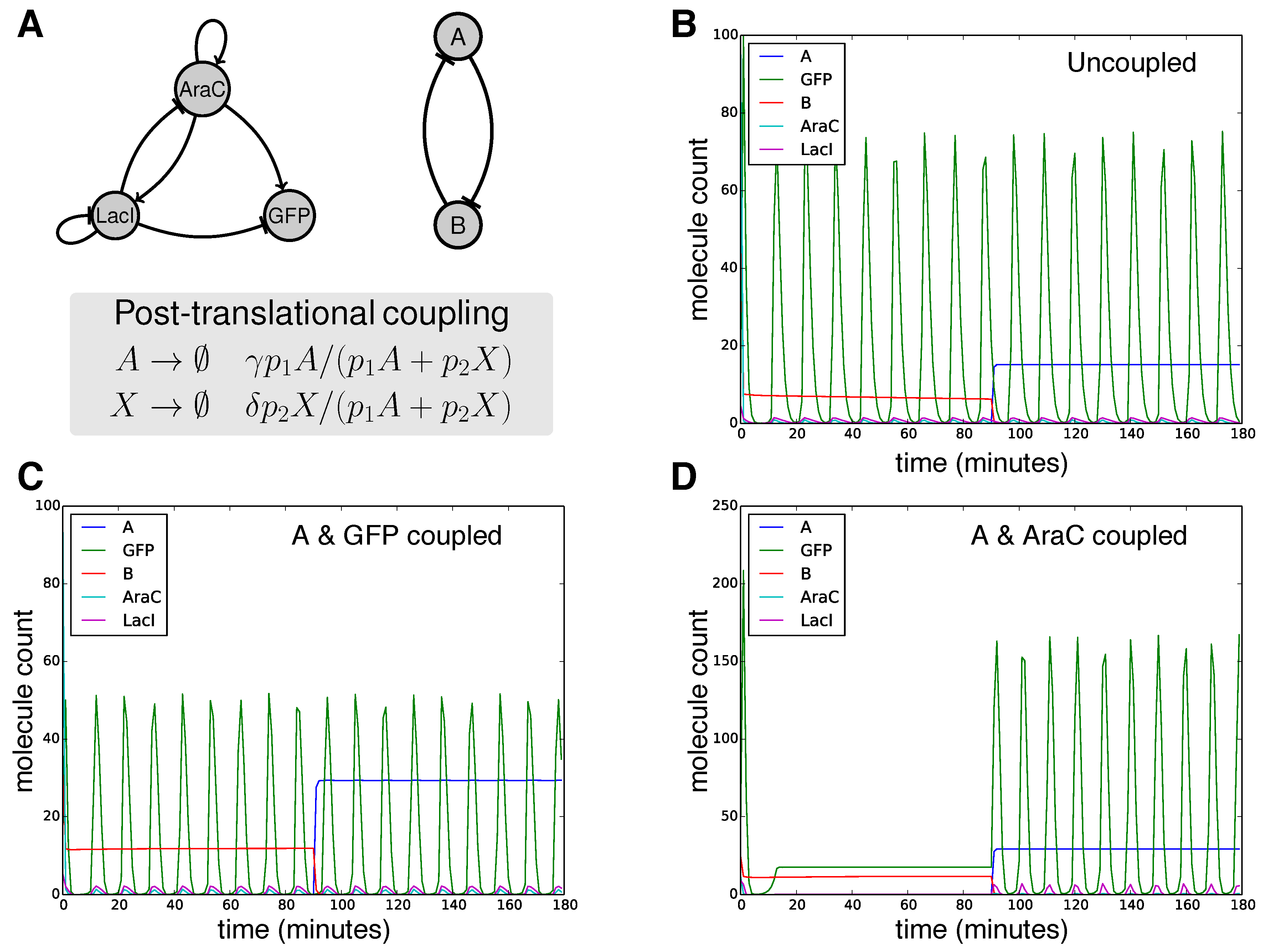
2.8. Designing with Core and Cross-Cutting Concerns: Post-Translational Coupling of a Bistable Switch and an Oscillator
3. Discussion
4. Materials and Methods
- SynBioWeaver is implemented in the Python package synbioweaver, licensed under the MIT licence and available on GitHub: https://github.com/ucl-cssb/synbioweaver
- The documentation is accessible at: http://synbioweaver.readthedocs.org
- The functionality described here is implemented within the examples of the package. Rule-based modelling requires that PySB and KaSim are installed. Biochemical network simulation on GPUs requires CUDA, PyCUDA [84] and cuda-sim. Bayesian inference requires ABC-SysBio.
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Endy, D. Foundations for engineering biology. Nature 2005, 438, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Canton, B.; Labno, A.; Endy, D. Refinement and standardization of synthetic biological parts and devices. Nat. Biotechnol. 2008, 26, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Arkin, A.P. A wise consistency: Engineering biology for conformity, reliability, predictability. Curr. Opin. Chem. Biol. 2013, 17, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, A.A.; Segall-Shapiro, T.H.; Voigt, C.A. Advances in genetic circuit design: Novel biochemistries, deep part mining, and precision gene expression. Curr. Opin. Chem. Biol. 2013, 17, 878–892. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, S.; Arkin, A.P. Contextualizing context for synthetic biology–identifying causes of failure of synthetic biological systems. Biotechnol. J. 2012, 7, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.S.; Surette, M.G.; Elowitz, M.B. Programming gene expression with combinatorial promoters. Mol. Syst. Biol. 2007, 3, 145. [Google Scholar] [CrossRef] [PubMed]
- Salis, H.M.; Mirsky, E.A.; Voigt, C.A. Automated design of synthetic ribosome binding sites to control protein expression. Nat. Biotechnol. 2009, 27, 946–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutalik, V.K.; Guimaraes, J.C.; Cambray, G.; Lam, C.; Christoffersen, M.J.; Mai, Q.A.; Tran, A.B.; Paull, M.; Keasling, J.D.; Arkin, A.P.; et al. Precise and reliable gene expression via standard transcription and translation initiation elements. Nat. Methods 2013, 10, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Mutalik, V.K.; Guimaraes, J.C.; Cambray, G.; Mai, Q.A.; Christoffersen, M.J.; Martin, L.; Yu, A.; Lam, C.; Rodriguez, C.; Bennett, G.; et al. Quantitative estimation of activity and quality for collections of functional genetic elements. Nat. Methods 2013, 10, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, D.; Ninfa, A.J.; Sontag, E.D. Modular cell biology: Retroactivity and insulation. Mol. Syst. Biol. 2008, 4, 161. [Google Scholar] [CrossRef] [PubMed]
- Jayanthi, S.; Nilgiriwala, K.S.; Del Vecchio, D. Retroactivity Controls the Temporal Dynamics of Gene Transcription. ACS Synth. Biol. 2013, 2, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Elf, J. Selective Charging of tRNA Isoacceptors Explains Patterns of Codon Usage. Science 2003, 300, 1718–1722. [Google Scholar] [CrossRef] [PubMed]
- Carrera, J.; Rodrigo, G.; Singh, V.; Kirov, B.; Jaramillo, A. Empirical model and in vivo characterization of the bacterial response to synthetic gene expression show that ribosome allocation limits growth rate. Biotechnol. J. 2011, 6, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Mather, W.H.; Hasty, J.; Tsimring, L.S.; Williams, R.J. Translational cross talk in gene networks. Biophys. J. 2013, 104, 2564–2572. [Google Scholar] [CrossRef] [PubMed]
- Gyorgy, A.; Jiménez, J.I.; Yazbek, J.; Huang, H.H.; Chung, H.; Weiss, R.; Del Vecchio, D. Isocost lines describe the cellular economy of genetic circuits. Biophys. J. 2015, 109, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Gorochowski, T.E.; Avcilar-Kucukgoze, I.; Bovenberg, R.A.; Roubos, J.A.; Ignatova, Z. A minimal model of ribosome allocation dynamics captures trade-offs in expression between endogenous and synthetic genes. ACS Synth. Biol. 2016, 5, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Ceroni, F.; Algar, R.; Stan, G.B.; Ellis, T. Quantifying cellular capacity identifies gene expression designs with reduced burden. Nat. Methods 2015, 12, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Cookson, N.A.; Mather, W.H.; Danino, T.; Mondragón-Palomino, O.; Williams, R.J.; Tsimring, L.S.; Hasty, J. Queueing up for enzymatic processing: Correlated signaling through coupled degradation. Mol. Syst. Biol. 2011, 7, 561. [Google Scholar] [CrossRef] [PubMed]
- Klumpp, S.; Zhang, Z.; Hwa, T. Growth Rate-Dependent Global Effects on Gene Expression in Bacteria. Cell 2009, 139, 1366–1375. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.; Gunderson, C.W.; Mateescu, E.M.; Zhang, Z.; Hwa, T. Interdependence of cell growth and gene expression: Origins and consequences. Science 2010, 330, 1099–1102. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, S.; Joachimiak, M.P.; Arkin, A.P. Effects of genetic variation on the E. coli host-circuit interface. Cell Rep. 2013, 4, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kitney, R.I.; Joly, N.; Buck, M. Engineering modular and orthogonal genetic logic gates for robust digital-like synthetic biology. Nat. Commun. 2011, 2, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stricker, J.; Cookson, S.; Bennett, M.R.; Mather, W.H.; Tsimring, L.S.; Hasty, J. A fast, robust and tunable synthetic gene oscillator. Nature 2008, 456, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Purcell, O.; Grierson, C.S.; Di Bernardo, M.; Savery, N.J. Temperature dependence of ssrA-tag mediated protein degradation. J. Biol. Eng. 2012, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Hussain, F.; Gupta, C.; Hirning, A.J.; Ott, W.; Matthews, K.S.; Josic, K.; Bennett, M.R. Engineered temperature compensation in a synthetic genetic clock. Proc. Natl. Acad. Sci. USA 2014, 111, 972–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, L.; Cox, R.S.; Weiss, R.; Arnold, F.H. Programmed population control by cell-cell communication and regulated killing. Nature 2004, 428, 868–871. [Google Scholar] [CrossRef] [PubMed]
- Moser, F.; Broers, N.J.; Hartmans, S.; Tamsir, A.; Kerkman, R.; Roubos, J.A.; Bovenberg, R.; Voigt, C.A. Genetic circuit performance under conditions relevant for industrial bioreactors. ACS Synth. Biol. 2012, 1, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Gorochowski, T.E.; Van Den Berg, E.; Kerkman, R.; Roubos, J.A.; Bovenberg, R.A. Using synthetic biological parts and microbioreactors to explore the protein expression characteristics of Escherichia coli. ACS Synth. Biol. 2013, 3, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Sleight, S.C.; Bartley, B.A.; Lieviant, J.A.; Sauro, H.M. Designing and engineering evolutionary robust genetic circuits. J. Biol. Eng. 2010, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.J.; Liu, P.; Nielsen, A.A.K.; Brophy, J.A.N.; Clancy, K.; Peterson, T.; Voigt, C.A. Characterization of 582 natural and synthetic terminators and quantification of their design constraints. Nat. Methods 2013, 10, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Sleight, S.C.; Sauro, H.M. Visualization of Evolutionary Stability Dynamics and Competitive Fitness of Escherichia coli Engineered with Randomized Multigene Circuits. ACS Synth. Biol. 2013, 2, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Rodriguez, J.; Yang, L.; Gorochowski, T.E.; Gordon, D.B.; Voigt, C.A. Memory and combinatorial logic based on DNA inversions: Dynamics and evolutionary stability. ACS Synth. Biol. 2015, 4, 1361–1372. [Google Scholar] [CrossRef] [PubMed]
- Rugbjerg, P.; Myling-Petersen, N.; Porse, A.; Sarup-Lytzen, K.; Sommer, M.O. Diverse genetic error modes constrain large-scale bio-based production. Nat. Commun. 2018, 9, 787. [Google Scholar] [CrossRef] [PubMed]
- Ceroni, F.; Boo, A.; Furini, S.; Gorochowski, T.E.; Borkowski, O.; Ladak, Y.N.; Awan, A.R.; Gilbert, C.; Stan, G.B.; Ellis, T. Burden-driven feedback control of gene expression. Nat. Methods 2018, 15, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Marguet, P.; You, L. Emergent bistability by a growth-modulating positive feedback circuit. Nat. Chem. Biol. 2009, 5, 842–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prindle, A.; Selimkhanov, J.; Li, H.; Razinkov, I.; Tsimring, L.S.; Hasty, J. Rapid and tunable post-translational coupling of genetic circuits. Nature 2014, 508, 387–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrell, J.; Rose, A. Temperature effects on microorganisms. Ann. Rev. Microbiol. 1967, 21, 101–120. [Google Scholar] [CrossRef] [PubMed]
- Laddad, R. AspectJ in Action: Enterprise AOP with Spring Applications; Manning Publications Co.: Shelter Island, NY, USA, 2009. [Google Scholar]
- Cai, Y.; Hartnett, B.; Gustafsson, C.; Peccoud, J. A syntactic model to design and verify synthetic genetic constructs derived from standard biological parts. Bioinformatics 2007, 23, 2760–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, M.; Phillips, A. Towards programming languages for genetic engineering of living cells. J. R. Soc. Interface 2009, 6 (Suppl. 4), S437–S450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.P.; Bergmann, F.T.; Chandran, D.; Sauro, H.M. Antimony: A modular model definition language. Bioinformatics 2009, 25, 2452–2454. [Google Scholar] [CrossRef] [PubMed]
- Mirschel, S.; Steinmetz, K.; Rempel, M.; Ginkel, M.; Gilles, E.D. PROMOT: Modular modeling for systems biology. Bioinformatics 2009, 25, 687–689. [Google Scholar] [CrossRef] [PubMed]
- Chandran, D.; Sauro, H.M. Hierarchical Modeling for Synthetic Biology. ACS Synth. Biol. 2012, 1, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.F.; Muhlich, J.L.; Bachman, J.A.; Sorger, P.K. Programming biological models in Python using PySB. Mol. Syst. Biol. 2013, 9, 646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liepe, J.; Barnes, C.; Cule, E.; Erguler, K.; Kirk, P.; Toni, T.; Stumpf, M.P.H. ABC-SysBio—Approximate Bayesian computation in Python with GPU support. Bioinformatics 2010, 26, 1797–1799. [Google Scholar] [CrossRef] [PubMed]
- Liepe, J.; Kirk, P.; Filippi, S.; Toni, T.; Barnes, C.P.; Stumpf, M.P.H. A framework for parameter estimation and model selection from experimental data in systems biology using approximate Bayesian computation. Nat. Protoc. 2014, 9, 439–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Liepe, J.; Sheng, X.; Stumpf, M.P.H.; Barnes, C. GPU accelerated biochemical network simulation. Bioinformatics 2011, 27, 874–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hucka, M.; Finney, A.; Sauro, H.M.; Bolouri, H.; Doyle, J.C.; Kitano, H.; Arkin, A.P.; Bornstein, B.J.; Bray, D.; Cornish-Bowden, A.; et al. The systems biology markup language (SBML): A medium for representation and exchange of biochemical network models. Bioinformatics 2003, 19, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Hürsch, W.L.; Lopes, C.V. Separation of Concerns; Northeastern University: Boston, MA, USA, 1995. [Google Scholar]
- Nielsen, A.A.; Der, B.S.; Shin, J.; Vaidyanathan, P.; Paralanov, V.; Strychalski, E.A.; Ross, D.; Densmore, D.; Voigt, C.A. Genetic circuit design automation. Science 2016, 352, aac7341. [Google Scholar] [CrossRef] [PubMed]
- Karr, J.R.; Sanghvi, J.C.; Macklin, D.N.; Gutschow, M.V.; Jacobs, J.M.; Bolival, B.; Assad-Garcia, N.; Glass, J.I.; Covert, M.W. A whole-cell computational model predicts phenotype from genotype. Cell 2012, 150, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Purcell, O.; Jain, B.; Karr, J.R.; Covert, M.W.; Lu, T.K. Towards a whole-cell modeling approach for synthetic biology. Chaos 2013, 23, 025112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S.; Densmore, D. Pigeon: A Design Visualizer for Synthetic Biology. ACS Synth. Biol. 2013, 2, 348–350. [Google Scholar] [CrossRef] [PubMed]
- Elowitz, M.B.; Leibler, S. A synthetic oscillatory network of transcriptional regulators. Nature 2000, 403, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Danos, V.; Laneve, C. Formal molecular biology. Theor. Comput. Sci. 2004, 325, 69–110. [Google Scholar] [CrossRef]
- Danos, V.; Feret, J.; Fontana, W.; Krivine, J. Scalable Simulation of Cellular Signaling Networks. In Asian Symposium on Programming Languages and Systems; Springer Berlin Heidelberg: Berlin/Heidelberg, Germany, 2007; pp. 139–157. [Google Scholar] [CrossRef]
- Ackers, G.K.; Johnson, A.D.; Shea, M.A. Quantitative model for gene regulation by lambda phage repressor. Proc. Natl. Acad. Sci. USA 1982, 79, 1129–1133. [Google Scholar] [CrossRef] [PubMed]
- Bornstein, B.J.; Keating, S.M.; Jouraku, A.; Hucka, M. LibSBML: An API library for SBML. Bioinformatics 2008, 24, 880–881. [Google Scholar] [CrossRef] [PubMed]
- Yates, G.T.; Smotzer, T. On the lag phase and initial decline of microbial growth curves. J. Theor. Biol. 2007, 244, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Yordanov, B.; Dalchau, N.; Grant, P.K.; Pedersen, M.; Emmott, S.; Haseloff, J.; Phillips, A. A Computational Method for Automated Characterization of Genetic Components. ACS Synth. Biol. 2014, 3, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Catanach, T.; McCardell, R.D.; Baetica, A.A.; Murray, R. Context Dependence of Biological Circuits. bioRxiv 2018. [Google Scholar] [CrossRef]
- Gardner, T.S.; Cantor, C.R.; Collins, J.J. Construction of a genetic toggle switch in Escherichia coli. Nature 2000, 403, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Litcofsky, K.D.; Afeyan, R.B.; Krom, R.J.; Khalil, A.S.; Collins, J.J. Iterative plug-and-play methodology for constructing and modifying synthetic gene networks. Nat. Methods 2012, 9, 1077–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, C.P.; Silk, D.; Sheng, X.; Stumpf, M.P.H. Bayesian design of synthetic biological systems. Proc. Natl. Acad. Sci. USA 2011, 108, 15190–15195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, C.P.; Silk, D.; Stumpf, M.P.H. Bayesian design strategies for synthetic biology. Interface Focus 2011, 1, 895–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilitchenko, L.; Liu, A.; Cheung, S.; Weeding, E.; Xia, B.; Leguia, M.; Anderson, J.C.; Densmore, D. Eugene—A domain specific language for specifying and constraining synthetic biological parts, devices, and systems. PLoS ONE 2011, 6, e18882. [Google Scholar] [CrossRef] [PubMed]
- Beal, J.; Lu, T.; Weiss, R. Automatic Compilation from High-Level Biologically-Oriented Programming Language to Genetic Regulatory Networks. PLoS ONE 2011, 6, e22490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beal, J.; Weiss, R.; Densmore, D.; Adler, A.; Appleton, E.; Babb, J.; Bhatia, S.; Davidsohn, N.; Haddock, T.; Loyall, J.; et al. An End-to-End Workflow for Engineering of Biological Networks from High-Level Specifications. ACS Synth. Biol. 2012, 1, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Dalchau, N.; Szép, G.; Hernansaiz-Ballesteros, R.; Barnes, C.P.; Cardelli, L.; Phillips, A.; Csikász-Nagy, A. Computing with biological switches and clocks. Nat. Comput. 2018. [Google Scholar] [CrossRef]
- Der, B.S.; Glassey, E.; Bartley, B.A.; Enghuus, C.; Goodman, D.B.; Gordon, D.B.; Voigt, C.A.; Gorochowski, T.E. DNAplotlib: Programmable visualization of genetic designs and associated data. ACS Synth. Biol. 2016, 6, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Galdzicki, M.; Clancy, K.P.; Oberortner, E.; Pocock, M.; Quinn, J.Y.; Rodriguez, C.A.; Roehner, N.; Wilson, M.L.; Adam, L.; Anderson, J.C.; et al. The Synthetic Biology Open Language (SBOL) provides a community standard for communicating designs in synthetic biology. Nat. Biotechnol. 2014, 32, 545–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- François, P.; Hakim, V. Design of genetic networks with specified functions by evolution in silico. Proc. Natl. Acad. Sci. USA 2004, 101, 580–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigo, G.; Carrera, J.; Jaramillo, A. Computational design of synthetic regulatory networks from a genetic library to characterize the designability of dynamical behaviors. Nucleic Acids Res. 2011, 39, e138. [Google Scholar] [CrossRef] [PubMed]
- Huynh, L.; Tagkopoulos, I. Fast and accurate circuit design automation through hierarchical model switching. ACS Synth. Biol. 2015, 4, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Woods, M.L.; Leon, M.; Perez-Carrasco, R.; Barnes, C.P. A statistical approach reveals designs for the most robust stochastic gene oscillators. ACS Synth. Biol. 2016, 5, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, T.; Fedorec, A.J.; Danino, T.; Barnes, C.P. Synthetic Biology and Engineered Live Biotherapeutics: Toward Increasing System Complexity. Cell Syst. 2018, 7, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.R.; Rubin, A.J.; Davis, J.H.; Ajo-Franklin, C.M.; Cumbers, J.; Czar, M.J.; de Mora, K.; Glieberman, A.L.; Monie, D.D.; Endy, D. Measuring the activity of BioBrick promoters using an in vivo reference standard. J. Biol. Eng. 2009, 3, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beal, J.; Haddock-Angelli, T.; Baldwin, G.; Gershater, M.; Dwijayanti, A.; Storch, M.; de Mora, K.; Lizarazo, M.; Rettberg, R. Quantification of bacterial fluorescence using independent calibrants. PLoS ONE 2018, 13, e0199432. [Google Scholar] [CrossRef] [PubMed]
- Beal, J.; Haddock-Angelli, T.; Farny, N.; Rettberg, R. Time to Get Serious about Measurement in Synthetic Biology. Trends Biotechnol. 2018, 36, 869–871. [Google Scholar] [CrossRef] [PubMed]
- Gorochowski, T.E.; Borujeni, A.E.; Park, Y.; Nielsen, A.A.; Zhang, J.; Der, B.S.; Gordon, D.B.; Voigt, C.A. Genetic circuit characterization and debugging using RNA-seq. Mol. Syst. Biol. 2017, 13, 952. [Google Scholar] [CrossRef] [PubMed]
- Geertz, M.; Shore, D.; Maerkl, S.J. Massively parallel measurements of molecular interaction kinetics on a microfluidic platform. Proc. Natl. Acad. Sci. USA 2012, 9, 16540–16545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braniff, N.; Ingalls, B. New Opportunities for Optimal Design of Dynamic Experiments in Systems and Synthetic Biology. Curr. Opin. Syst. Biol. 2018, 9, 42–48. [Google Scholar] [CrossRef]
- Subsoontorn, P.; Kim, J.; Winfree, E. Ensemble Bayesian analysis of bistability in a synthetic transcriptional switch. ACS Synth. Biol. 2012, 1, 299–316. [Google Scholar] [CrossRef] [PubMed]
- Klöckner, A.; Pinto, N.; Lee, Y.; Catanzaro, B.; Ivanov, P.; Fasih, A. PyCUDA and PyOpenCL: A Scripting-Based Approach to GPU Run-Time Code Generation. Parallel Comput. 2012, 38, 157–174. [Google Scholar] [CrossRef]










© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boeing, P.; Leon, M.; Nesbeth, D.N.; Finkelstein, A.; Barnes, C.P. Towards an Aspect-Oriented Design and Modelling Framework for Synthetic Biology. Processes 2018, 6, 167. https://doi.org/10.3390/pr6090167
Boeing P, Leon M, Nesbeth DN, Finkelstein A, Barnes CP. Towards an Aspect-Oriented Design and Modelling Framework for Synthetic Biology. Processes. 2018; 6(9):167. https://doi.org/10.3390/pr6090167
Chicago/Turabian StyleBoeing, Philipp, Miriam Leon, Darren N. Nesbeth, Anthony Finkelstein, and Chris P. Barnes. 2018. "Towards an Aspect-Oriented Design and Modelling Framework for Synthetic Biology" Processes 6, no. 9: 167. https://doi.org/10.3390/pr6090167