
Pachyrhizus erosus Inhibits Adipogenesis via the Leptin-PPARγ-FAS Pathway in a High-Fat Diet-Induced Mouse Model

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Pachyrhizus erosus Preparation
2.2. Animal Experiments and Bodyweight Measurement
2.3. Ethics Statement
2.4. Blood Chemistry Analysis and Organ Weight Measurement
2.5. Histopathological Analysis
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Real-Time Polymerase Chain Reaction (RT-PCR) Analysis
2.8. Immunofluorescent Analysis
2.9. Inulin Analysis
2.10. Statistical Analysis
3. Results
3.1. P. erosus Effectively Controlled HFD-Induced Body Weight Gaining
3.2. P. erosus Decreased Obesity-Related Biomarkers in the Serum
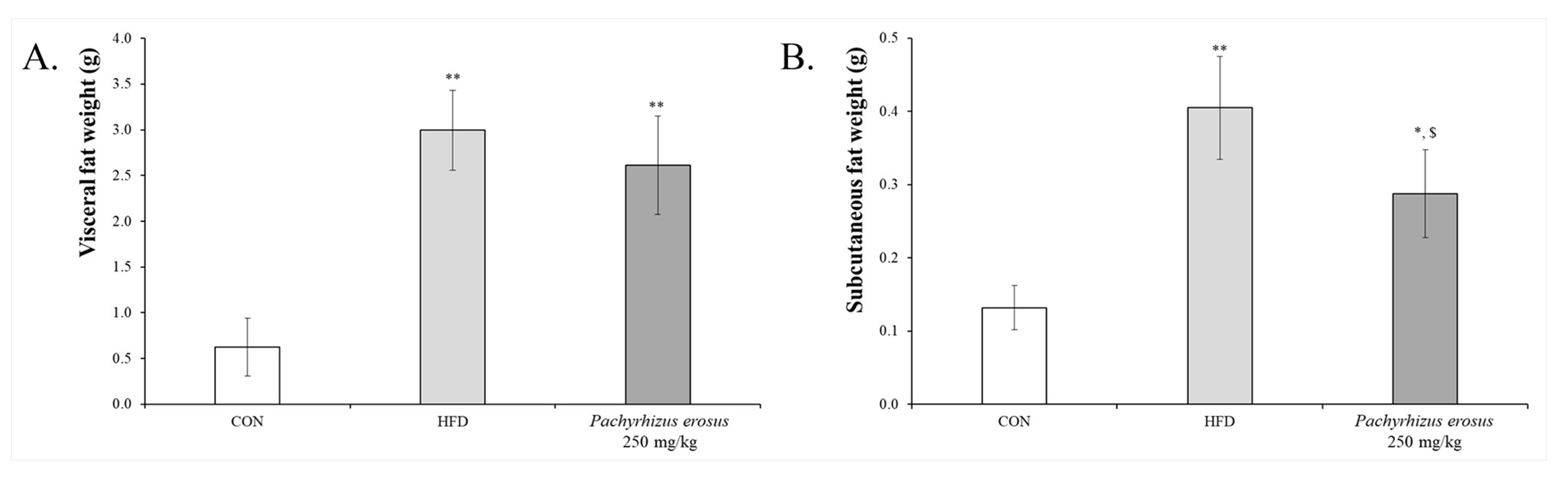
3.3. P. erosus Suppressed Fat Tissues Weight Gain
3.4. P. erosus Significantly Controlled the Fat Accumulation in the Liver
3.5. P. erosus Inhibited Adipogenesis via Leptin-PPARγ/C/EBPα-FAS
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. Obesity. 2 May 2022. Available online: https://www.who.int/health-topics/obesity#tab=tab_1 (accessed on 1 August 2022).
- Wright, S.M.; Aronne, L.J. Causes of obesity. Abdom. Imaging 2012, 37, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Aronne, L.J.; Nelinson, D.S.; Lillo, J.L. Obesity as a disease state: A new paradigm for diagnosis and treatment. Clin. Cornerstone 2009, 9, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Kushner, R.F.; Roth, J.L. Assessment of the obese patient. Endocrinol. Metab. Clin. N. Am. 2003, 32, 915–933. [Google Scholar] [CrossRef] [PubMed]
- World Obesity Federation. World Obesity Atlas 2022. Available online: https://s3-eu-west-1.amazonaws.com/wof-files/World_Obesity_Atlas_2022.pdf (accessed on 1 August 2022).
- Pillon, N.J.; Loos, R.J.F.; Marshall, S.M.; Zierath, J.R. Metabolic consequences of obesity and type 2 diabetes: Balancing genes and environment for personalized care. Cell 2021, 184, 1530–1544. [Google Scholar] [CrossRef]
- Powell-Willey, T.M.; Poirier, C.P.; Burke, L.E.; Despres, J.P.; Gordon-Larsen, P.; Lavie, C.J.; Lear, S.A.; Ndumele, C.E.; Neeland, I.J.; Sanders, P.; et al. American Heart Association Council on Lifestyle and Cardiometabolic Health; Council on Cardiovascular and Stroke Nursing; Council on Clinical Cardiology; Council on Epidemiology and Prevention; and Stroke Council. Obesity and cardiovascular disease: A scientific statement from the American Heart Association. Circulation 2021, 143, e984–e1010. [Google Scholar] [CrossRef]
- Wang, X.; Rao, H.; Liu, F.; Wei, L.; Li, H.; Wu, C. Recent advances in adiopose tissue dysfunction and its role in the pathogenesis of non-alcoholic fatty liver disease. Cells 2021, 10, 3300. [Google Scholar] [CrossRef]
- Kushner, R.F.; Weinsier, R.L. Evaluation of the obese patient. Med. Clin. N. Am. 2000, 84, 387–399. [Google Scholar] [CrossRef]
- Taylor, E.B. The complex role of adipokines in obesity, inflammation, and autoimmunity. Clin. Sci. 2021, 135, 731–752. [Google Scholar] [CrossRef]
- Nawaz, A.; Aminuddin, A.; Kado, T.; Takikawa, A.; Yamamoto, S.; Tsuneyama, K.; Igarashi, Y.; Ikutani, M.; Nishida, Y.; Nagai, Y.; et al. CD206+ M2-like macrophages regulate systemic glucose metabolism by inhibiting proliferation of adipocyte progenitors. Nat. Commun. 2017, 8, 286. [Google Scholar] [CrossRef] [Green Version]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Kratz, M.; Coasts, B.R.; Hisert, K.B.; Hagman, D.; Mutskov, V.; Peris, E.; Schoenfelt, K.Q.; Kuzma, J.N.; Larson, I.; Billing, P.S.; et al. Metabolic dysfunction drives a mechanistically distinct proinflammatory phenotype in adipose tissue macrophage. Cell Metab. 2014, 20, 614–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Department of Nutritional Function Research. Possible to help body fat decrease. In Efficacy Evaluation Guide for Functional Food; Korea National Institute of Food and Drug Safety Evaluation: Osong, Republic of Korea, 2019; pp. 8–10. [Google Scholar]
- Gwon, S.Y.; Ahn, J.Y.; Kim, T.W.; Ha, T.Y. Zanthoxylum peperitum DC ethanol extract suppresses fat accumulation in adipocytes and high fat diet-induced obese mice by regulating adipogenesis. J. Nutr. Sci. Vitaminol. 2012, 58, 393–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palhinha, L.; Liechocki, S.; Hottz, E.D.; Pereira, J.A.d.S.; de Almeida, C.J.; Moraes-Vieira, P.M.M.; Bozza, P.T.; Maya-Monteiro, C.M. Leptin induces proadipogenic and proinflammatory signaling in adipocytes. Front. Endocrinol. 2019, 10, 841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPα induces adipogenesis through PPARγ: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Jensen-Urstad, A.P.L.; Semenkovich, C.F. Fatty acid synthase and liver triglyceride metabolism: Housekeeper or messenger? Biochim. Biophys. Acta 2012, 1821, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Muller, T.D.; Bluher, M.; Tschop, M.H.; DiMarchi, R.D. Anti-obesity drug discovery: Advances and challenges. Nat. Rev. Drug Discov. 2022, 21, 201–223. [Google Scholar] [CrossRef]
- Sanderson, H.; Prance, G.; Nesbitt, M. (Eds.) The Culture History of Plants; Routledge: New York, NY, USA, 2005; ISBN 0-415-92746-3. [Google Scholar]
- Pertiwi, R.; Samudra, A.G.; Wati, N.K.; Hanuun, A.; Yudha, S.S.; Notriawan, D.; Wibowo, R.H. Gastroprotective activities of Peperomia pellucida L. and Pachyrhizus erosus L. extracts combination on ethanol-induced rats. Inflammopharmacolgy 2022, 30, 2139–2144. [Google Scholar] [CrossRef]
- Santoso, P.; Amelia, A.; Rahayu, R. Jicama (Pachyrhizus erosus) fiber prevents excessive blood glucose and body weight increase without affective food intake in mice fed with high-sugar diet. J. Adv. Vet. Anim. Res. 2019, 6, 222–230. [Google Scholar] [CrossRef]
- Park, C.J.; Lee, H.A.; Han, J.S. Jicama (Pachyrhizus erosus) extract increases insulin sensitivity and regulates hepatic glucose in C57BL/Ksj-db/db mice. J. Clin. Biochem. Nutr. 2016, 58, 56–63. [Google Scholar] [CrossRef] [Green Version]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugarts and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicyclic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Almari, M.; Mohammad, A.; Abubaker, J.; Ziyab, A.H. Obesity and prediabetes are jointly associated with lipid abnormalities among adolescents: A cross-sectional study. Diabetes Metab. Syndr. Obes. 2021, 14, 345–353. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, K.J.; Kim, Y.H.; Kim, D.B.; Shin, G.H.; Cho, J.H.; Kim, B.K.; Lee, B.Y.; Lee, O.H. Codonopsis lanceolate extract prevents diet-induced obesity in C57BL/6 mice. Nutrients 2014, 6, 4663–4677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Nara, N.; Nakayama, Y.; Okamoto, S.; Tamura, H.; Kiyono, M.; Muraoka, M.; Tanaka, K.; Taya, C.; Shitara, H.; Ishii, R.; et al. Disruption of CXC motif chemokine ligand-14 in mice ameliorates obesity-induced insulin resistance. J. Biol. Chem. 2007, 282, 30794–30803. [Google Scholar] [CrossRef] [Green Version]
- Duffaut, C.; Zakaroff-Girard, A.; Bourlier, V.; Decaunes, P.; Maumus, M.; Chiotasso, P.; Sengenes, C.; Lafontan, M.; Galitzky, J.; Bouloumie, A. Interplay between human adipocytes and T lymphocytes in obesity. CCL20 as an adipochemokine and T lymphocytes as lipogenic modulators. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1608–1614. [Google Scholar] [CrossRef] [Green Version]
- Beisner, J.; Rosa, L.F.; Kaden-Volynets, V.; Stolzer, I.; Gunther, C.; Bischoff, S.C. Prebiotic inulin and sodium butyrate attenuate obesity-induced intestinal barrier dysfunction by induction of antimicrobial peptides. Front. Immunol. 2021, 12, 678360. [Google Scholar] [CrossRef]
- Van der Beek, C.M.; Canfora, E.; Kip, A.M.; Gorissen, S.H.M.; Damink, S.W.M.O.; Van Eijk, H.M.; Holst, J.J.; Blaak, E.E.; Dejong, C.H.C.; Lenaerts, K. The prebiotic inulin improves substrate metabolism and promotes short-chain fatty acid production in overweight to obese man. Metabolism 2018, 87, 25–35. [Google Scholar] [CrossRef]
- Antel, J.; Hebenbrand, J. Weight-reducing side effects of the antiepileptic agents topiramate and zonisamide. Handb. Exp. Pharmacol. 2012, 209, 433–466. [Google Scholar] [CrossRef]
- Rufino, A.T.; Costa, V.M.; Carvalho, F.; Fernandes, E. Flavonoids as antiobesity agents: A review. Med. Res. Rev. 2021, 41, 556–585. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 mg/mL of Pachyrhizus erosus Extract | ||
|---|---|---|
| Total Sugar (mg/mL) | Reducing Sugar (mg/mL) | Inulin (mg/mL) |
| 0.78 ± 0.029 | 0.39 ± 0.008 | 0.38 ± 0.029 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-M.; Bok, S.-H.; Kim, M.-H.; Lee, K.S.; Park, D.-H. Pachyrhizus erosus Inhibits Adipogenesis via the Leptin-PPARγ-FAS Pathway in a High-Fat Diet-Induced Mouse Model. Processes 2023, 11, 735. https://doi.org/10.3390/pr11030735
Lee S-M, Bok S-H, Kim M-H, Lee KS, Park D-H. Pachyrhizus erosus Inhibits Adipogenesis via the Leptin-PPARγ-FAS Pathway in a High-Fat Diet-Induced Mouse Model. Processes. 2023; 11(3):735. https://doi.org/10.3390/pr11030735
Chicago/Turabian StyleLee, Seung-Min, So-Hyeon Bok, Min-Hee Kim, Ki Sun Lee, and Dae-Hun Park. 2023. "Pachyrhizus erosus Inhibits Adipogenesis via the Leptin-PPARγ-FAS Pathway in a High-Fat Diet-Induced Mouse Model" Processes 11, no. 3: 735. https://doi.org/10.3390/pr11030735