


Biological Methanation in an Anaerobic Biofilm Reactor—Trace Element and Mineral Requirements for Stable Operation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Lab Scale Experiments
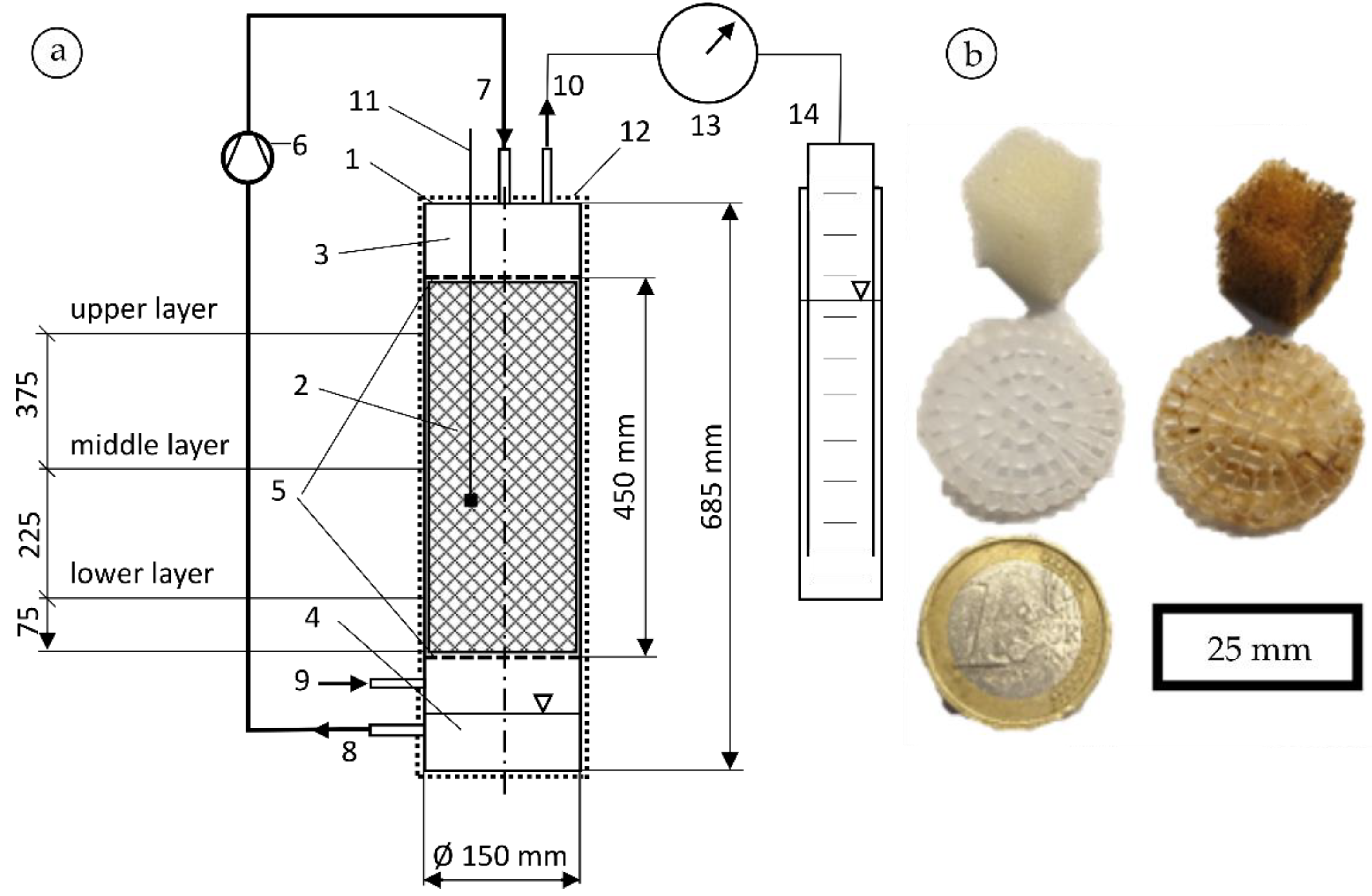
2.1.1. Biofilm Reactor Setup
2.1.2. Trickling Liquid and Trace Element Dosing
2.1.3. Biomass Sampling
2.2. Analytical Methods
2.2.1. Chemical Standard Analysis
2.2.2. Dry Matter- and Organic Dry Matter Concentration of the Biofilm Carriers
2.2.3. Biomass and Trickling Liquid Sample Revealing and Nutrient Analysis (ICP-OES)
2.2.4. Microscopic Imaging and Amplicon Sequencing (16S rRNA Gene)
3. Results and Discussion
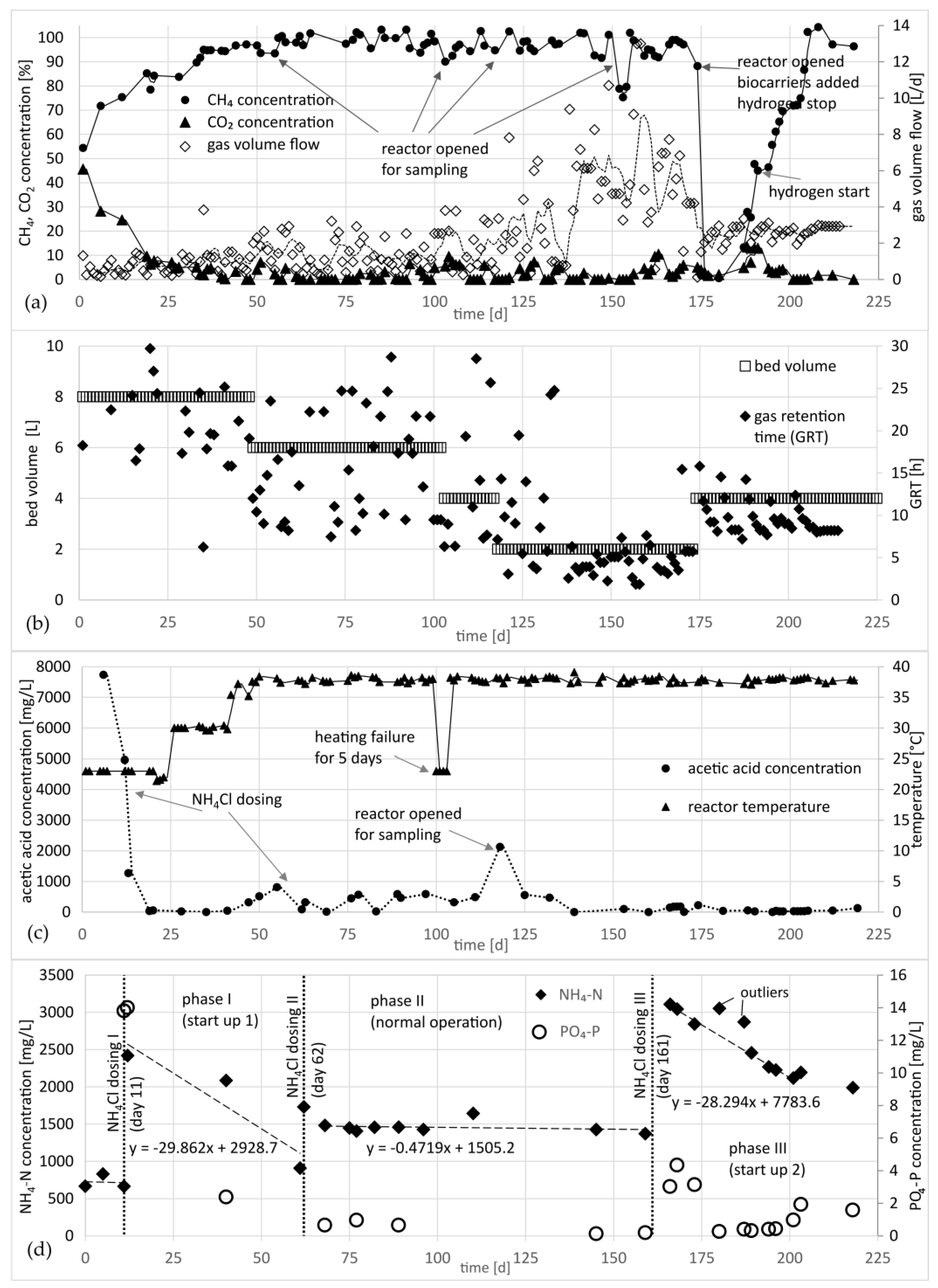
3.1. Raw Gas Composition
3.2. Product Gas-Flow and -Composition, Reactor Temperature, Ammonium Concentration, Nitrogen Demand, and Acetic Acid Concentration in the Biofilm Reactor
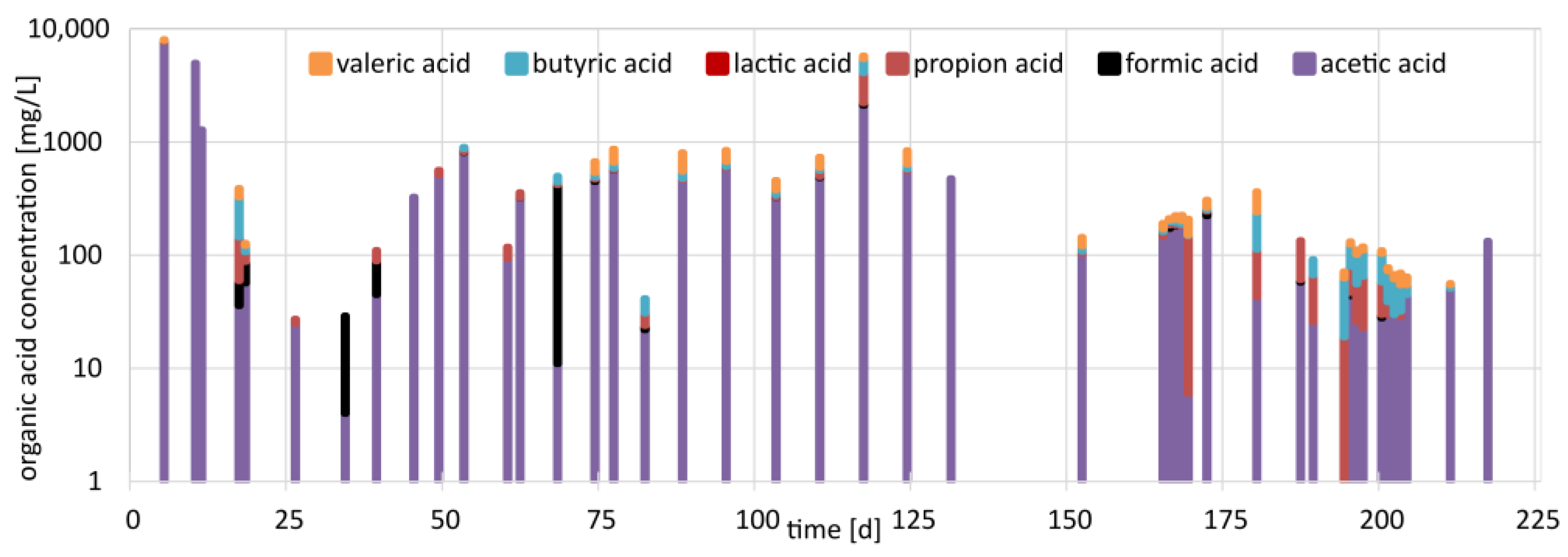
3.3. Organic Acid Concentrations in the Trickling Liquid
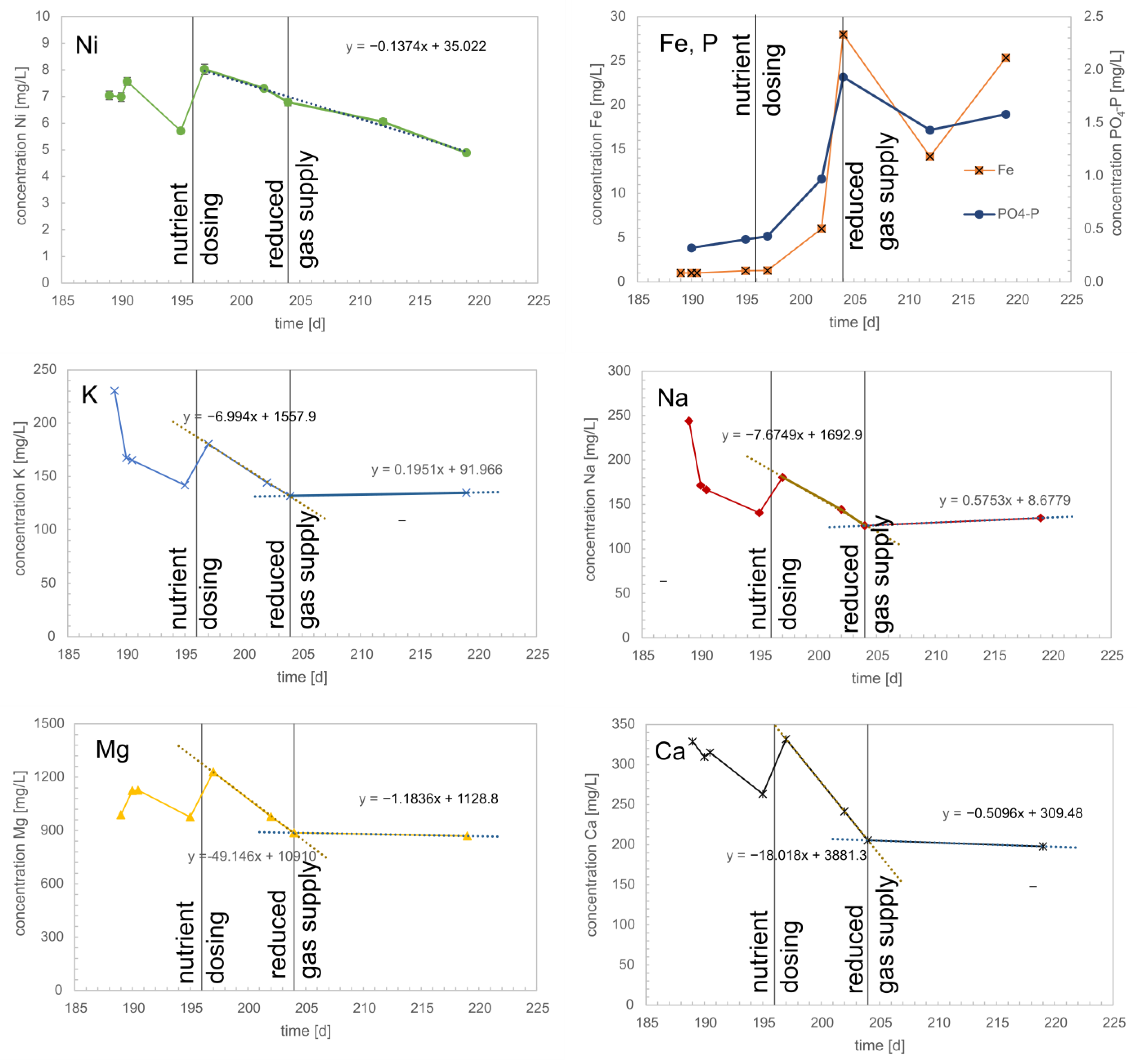
3.4. Trace Element Concentration in the Trickling Liquid and Biomass
3.5. Nutrient Demand of the Biomass
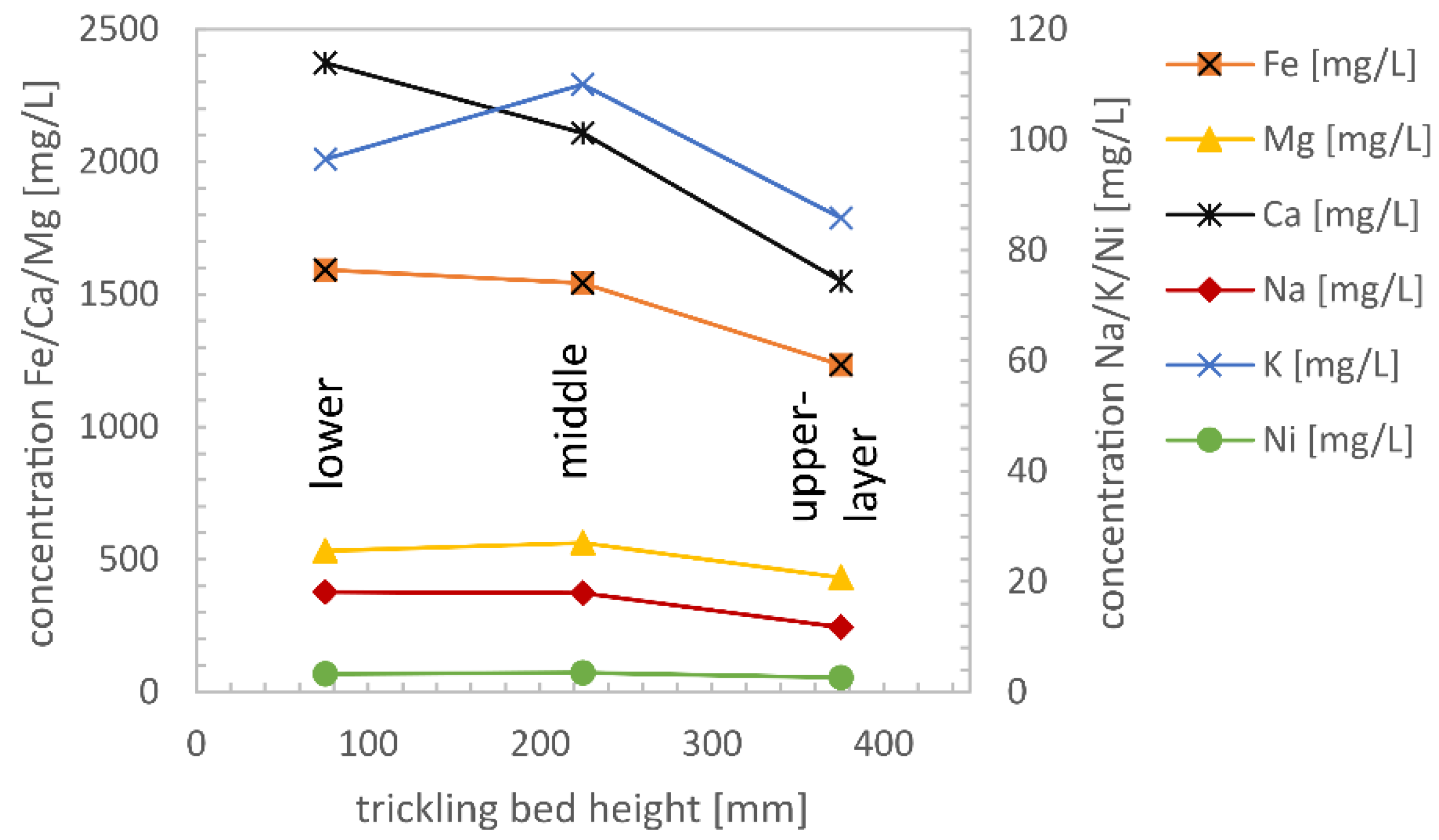
3.6. Trace Element Concentration in the Biomass according to Sampling Position and Time
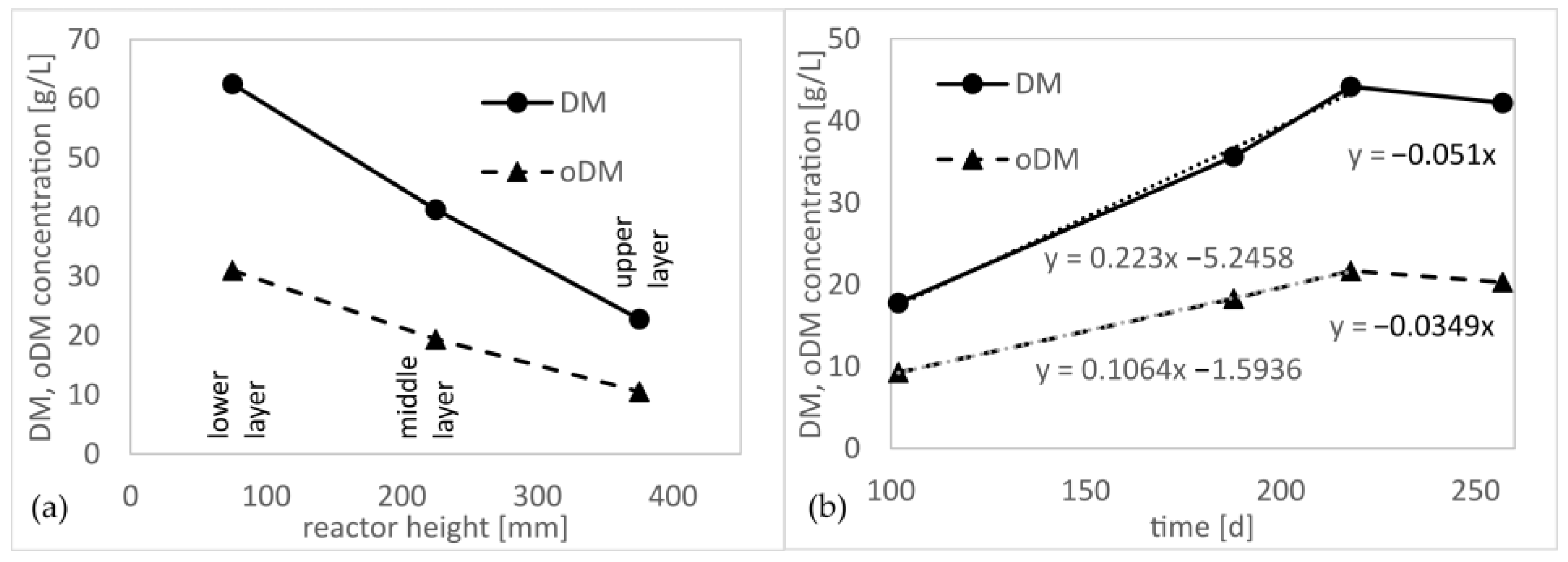
3.7. Biomass Concentration in the Trickling Bed
3.8. Microscopy Imaging
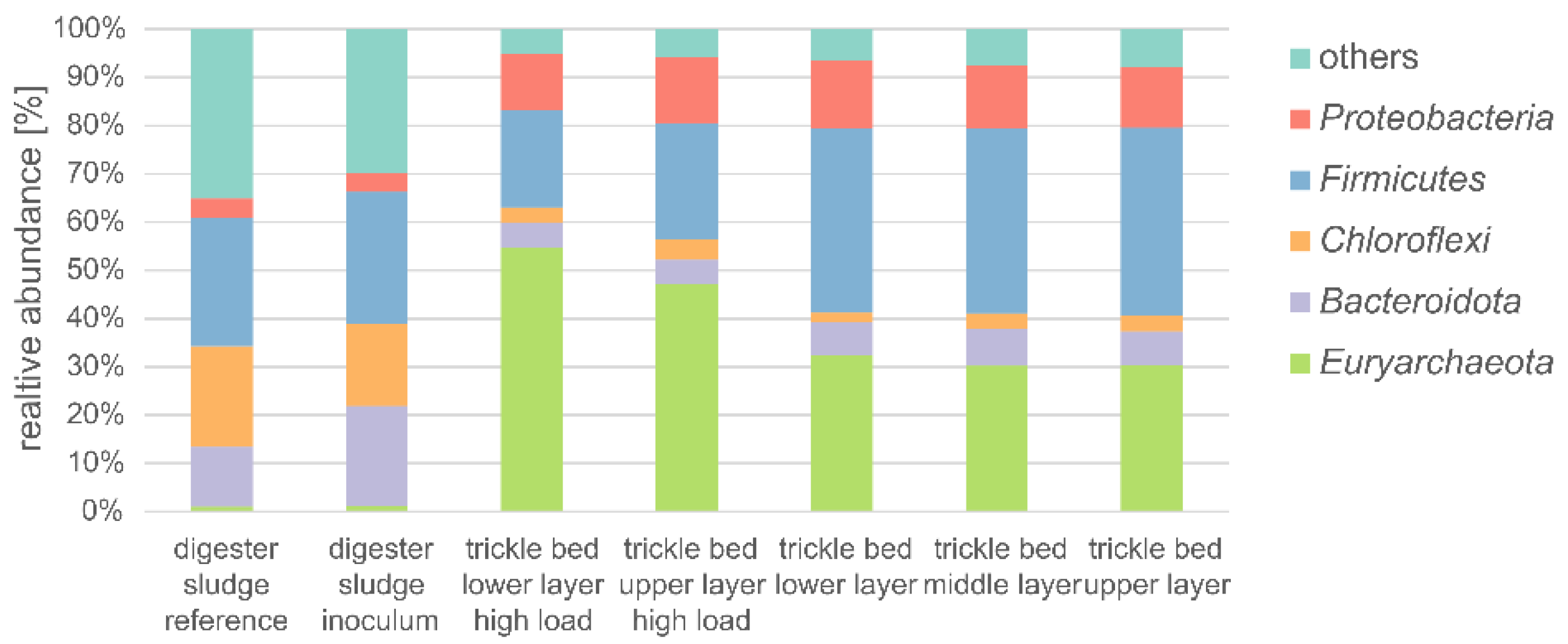
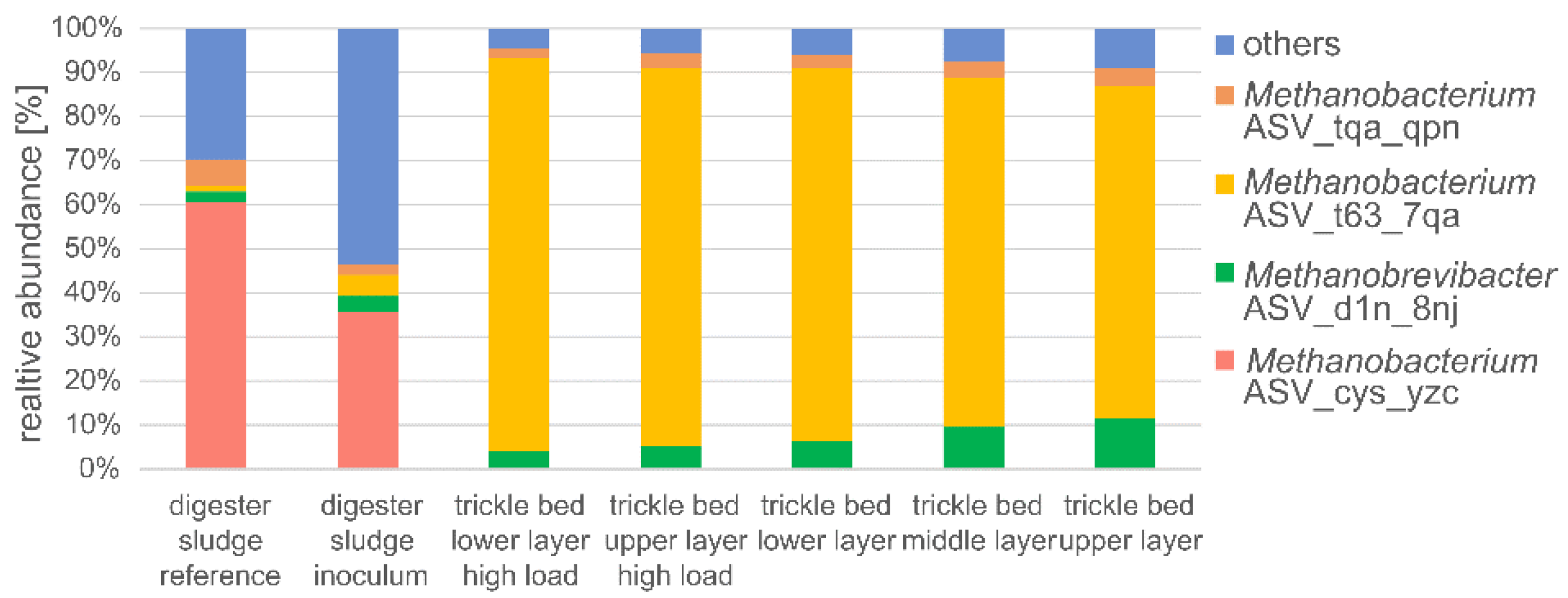
3.9. Microbial Community Dynamics of the Biomass
4. Conclusions
- (1)
- An anaerobic trickling bed reactor was operated at mesophilic conditions for 225 days. The biological methanation of digester gas produced from municipal raw sludge was demonstrated under realistic conditions;
- (2)
- Within 35 days after start-up, the methane concentration in the product gas reached 95% (65% in the raw gas);
- (3)
- Two different biofilm carrier materials were tested, PU-foam cubes and structured packing media made of PE-hard plastic. Microscopic images showed a biofilm with a thickness of approximately 5–10 µm on both carrier types, whereby, depending on the gas load, more biomass growth occurs at the raw gas inlet in the lower part of the reactor;
- (4)
- To promote the hydrogenotrophic pathway, NH4Cl was dosed to hold the NH4-N concentration in the trickling liquid between 1500 mg/L and 3000 mg/L. At ammonium levels below 1000 mg/L, the acetic acid concentration increased, and it is assumed that the acetoclastic methanogens became dominant;
- (5)
- The gas load was increased gradually, and stable methanation with methane concentration >95% was demonstrated at gas retention times between 2.5 h and 5 h;
- (6)
- After adding additional biofilm carriers and two weeks of an operational pause without gas supply at mesophilic conditions, 21 days after starting the raw gas and hydrogen supply again, the methane concentration reached 95% again;
- (7)
- Tests for trace elements and minerals (Na, K, Ca, Mg, Ni, Fe, and P) were used to determine biomass-specific uptake rates during the biological methanation process. Some trace elements such as Ni and Fe are required at low concentrations for the formation of hydrogenase enzymes independent of reactor load, some minerals are required load dependent (Na, K, Ca, and Mg), and for some elements, the concentration is strongly depending on the oxidation state (Fe, P). Remarkable is the load-dependent demand of the alkali (Na, K) and alkaline earth metals (Mg, Ca), which are needed in the metabolism of the archaea. For iron and phosphorus, a correlation between the oxidation state (FeII or FeIII) after opening the reactor for biomass sampling is assumed.
- (8)
- For Na, K, Ca, Mg, and Ni biomass specific uptake rates were calculated, and a monthly dosage recommendation was given;
- (9)
- Although microscopic examination of the biofilm did not reveal substantial differences between the different loads of the rector, the abundance and composition of the hydrogenotrophic biomass in the biofilm on the growing media changed dramatically. At high loadings, Methanobacterium sp. strongly dominated the microbial community with a relative abundance of up to 49%, while it was very rare in the inoculated sludge and the reference sludge with <0.05%.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Angelidaki, I.; Petersen, S.P.; Ahring, B.K. Effects of lipids on thermophilic anaerobic digestion and reduction of lipid inhibition upon addition of bentonite. Appl. Microb. Biotechnol. 1990, 33, 469–472. [Google Scholar] [CrossRef]
- Angelidaki, I.; Treu, L.; Tsapekos, P.; Luo, G.; Campanaro, S.; Wenzel, H.; Kougias, P.G. Biogas upgrading and utilization: Current status and perspectives. Biotechnol. Adv. 2018, 36, 452–466. [Google Scholar] [CrossRef] [Green Version]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.T.; Yde, L.; Triolo, J.M.; Wenzel, H. Optimizing the dosing and trickling of nutrient media for thermophilic biomethanation in a biotrickling filter. Biochem. Eng. J. 2021, 176, 108220. [Google Scholar] [CrossRef]
- Banks, C.J.; Zhang, Y.; Jiang, Y.; Heaven, S. Trace element requirements for stable food waste digestion at elevated ammonia concentrations. Bioresour. Technol. 2012, 104, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Bassani, I.; Kougias, P.G.; Treu, L.; Angelidaki, I. Biogas Upgrading via Hydrogenotrophic Methanogenesis in Two-Stage Continuous Stirred Tank Reactors at Mesophilic and Thermophilic Conditions. Environ. Sci. Technol. 2015, 49, 12585–12593. [Google Scholar] [CrossRef]
- BMNT. Mission 2030—Die Österreichische Klima- und Energiestrategie (Mission 2030—The Austrian Climate and Energy Strategy); Federal Ministry Republic of Austria Sustainability and Tourism: Vienna, Austria, 2018; p. 80.
- BMNT. Integrierter Nationaler Energie- und Klimaplan für Österreich—Periode 2021–2030 (Integrated National Energy and Climate Plan for Austria—Period 2021–2030); Federal Ministry Republic of Austria Sustainability and Tourism: Vienna, Austria, 2019; p. 272.
- Burkhardt, M.; Busch, G. Methanation of hydrogen and carbon dioxide. Appl. Energy 2013, 111, 74–79. [Google Scholar] [CrossRef]
- Burkhardt, M.; Koschack, T.; Busch, G. Biocatalytic methanation of hydrogen and carbon dioxide in an anaerobic three-phase system. Bioresour. Technol. 2015, 178, 330–333. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; Sankaran, K.; Fukuyama, J.A.; McMurdie, P.J.; Holmes, S.P. Bioconductor Workflow for Microbiome Data Analysis: From raw reads to community analyses. F1000Res 2016, 5, 1492. [Google Scholar] [CrossRef] [PubMed]
- Can, M.; Armstrong, F.A.; Ragsdale, S.W. Structure, Function, and Mechanism of the Nickel Metalloenzymes, CO Dehydrogenase, and Acetyl-CoA Synthase. Chem. Rev. 2014, 114, 4149–4174. [Google Scholar] [CrossRef] [PubMed]
- Demirel, B.; Scherer, P. Trace element requirements of agricultural biogas digesters during biological conversion of renewable biomass to methane. Biomass Bioenergy 2011, 35, 992–998. [Google Scholar] [CrossRef]
- Deublein, D.; Steinhauser, A. Biogas from Waste and Renewable Resources: An Introduction; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Diaz, I.; Perez, C.; Alfaro, N.; Fdz-Polanco, F. A feasibility study on the bioconversion of CO2 and H2 to biomethane by gas sparging through polymeric membranes. Bioresour. Technol. 2015, 185, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Feickert Fenske, C.; Md, Y.; Strübing, D.; Koch, K. Preliminary gas flow experiments identify improved gas flow conditions in a pilot-scale trickle bed reactor for H2 and CO2 biological methanation. Bioresour. Technol. 2023, 371, 128648. [Google Scholar] [CrossRef]
- Fu, S.; Angelidaki, I.; Zhang, Y. In Situ Biogas Upgrading by CO2-to-CH4 Bioconversion. Trends Biotechnol. 2021, 39, 336–347. [Google Scholar] [CrossRef]
- Garcia, J.-L.; Patel, B.K.C.; Ollivier, B. Taxonomic, Phylogenetic, and Ecological Diversity of Methanogenic Archaea. Anaerobe 2000, 6, 205–226. [Google Scholar] [CrossRef]
- Götz, M.; Lefebvre, J.; Mörs, F.; McDaniel Koch, A.; Graf, F.; Bajohr, S.; Reimert, R.; Kolb, T. Renewable Power-to-Gas: A technological and economic review. Renew. Energy 2016, 85, 1371–1390. [Google Scholar] [CrossRef] [Green Version]
- Ju, D.-H.; Shin, J.-H.; Lee, H.-K.; Kong, S.-H.; Kim, J.-I.; Sang, B.-I. Effects of pH conditions on the biological conversion of carbon dioxide to methane in a hollow-fiber membrane biofilm reactor (Hf–MBfR). Desalination 2008, 234, 409–415. [Google Scholar] [CrossRef]
- Kim, S.; Choi, K.; Chung, J. Reduction in carbon dioxide and production of methane by biological reaction in the electronics industry. Int. J. Hydrogen Energy 2013, 38, 3488–3496. [Google Scholar] [CrossRef]
- Kleemann, A. Secific Aerea of Linpor Biocarriers; Strabag Water Technologies: Vienna, Austria, 2018. [Google Scholar]
- Kougias, P.G.; Treu, L.; Benavente, D.P.; Boe, K.; Campanaro, S.; Angelidaki, I. Ex-situ biogas upgrading and enhancement in different reactor systems. Bioresour. Technol. 2017, 225, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Lecker, B.; Illi, L.; Lemmer, A.; Oechsner, H. Biological hydrogen methanation—A review. Bioresour. Technol. 2017, 245 Pt A, 1220–1228. [Google Scholar] [CrossRef]
- Lee, J.C.; Kim, J.H.; Chang, W.S.; Pak, D. Biological conversion of CO2 to CH4 using hydrogenotrophic methanogen in a fixed bed reactor. J. Chem. Technol. Biotechnol. 2012, 87, 844–847. [Google Scholar] [CrossRef]
- Luo, G.; Angelidaki, I. Integrated biogas upgrading and hydrogen utilization in an anaerobic reactor containing enriched hydrogenotrophic methanogenic culture. Biotechnol. Bioeng. 2012, 109, 2729–2736. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Angelidaki, I. Hollow fiber membrane based H2 diffusion for efficient in situ biogas upgrading in an anaerobic reactor. Appl. Microb. Biotechnol. 2013, 97, 3739–3744. [Google Scholar] [CrossRef]
- Markthaler, S.; Plankenbühler, T.; Miederer, J.; Kolb, S.; Herkendell, K.; Karl, J. Combined Two-Model CFD Simulation of Trickle Bed Reactors with Head-Sump Extension: Case Study on Hydrodynamics and Biological Methanation. Ind. Eng. Chem. Res. 2022, 61, 4134–4152. [Google Scholar] [CrossRef]
- Mattiasson, B. Ekologisk Lunga for Biogasuppgradering. (Ecological Lungs for Biogas Upgrading). 2005. Available online: www.sgc.se/ckfinder/userfiles/files/SBGF610401.pdf (accessed on 4 January 2023).
- Maus, I.; Wibberg, D.; Stantscheff, R.; Cibis, K.; Eikmeyer, F.-G.; König, H.; Pühler, A.; Schlüter, A. Complete genome sequence of the hydrogenotrophic Archaeon Methanobacterium sp. Mb1 isolated from a production-scale biogas plant. J. Biotechnol. 2013, 168, 734–736. [Google Scholar] [CrossRef]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.L.; Agarwala, R.; Schäffer, A.A. Database indexing for production MegaBLAST searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, R.; Meier, L.; Diaz, I.; Jeison, D. A review on the state-of-the-art of physical/chemical and biological technologies for biogas upgrading. Rev. Environ. Sci. Bio/Technol. 2015, 14, 727–759. [Google Scholar] [CrossRef] [Green Version]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Pjevac, P.; Hausmann, B.; Schwarz, J.; Kohl, G.; Herbold, C.W.; Loy, A.; Berry, D. An Economical and Flexible Dual Barcoding, Two-Step PCR Approach for Highly Multiplexed Amplicon Sequencing. Front. Microbiol. 2021, 12, 669776. [Google Scholar] [CrossRef] [PubMed]
- Pratofiorito, G.; Hackbarth, M.; Mandel, C.; Madlanga, S.; West, S.; Horn, H.; Hille-Reichel, A. A membrane biofilm reactor for hydrogenotrophic methanation. Bioresour. Technol. 2021, 321, 124444. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Rachbauer, L.; Voitl, G.; Bochmann, G.; Fuchs, W. Biological biogas upgrading capacity of a hydrogenotrophic community in a trickle-bed reactor. Appl. Energy 2016, 180, 483–490. [Google Scholar] [CrossRef]
- Rittmann, S.; Seifert, A.; Herwig, C. Essential prerequisites for successful bioprocess development of biological CH4 production from CO2 and H2. Crit. Rev. Biotechnol. 2015, 35, 141–151. [Google Scholar] [CrossRef]
- Scherer, P.; Lippert, H.; Wolff, G. Composition of the major elements and trace elements of 10 methanogenic bacteria determined by inductively coupled plasma emission spectrometry. Biol. Trace Elem. Res. 1983, 5, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Stantscheff, R.; Kuever, J.; Rabenstein, A.; Seyfarth, K.; Dröge, S.; König, H. Isolation and differentiation of methanogenic Archaea from mesophilic corn-fed on-farm biogas plants with special emphasis on the genus Methanobacterium. Appl. Microbiol. Biotechnol. 2014, 98, 5719–5735. [Google Scholar] [CrossRef]
- Sterner, M.; Stadler, I. Energiespeicher—Bedarf, Technologien, Integration; Springer Vieweg: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Strevett, K.A.; Vieth, R.F.; Grasso, D. Chemo-autotrophic biogas purification for methane enrichment: Mechanism and kinetics. Chem. Eng. J. Biochem. Eng. J. 1995, 58, 71–79. [Google Scholar] [CrossRef]
- Strübing, D.; Huber, B.; Lebuhn, M.; Drewes, J.E.; Koch, K. High performance biological methanation in a thermophilic anaerobic trickle bed reactor. Bioresour. Technol. 2017, 245 Pt A, 1176–1183. [Google Scholar] [CrossRef]
- Tauber, J.; Parravicini, V.; Svardal, K.; Krampe, J. Quantifying methane emissions from anaerobic digesters. Water Sci. Technol. 2019, 80, 1654–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tauber, J.; Ramsbacher, A.; Svardal, K.; Krampe, J. Energetic Potential for Biological Methanation in Anaerobic Sewage Sludge Digesters in Austria. Energies 2021, 14, 6618. [Google Scholar] [CrossRef]
- Thauer, R.K. The Wolfe cycle comes full circle. Proc. Natl. Acad. Sci. USA 2012, 109, 15084–15085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thauer, R.K.; Kaster, A.-K.; Goenrich, M.; Schick, M.; Hiromoto, T.; Shima, S. Hydrogenases from Methanogenic Archaea, Nickel, a Novel Cofactor, and H2 Storage. Annu. Rev. Biochem. 2010, 79, 507–536. [Google Scholar] [CrossRef] [Green Version]
- Tsapekos, P.; Treu, L.; Campanaro, S.; Centurion, V.B.; Zhu, X.; Peprah, M.; Zhang, Z.; Kougias, P.G.; Angelidaki, I. Pilot-scale biomethanation in a trickle bed reactor: Process performance and microbiome functional reconstruction. Energy Convers. Manag. 2021, 244, 114491. [Google Scholar] [CrossRef]
- United Nations. Paris Agreement. 2015; p. 27. Available online: https://unfccc.int/sites/default/files/english_paris_agreement.pdf (accessed on 7 January 2023).
- Voelklein, M.A.; Rusmanis, D.; Murphy, J.D. Biological methanation: Strategies for in-situ and ex-situ upgrading in anaerobic digestion. Appl. Energy 2019, 235, 1061–1071. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, X.; Yan, Q.; Zhang, Y.; Angelidaki, I. Microbial community response to ammonia levels in hydrogen assisted biogas production and upgrading process. Bioresour. Technol. 2020, 296, 122276. [Google Scholar] [CrossRef]
- Xingfeng. Pingxiang City, China. Moving Bed MBBR Bio Filter Media. 2021. Available online: https://chemxing.en.made-in-china.com/product/DNUnyYibbXVQ/China-Moving-Bed-Mbbr-Bio-Filter-Media.html (accessed on 1 March 2021).
- Yuan, G. Stickstoffelimination unter Einsatz von Extern Gezüchteten Immobilisierten Nitrifikanten (Nitrogen Elimination with use of Externally Reared Immobolized Nitrifiers); Stuttgarter Berichte zur Siedlungswasserwirtschaft, Bd. 144; Kommissionsverlag, R.O., Ed.; Oldenbourg Wissenschaftsverlag: Munich, Germany, 1997. [Google Scholar]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, Z.; Ren, H.-Y.; Nan, J.; Chen, C.; Ren, N.-Q. Improving biogas upgrading and liquid chemicals production simultaneously by a membrane biofilm reactor. Bioresour. Technol. 2020, 313, 123693. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | H | N | Na | K | S | P | Ca (Order I) | Ca (Order II) |
|---|---|---|---|---|---|---|---|---|
| [% w/w] | [%] | [%] | [%] | [%] | [%] | [%] | [ppm] | [ppm] |
| 37–44 | 5.5–6.5 | 9.5–12.8 | 0.3–4.0 | 0.13–5.0 | 0.56–1.2 | 0.5–2.8 | 85–550 | 1000–4500 |
| Mg | Fe | Ni | Co | Mo | Zn | Cu | Mn | |
| [%] | [%] | [ppm] | [ppm] | [ppm] | [ppm] | [ppm] | [ppm] | |
| 0.09–0.53 | 0.07–0.28 | 65–180 | 10–120 | 10–70 | 50–630 | <10–160 | <5–25 |
| Reference Element X | Ju et al. [20] | Angelidaki et al. [38] | Strübing et al. [14] | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Compound | Element | Compound | Element | Compound | Element | |||||
| [Name] | [Formula] | [mg/L] | [mg X/L] | [Formula] | [mg/L] | [mg X/L] | [mg X/L] Reactor | [Formula] | [mg/L] Stock Solution | [mg X/L] Trickling Liquid |
| Membrane Reactor | Stock Solution | |||||||||
| Mg | MgCl2·6H2O | 16.05 | 1.919 | MgCl2·6H2O | 10,000 | 1195.512 | 12.274 | MgCl2·6H2O | 300 | 35.865 |
| Ca | CaCl2·2H2O | 1.2 | 0.327 | CaCl2·2H2O | 5000 | 1363.068 | 13.994 | - | - | - |
| Zn | ZnCl2 | 5.91 | 2.835 | ZnCl2 | 50 | 23.988 | 0.0246 | - | - | - |
| Mo | Na2Mo·2H2O | 1.29 | 0.695 | (NH4)6Mo7O24·4H2O | 50 | 27.171 | 0.0279 | (NH4)6Mo7O24·2H2O | 1.5 | 0.84 |
| Se | - | - | - | Na2SeO3·5H2O | 100 | 36.723 | 0.0377 | Na2SeO3·5H2O | 0.1 | 0.0367 |
| Ni | - | - | - | NiCl2·6H2O | 92 | 22.718 | 0.0233 | NiCl2·6H2O | 9 | 2.222 |
| Mn | MnCl2·4H2O | 13.19 | 3.662 | MnCl2·4H2O | 50 | 13.88 | 0.0143 | - | - | - |
| Cu | CuCl2·2H2O | 2.61 | 0.973 | CuCl2·2H2O | 38 | 14.164 | 0.0145 | - | - | - |
| Co | CoCl2·6H2O | 0.3 | 0.0801 | CoCl2·6H2O | 50 | 12.385 | 0.0127 | CoCl2·6H2O | 1.5 | 0.372 |
| K | KCl | 1 | 0.524 | K2HPO4·3H2O | 200,000 | 34,255.871 | 70.330 | K2HPO4 | load depending (60–200 mg/Lreactor·d) | |
| Fe | FeCl2·2H2O | 5.23 | 1.794 | FeCl2·4H2O | 0.686 | 0.000705 | FeCl2·4H2O | 750 | 210.678 | |
| EDTA | EDTA | 9.75 | 9.75 | EDTA | 500 | 500 | EDTA | 750 | 750 | |
| Na | NaCl | 200 | 78.678 | 10,000 | 3933.949 | 40.389 | Na2CO3 | 9000 | 1952.168 | |
| P | (NH4)2HPO4 | 200 | 46.909 | K2HPO4·3H2O | 200,000 | 27,140.702 | 55.722 | - | - | - |
| B | - | - | - | H3BO3 | 50 | 8.743 | 0.00898 | - | - | - |
| Al | - | - | - | AlCl3 | 50 | 10.118 | 0.0104 | - | - | - |
| S | - | - | - | Cystein hydrochloride C3H7NO2S | 500 | 98.494 | 1.001 | Na2S·9H2O | load depending (18–300 mg/Lreactor·d) | |
| N | (NH4Cl)2HPO4 | 200 | 21.213 | NH4Cl | 100,000 | 26,185.227 | 268.842 | NH4Cl | 7300 | 1911.522 |
| Element X | Compound | Element | |
|---|---|---|---|
| [Name] | [Formula] | [mg/L] | [mg X/L] |
| Stock Solution | Trickling Liquid | ||
| Mg | MgCl2·6H2O | 300 | 35.865 |
| Ca | CaCl2·2H2O | 51.308 | 13.994 |
| Zn | ZnCl2 | 0.0514 | 0.0246 |
| Mo | (NH4)6Mo7O24·2H2O | 1.5 | 0.84 |
| Ni | NiCl2·6H2O | 9 | 2.222 |
| Mn | MnCl2·4H2O | 0.0514 | 0.0143 |
| Cu | CuCl2·2H2O | 0.0390 | 0.0145 |
| Co | CoCl2·6H2O | 9 | 2.228 |
| K | K2HPO4·3H2O | 410.614 | 70.338 |
| Fe | FeCl2·4H2O | 750 | 211 |
| EDTA | EDTA | 750 | |
| B | H3BO3 | 0.0514 | 0.00898 |
| Al | AlCl3 | 0.0513 | 0.0104 |
| Na | NaCl | 102.679 | 40.393 |
| N | NH4Cl | 7300 | 1912 |
| P | contained in K2HPO4·3H2O | 27.955 | |
| S | supply via H2S in the biogas | 600–2000 mg/d | |
| Se | not contained | - | - |
| Parameter | Method | Comment |
|---|---|---|
| COD | DIN 38409 | DEV H 43-1 Short-term method |
| TKN, TN | DIN 19684 part 4 | DEV H11 Photometric variant |
| NH4-N | DIN 38406 | DEV D5-1 Photometric |
| PO4-P | DIN 38405 | DEV D11-3 Photometric |
| organic acids | DIN EN 17294 | Aminex HPX-87 H column, 5 mN H2SO4 mobile phase, UV-detector at 625 nm |
| DM, oDM | DIN 38409 part 1 | DEV H1 Annealing temperature 550 °C |
| SS | DIN 38409 part 2 | |
| pH-value | DIN 38404 part 15 | DEV C5 WTW SenTix20 combination electrode |
| CH4 | CO2 | O2 | H2S | H2 | H2O | Temperature | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | Mean, min/max | Mean ± SD | |||||||
| [%] | [%] | [%] | [ppm] | [%] | [%] | [°C] | |||||
| 65.4 | ±2.83 | 28.7 | ±4.55 | 0.4 | ±0.21 | 205 | 0–1958 | 0 | 6.52 | 37.9 | ±0.16 |
| CH4 | CO2 | O2 | H2S | H2 | pH-Value | Temperature |
|---|---|---|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
| [%] | [%] | [%] | [ppm] | [ppm] | [-] | [°C] |
| 96.6 ± 5.91 | 1.6 ± 2.42 | 0 ± 0.00 | 857 ± 861 | 19,169 ± 13,689 | 7.81 ± 0.45 | 36.9 ± 3.77 |
| Trace Element | Start Concentration | Demand | Range | Decay | Monthly Dosing |
|---|---|---|---|---|---|
| Name | [mg/L] | [mg/(L·d)] | [d] | [mg/(L·d)] | [mg/(L·mo)] |
| Fe | 28 | - | - | - | - |
| Ni | 8 | −0.13 | 61 | −0.13 | 4 |
| K | 230 | −6.95 | 33 | 0.20 | 210 |
| Na | 180 | −7.67 | 23 | 0.58 | 230 |
| Ca | 330 | −18.02 | 18 | −0.51 | 540 |
| Mg | 1230 | −49.15 | 25 | −1.18 | 1475 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tauber, J.; Möstl, D.; Vierheilig, J.; Saracevic, E.; Svardal, K.; Krampe, J. Biological Methanation in an Anaerobic Biofilm Reactor—Trace Element and Mineral Requirements for Stable Operation. Processes 2023, 11, 1013. https://doi.org/10.3390/pr11041013
Tauber J, Möstl D, Vierheilig J, Saracevic E, Svardal K, Krampe J. Biological Methanation in an Anaerobic Biofilm Reactor—Trace Element and Mineral Requirements for Stable Operation. Processes. 2023; 11(4):1013. https://doi.org/10.3390/pr11041013
Chicago/Turabian StyleTauber, Joseph, Daniel Möstl, Julia Vierheilig, Ernis Saracevic, Karl Svardal, and Jörg Krampe. 2023. "Biological Methanation in an Anaerobic Biofilm Reactor—Trace Element and Mineral Requirements for Stable Operation" Processes 11, no. 4: 1013. https://doi.org/10.3390/pr11041013