
Anti-Obesity Activity in 3T3-L1 Cells of Cornus officinalis Fruits Harvested at Different Times
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Cell Culture of 3T3-L1 Pre-Adipocytes
2.3. Cell Viability of Pre-Adipocytes
2.4. Differentiation of Pre-Adipocytes
2.5. Oil Red O Staining of Adipocytes
2.6. Western Blot Analysis of Adipocytes
2.7. Antioxidant Activity Assays
2.7.1. DPPH Free Radical Scavenging Assay
2.7.2. ABTS Radical Scavenging Assay
2.7.3. Determination of TP Content
2.7.4. FRAP Assay
2.8. HPLC Analysis of Major Active Components
2.9. Statistical Analysis
3. Results
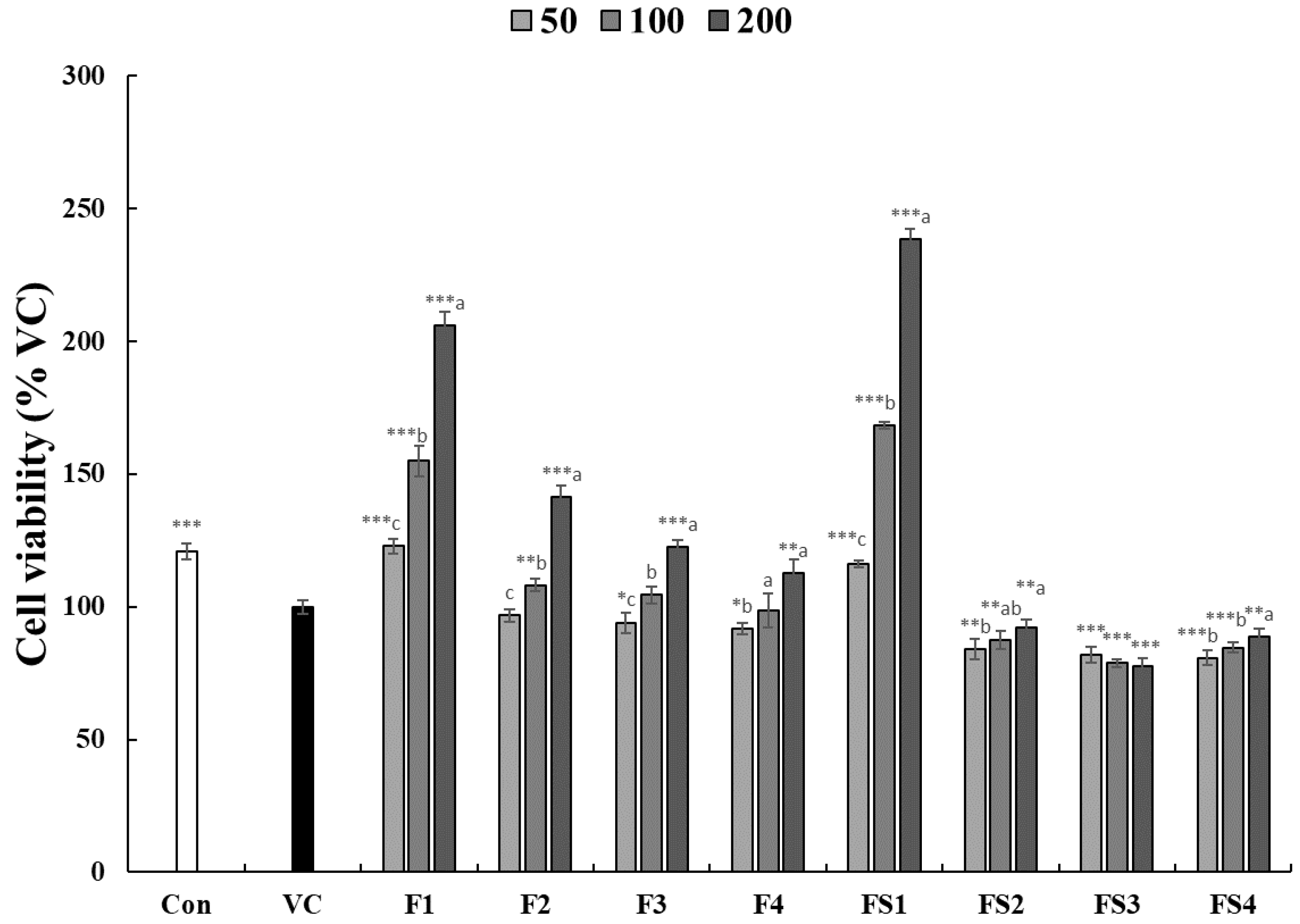
3.1. Effect of COF on Cell Viability
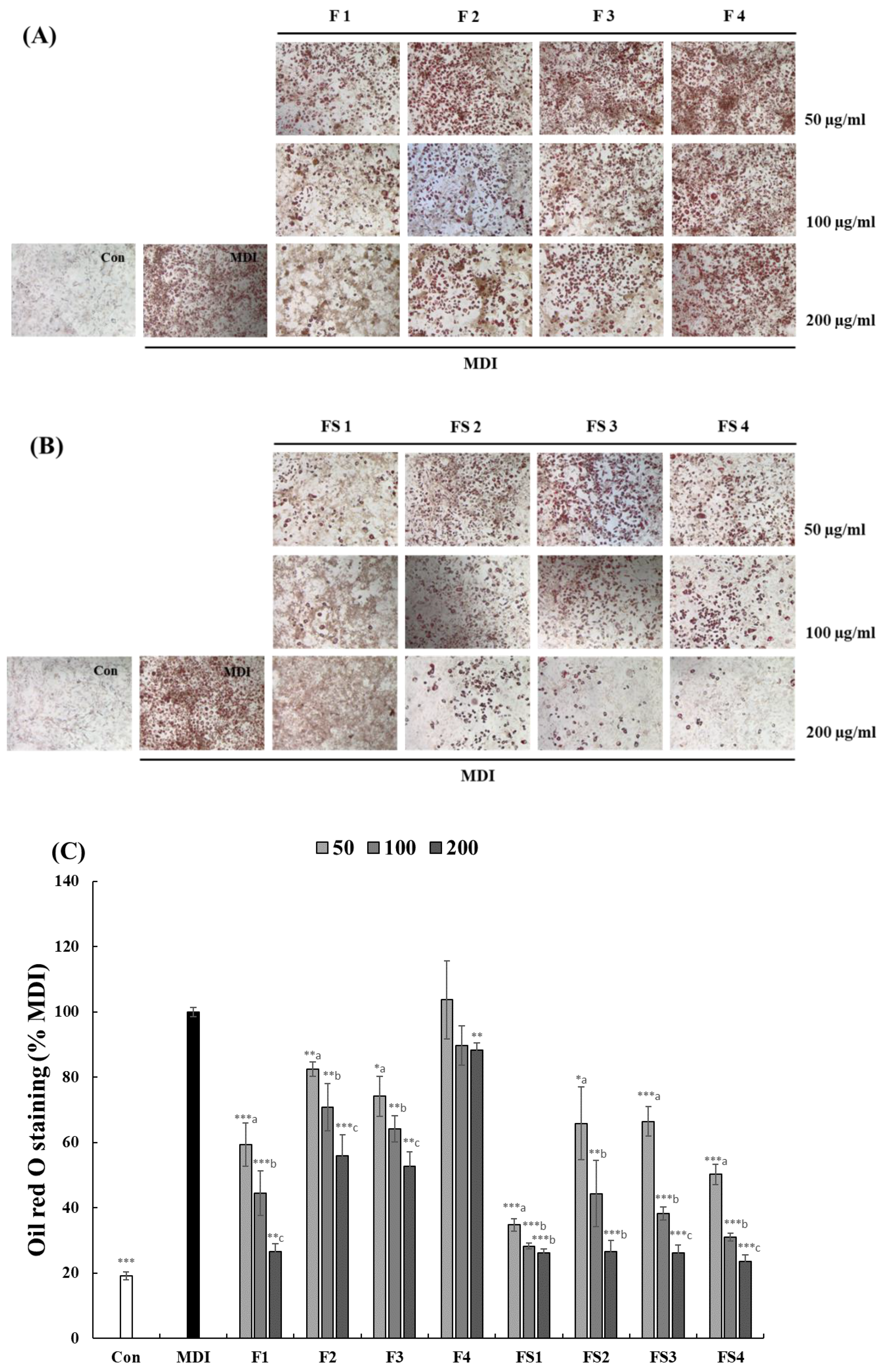
3.2. Effect of COF on 3T3-L1 Cell Differentiation
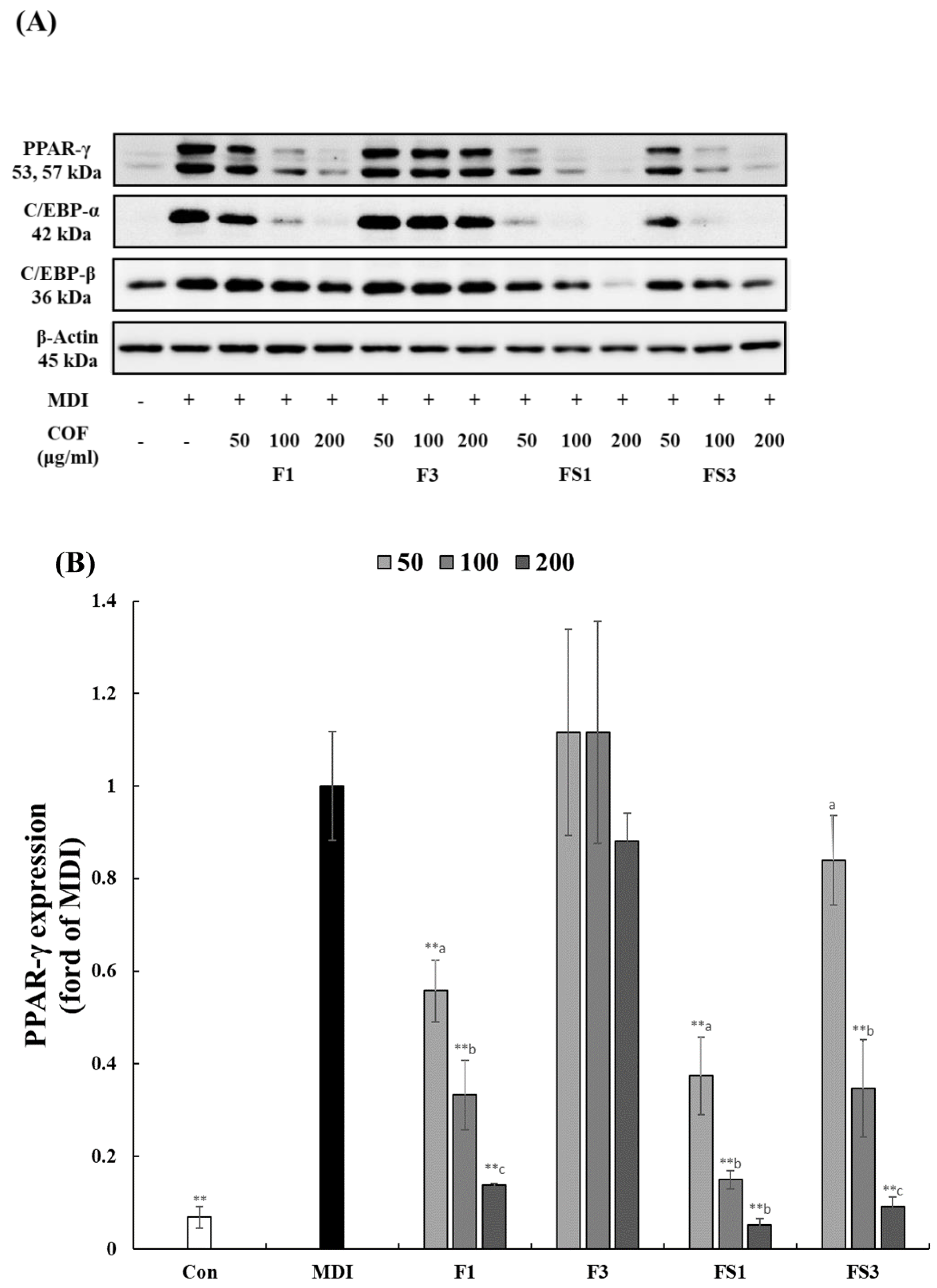
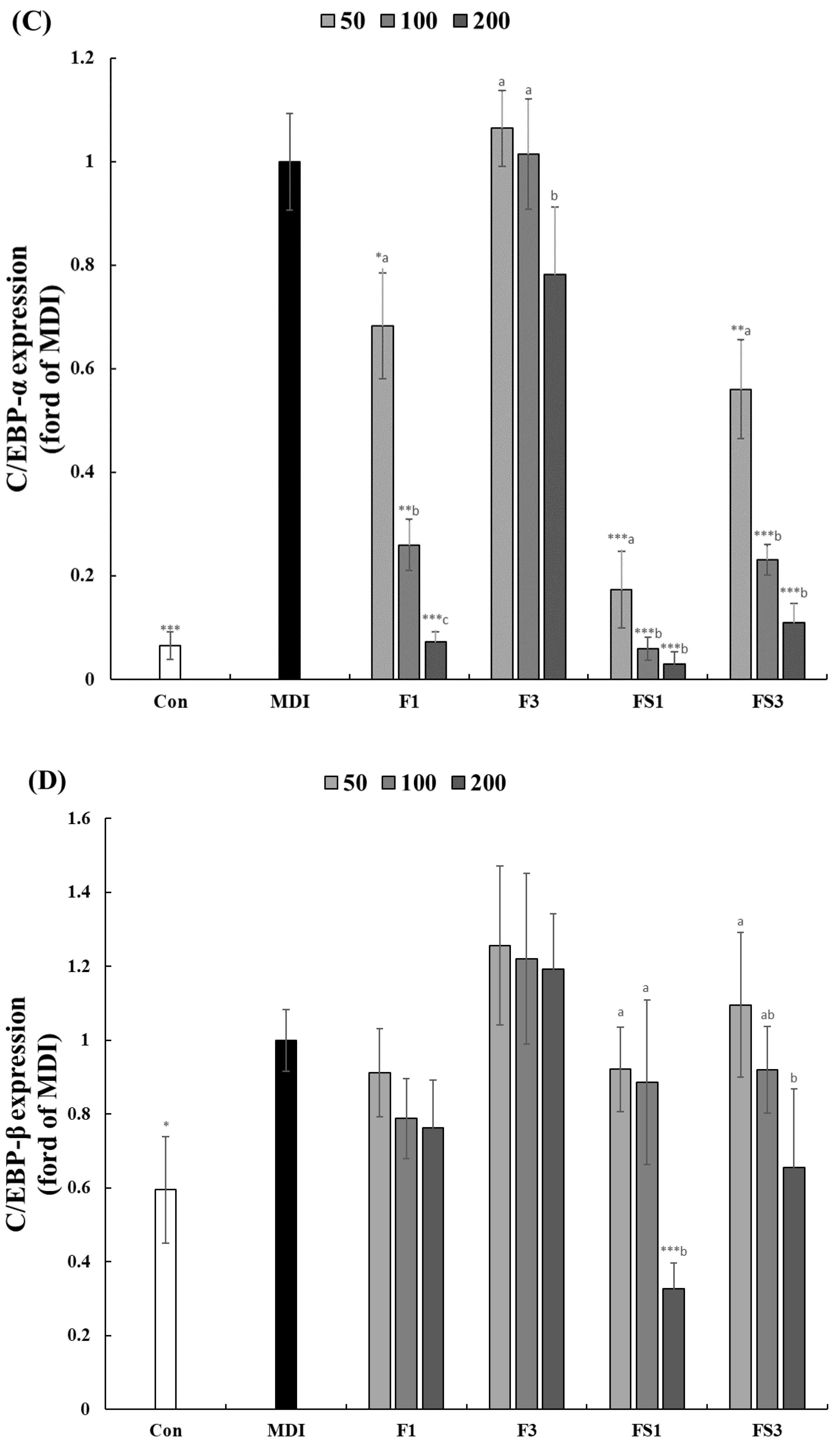
3.3. COF Inhibits the Expression of C/EBPα, β, and PPAR-γ
3.4. Antioxidant Activity of COF
3.5. Active Components
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Park, E.K.; Lee, C.G.; Kim, J.; Lim, E.; Hwang, S.; Yun, S.H.; Jeong, S.Y. Anti-osteoporotic effects of the herbal mixture of Cornus officinalis and Achyranthes japonica in vitro and in vivo. Plants 2020, 9, 1114. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.R.; Jeune, K.H.; Park, S.Y.; Jang, S.J. Toxicity and lectins constituents from the seed of Cornus officinalis. Kor. J. Pharmacogn. 1993, 24, 177–182. [Google Scholar]
- Hwang, K.A.; Hwang, Y.J.; Song, J. Antioxidant activities and oxidative stress inhibitory effects of ethanol extracts from Cornus officinalis on raw 264.7 cells. BMC Complement Altern. Med. 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Zhu, Z.; Zhang, H.; Tan, G.; Chen, X.; Chai, Y. Qualitative and quantitative analysis of Fructus corni using ultrasound-assisted microwave extraction and high-performance liquid chromatography coupled with diode array UV detection and time-of-flight mass spectrometry. J. Pharm. Biomed. Anal. 2011, 55, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, Y.; Dong, L.; Gao, Q.; Yin, L.; Quan, H.; Chen, R.; Fu, X.; Lin, D. Ethnopharmacology, phytochemistry, and pharmacology of Cornus officinalis Sieb. et Zucc. J. Ethnopharmacol. 2018, 213, 280–301. [Google Scholar] [CrossRef]
- Hwang, J.H.; Lee, S.W.; Han, K.H.; Seo, H.J.; Kim, J.D. Anti-angiogenesis and anti-adipogenesis effects of Anthrisci radix extract. Biotechnol. Bioproc. Eng. 2013, 18, 164–172. [Google Scholar] [CrossRef]
- Lee, W.J.; Koh, E.H.; Won, J.C.; Kim, M.S.; Park, J.Y.; Lee, K.U. Obesity: The role of hypothalamic AMP-activated protein kinase in body weight regulation. Int. J. Biochem. Cell Biol. 2005, 37, 2254–2259. [Google Scholar] [CrossRef]
- Park, C.H.; Rhyu, D.Y.; Sharma, B.R.; Yokozawa, T. Inhibition of preadipocyte differentiation and lipid accumulation by 7-O-galloyl-d-sedoheptulose treatment in 3T3-L1 adipocytes. Biomed. Prev. Nutr. 2013, 3, 319–324. [Google Scholar] [CrossRef]
- Farmer, S.R. Regulation of PPARc activity during adipogenesis. Int. J. Obes. 2005, 29, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Guan, H.P.; Ishizuka, T.; Chui, P.C.; Lehrke, M.; Lazar, M.A. Corepressors selectively control the transcriptional activity of PPARc in adipocytes. Genes. Dev. 2005, 19, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Christy, R.J.; Kaestner, K.H.; Geiman, D.E.; Lane, M.D. CCAAT/enhancer binding protein gene promoter: Binding of nuclear factors during differentiation of 3T3-L1 preadipocytes. Proc. Natl. Acad. Sci. USA 1991, 88, 2593–2597. [Google Scholar] [CrossRef] [PubMed]
- Hamm, J.K.; Park, B.H.; Farmer, S.R. A role for C/EBPβ in regulating peroxisome proliferator-activated receptor γ activity during adipogenesis in 3T3-L1 preadipocytes. J. Biol. Chem. 2001, 276, 18464–18471. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.H.; Kwon, Y.J.; So, J.S. Anti-adipogenic effects of Corni fructus in 3T3-L1 preadipocytes. Biotechnol. Bioprocess Eng. 2014, 19, 52–57. [Google Scholar] [CrossRef]
- Tepe, B.; Sokmen, M.; Akpulat, H.A.; Sokmen, A. Screening of the antioxidant potentials of six Salvia species from Turkey. Food Chem. 2006, 95, 200–204. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice Evans, C. 1999 Antioxidant activity applying an im-proved ABTS radical cation decolorization assay. Free Radicals Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Mau, J.L.; Lin, H.C.; Song, S.F. Antioxidant properties of several specialty mushrooms. Food Res Int. 2002, 35, 519–526. [Google Scholar] [CrossRef]
- Jang, Y.; Kang, H.; Kim, J.; Lee, S.H. Anti-obesity effects of an enzymatic extract of mandarin (Citrus unshiu) peel in 3T3-L1 adipocytes. Korean J. Food Sci. Technol. 2021, 53, 149–153. [Google Scholar]
- Park, M.W.; Park, Y.K.; Kim, E.S. Properties of phenolic compounds in unripe apples. J. East Asian Soc. Dietary Life 2004, 14, 343–347. [Google Scholar]
- Danielewski, M.; Matuszewska, A.; Szeląg, A.; Sozański, T. The impact of anthocyanins and iridoids on transcription factors crucial for lipid and cholesterol homeostasis. Int. J. Mol. Sci. 2021, 22, 6074. [Google Scholar] [CrossRef]
- Boccellino, M.; D’ Angelo, S. Anti-obesity effects of polyphenol intake: Current status and future possibilities. Int. J. Mol. Sci. 2020, 21, 5642. [Google Scholar] [CrossRef] [PubMed]
- Pablo, C.; Daiana, S.; Soledad, C.; Virginia, D.; Lelia, D.; Juan, C.C.; Liliana, N.G. N-Acetylcysteine reduces markers of differentiation in 3T3-L1 adipocytes. Int. J. Mol. Sci. 2011, 12, 6936–6951. [Google Scholar]
- Lee, Y.J.; Yoon, B.R.; Choi, H.S.; Lee, B.Y.; Lee, O.H. Effect of Sargassum micracanthum extract on lipid accumulation and reactive oxygen species (ROS) production during differentiation of 3T3-L1 preadipocytes. Korean J. Food Preserv. 2012, 19, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Ye, Z.; Tian, T.; Zhu, R.; Liu, C.; Fang, X.; Zhang, D.; Fu, M.; Gao, S.; Zhao, D. Loganin regulates glycolipid metabolism by influencing intestinal microbiota and AMPK signaling in obese mice. J. Tradit. Chin. Med. Sci. 2022, 9, 321–329. [Google Scholar] [CrossRef]
- Yang, Q.; Shu, F.; Gong, J.; Ding, P.; Cheng, R.; Li, J.; Tong, R.; Ding, L.; Sun, H.; Huang, W.; et al. Sweroside ameliorates NAFLD in high-fat diet induced obese mice through the regulation of lipid metabolism and inflammatory response. J. Ethnopharmacol. 2020, 255, 112556. [Google Scholar] [CrossRef]
- Kim, N.H.; Jegal, J.; Kim, Y.N.; Heo, J.D.; Rho, J.R.; Yang, M.H.; Jeong, E.J. Chokeberry extract and its active polyphenols suppress adipogenesis in 3T3-L1 adipocytes and modulates fat accumulation and insulin resistance in diet-induced obese mice. Nutrients 2018, 10, 1734. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPAR is required for differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Gustafson, B.; Smith, U. Cytokines promote Wnt signaling and inflammation and impair the normal differentiation and lipid accumulation in 3T3-L1 preadipocytes. J. Biol. Chem. 2006, 281, 9507–9516. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Harvest Time | |
|---|---|---|
| F1 | Pericarp | 15 September |
| F2 | 15 October | |
| F3 | 15 November | |
| F4 | 15 December | |
| FS1 | Pericarp + seeds | 15 September |
| FS2 | 15 October | |
| FS3 | 15 November | |
| FS4 | 15 December |
| Compound | Linear Range (μg/mL) | Calibration Equation | Correlation Coefficient (r2) |
|---|---|---|---|
| Gallic acid | 2.5–100 | y = 10.5884x + 7.5749 | R2 = 0.9998 |
| Morroniside | 40–200 | y = 4.1525x + 11.5427 | R2 = 0.9998 |
| Loganin | 2.5–100 | y = 4.4595x − 0.3537 | R2 = 1.0000 |
| Verbenalin | 10–400 | y = 4.6197x − 18.8144 | R2 = 0.9996 |
| Sweroside | 2.5–200 | y = 7.8192x − 6.9401 | R2 = 0.9997 |
| Cornuside | 2.5–100 | y = 5.7834x − 0.4795 | R2 = 1.0000 |
| DPPH (IC50, μg/mL) | ABTS (IC50, μg/mL) | FRAP (mg TE/g) | TPC (mg GAE/g) | |
|---|---|---|---|---|
| F1 | 55.93 ± 0.36 d | 41.37 ± 0.27 d | 539.95 ± 1.57 c | 135.84 ± 3.08 d |
| F2 | 106.56 ± 0.29 c | 81.72 ± 0.57 c | 193.92 ± 1.57 f | 68.51 ± 1.68 e |
| F3 | 130.51 ± 0.81 b | 93.48 ± 0.64 b | 140.41 ± 0.00 g | 69.40 ± 1.33 e |
| F4 | 219.60 ± 1.54 a | 151.44 ± 2.52 a | 71.67 ± 0.20 h | 39.37 ± 1.67 f |
| FS1 | 15.89 ± 0.03 g | 13.28 ± 0.10 g | 871.16 ± 2.36 a | 288.96 ± 4.23 a |
| FS2 | 35.83 ± 0.05 e | 31.58 ± 0.09 e | 413.42 ± 0.79 d | 142.07 ± 3.33 c |
| FS3 | 35.24 ± 0.16 e | 30.77 ± 0.35 e | 401.63 ± 1.36 e | 139.18 ± 3.67 cd |
| FS4 | 25.54 ± 0.08 f | 21.97 ± 0.16 f | 549.02 ± 1.57 b | 187.84 ± 3.42 b |
| Trolox | 9.35 ± 0.13 | |||
| Ascorbic acid | 7.89 ± 0.08 |
| Gallic Acid (mg/g) | Morroniside (mg/g) | Loganin (mg/g) | Verbenalin (mg/g) | Sweroside (mg/g) | Cornuside (mg/g) | |
|---|---|---|---|---|---|---|
| F1 | 1.49 ± 0.08 a | 63.95 ± 0.49 a | 25.85 ± 0.14 c | 3.19 ± 0.51 d | 2.71 ± 0.14 d | 4.07 ± 0.07 b |
| F2 | 0.28 ± 0.03 e | 25.38 ± 0.62 d | 10.09 ± 0.22 f | 0.66 ± 0.04 e | 0.94 ± 0.10 e | 1.62 ± 0.02 d |
| F3 | 0.14 ± 0.01 f | 23.56 ± 0.76 e | 11.79 ± 0.36 f | 0.75 ± 0.06 e | 0.98 ± 0.06 e | 1.61 ± 0.07 d |
| F4 | 1.02 ± 0.01 b | 31.15 ± 0.95 c | 15.42 ± 0.38 e | 0.57 ± 0.06 e | 1.02 ± 0.10 e | 1.85 ± 0.03 d |
| FS1 | 1.50 ± 0.07 a | 48.28 ± 0.32 b | 38.95 ± 0.39 a | 37.10 ± 0.80 a | 12.52 ± 0.47 a | 5.68 ± 0.10 a |
| FS2 | 0.82 ± 0.11 c | 20.11 ± 1.50 f | 20.53 ± 2.53 d | 17.55 ± 2.77 c | 6.95 ± 1.05 c | 2.20 ± 0.28 c |
| FS3 | 0.49 ± 0.11 d | 20.00 ± 1.51 f | 20.44 ± 1.25 d | 16.90 ± 0.46 c | 6.36 ± 0.38 c | 1.82 ± 0.22 d |
| FS4 | 1.39 ± 0.06 a | 26.76 ± 1.38 d | 28.49 ± 1.08 b | 27.05 ± 1.16 b | 8.82 ± 0.53 b | 2.33 ± 0.10 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.S.; Choi, S.J.; Kim, H.D.; Kang, M.H.; Ji, Y.-J.; Kim, G.-S.; Jang, G.Y. Anti-Obesity Activity in 3T3-L1 Cells of Cornus officinalis Fruits Harvested at Different Times. Processes 2022, 10, 2008. https://doi.org/10.3390/pr10102008
Lee ES, Choi SJ, Kim HD, Kang MH, Ji Y-J, Kim G-S, Jang GY. Anti-Obesity Activity in 3T3-L1 Cells of Cornus officinalis Fruits Harvested at Different Times. Processes. 2022; 10(10):2008. https://doi.org/10.3390/pr10102008
Chicago/Turabian StyleLee, Eun Suk, Su Ji Choi, Hyung Don Kim, Min Hye Kang, Yun-Jeong Ji, Geum-Soog Kim, and Gwi Yeong Jang. 2022. "Anti-Obesity Activity in 3T3-L1 Cells of Cornus officinalis Fruits Harvested at Different Times" Processes 10, no. 10: 2008. https://doi.org/10.3390/pr10102008