
Effects of Sepsis on Immune Response, Microbiome and Oxidative Metabolism in Preterm Infants
 , , , , , , , ,
, , , , , , , ,  and
and 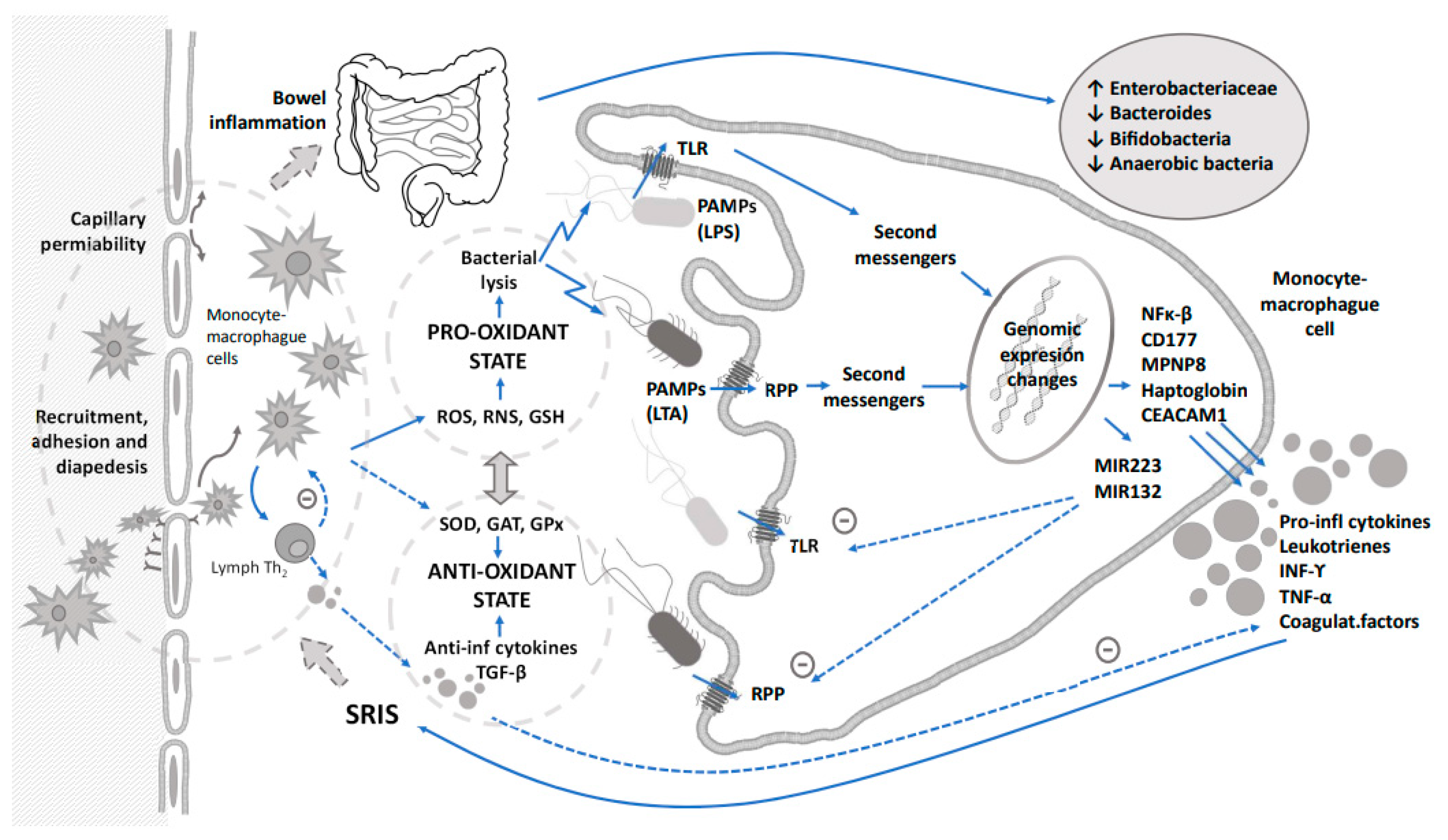{kind=link}
Abstract
:1. Introduction
2. Immune Response
3. Oxidative Stress and Metabolic Processes
3.1. Principles of Oxidative Stress
3.2. Pathophysiology of Newborn Sepsis
3.3. Oxidative Stress in Preterm Infants with Sepsis
3.4. Biomarkers of Oxidative Stress in Neonatal Sepsis
3.5. Antioxidant Components of Human Milk
4. Microbiome
4.1. Microbiome and Sepsis
4.2. Bacterial Metabolism
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fleischmann, C.; Reichert, F.; Cassini, A.; Horner, R.; Harder, T.; Markwart, R.; Tröndle, M.; Savova, Y.; Kissoon, N.; Schlattmann, P.; et al. Global incidence and mortality of neonatal sepsis: A systematic review and meta-analysis. Arch. Dis. Child. 2021, 106, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Puopolo, K.M.; Benitz, W.E.; Zaoutis, T.E.; Committee on Fetus and Newborn; Committee on Infectious Diseases. Management of Neonates Born at ≤ 34 6/7 Weeks’ Gestation With Suspected or Proven Early-Onset Bacterial Sepsis. Pediatrics 2018, 142, e20182896. [Google Scholar] [CrossRef] [Green Version]
- Flannery, D.D.; Puopolo, K.M. Neonatal Early-Onset Sepsis. Neoreviews 2022, 23, 756–770. [Google Scholar] [CrossRef]
- Brumbaugh, J.E.; Bell, E.F.; Do, B.T.; Greenberg, R.G.; Stoll, B.J.; DeMauro, S.B.; Harmon, H.M.; Hintz, S.R.; Das, A.; Puopolo, K.M.; et al. Incidence of and Neurodevelopmental Outcomes After Late-Onset Meningitis Among Children Born Extremely Preterm. JAMA Netw. Open. 2022, 5, e2245826. [Google Scholar] [CrossRef]
- Coggins, S.A.; Glaser, K. Updates in Late-Onset Sepsis: Risk Assessment, Therapy, and Outcomes. Neoreviews 2022, 23, 738–755. [Google Scholar] [CrossRef] [PubMed]
- Wynn, J.L.; Wong, H.R.; Shanley, T.P.; Bizzarro, M.J.; Saiman, L.; Polin, R.A. Time for a neonatal-specific consensus definition for sepsis. Pediatr. Crit. Care Med. 2014, 15, 523–528. [Google Scholar] [CrossRef] [Green Version]
- Shane, A.L.; Sánchez, P.J.; Stoll, B.J. Neonatal sepsis. Lancet 2017, 390, 1770–1780. [Google Scholar] [CrossRef] [PubMed]
- Biondi, E.A.; Mischler, M.; Jerardi, K.E.; Statile, A.M.; French, J.; Evans, R.; Lee, V.; Chen, C.; Asche, C.; Ren, J.; et al. Pediatric Research in Inpatient Settings (PRIS) Network. Blood culture time to positivity in febrile infants with bacteremia. JAMA Pediatr. 2014, 168, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Straub, J.; Paula, H.; Mayr, M.; Kasper, D.; Assadian, O.; Berger, A.; Rittenschober-Böhm, J. Diagnostic accuracy of the ROCHE Septifast PCR system for the rapid detection of blood pathogens in neonatal sepsis-A prospective clinical trial. PLoS ONE 2017, 12, e0187688. [Google Scholar] [CrossRef] [Green Version]
- van den Brand, M.; van den Dungen, F.A.M.; Bos, M.P.; van Weissenbruch, M.M.; van Furth, A.M.; de Lange, A.; Rubenjan, A.; Peters, R.P.H.; Savelkoul, P.H.M. Evaluation of a real-time PCR assay for detection and quantification of bacterial DNA directly in blood of preterm neonates with suspected late-onset sepsis. Crit. Care 2018, 22, 105. [Google Scholar] [CrossRef]
- Pammi, M.; Flores, A.; Leeflang, M.; Versalovic, J. Molecular assays in the diagnosis of neonatal sepsis: A systematic review and meta-analysis. Pediatrics 2011, 128, e973–e985. [Google Scholar] [CrossRef] [PubMed]
- Donà, D.; Mozzo, E.; Mardegan, V.; Trafojer, U.; Lago, P.; Salvadori, S.; Baraldi, E.; Giaquinto, C. Antibiotics Prescriptions in the Neonatal Intensive Care Unit: How to Overcome Everyday Challenges. Am. J. Perinatol. 2017, 34, 1169–1177. [Google Scholar] [PubMed]
- Collado, M.C.; Cernada, M.; Neu, J.; Pérez-Martínez, G.; Gormaz, M.; Vento, M. Factors influencing gastrointestinal tract and microbiota immune interaction in preterm infants. Pediatr. Res. 2015, 77, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Ting, J.Y.; Synnes, A.; Roberts, A.; Deshpandey, A.; Dow, K.; Yoon, E.W.; Lee, K.S.; Dobson, S.; Lee, S.K.; Shah, P.S. Association Between Antibiotic Use and Neonatal Mortality and Morbidities in Very Low-Birth-Weight Infants Without Culture-Proven Sepsis or Necrotizing Enterocolitis. JAMA Pediatr. 2016, 170, 1181–1187. [Google Scholar] [CrossRef]
- Cernada, M.; Badía, N.; Modesto, V.; Alonso, R.; Mejías, A.; Golombek, S.; Vento, M. Cord blood interleukin-6 as a predictor of early-onset neonatal sepsis. Acta Paediatr. 2012, 101, e203–e207. [Google Scholar] [CrossRef] [PubMed]
- Auriti, C.; Fiscarelli, E.; Ronchetti, M.P.; Argentieri, M.; Marrocco, G.; Quondamcarlo, A.; Seganti, G.; Bagnoli, F.; Buonocore, G.; Serra, G.; et al. Procalcitonin in detecting neonatal nosocomial sepsis. Arch. Dis. Child Fetal Neonatal. 2012, 97, F368–F370. [Google Scholar] [CrossRef]
- Stocker, M.; Fontana, M.; El Helou, S.; Wegscheider, K.; Berger, T.M. Use of procalcitonin-guided decision-making to shorten antibiotic therapy in suspected neonatal early-onset sepsis: Prospective randomized intervention trial. Neonatology 2010, 97, 165–174. [Google Scholar] [CrossRef]
- Ng, P.C. Diagnostic markers of infection in neonates. Arch. Dis. Child Fetal Neonatal. 2004, 89, F229–F235. [Google Scholar] [CrossRef] [Green Version]
- Renwick, V.L.; Stewart, C.J. Exploring functional metabolites in preterm infants. Acta Paediatr. 2022, 111, 45–53. [Google Scholar] [CrossRef]
- Goenka, A.; Kollmann, T.R. Development of immunity in early life. J. Infect. 2015, 71 (Suppl. 1), S112–S120. [Google Scholar] [CrossRef]
- Dias, M.L.; O’Connor, K.M.; Dempsey, E.M.; O’Halloran, K.D.; McDonald, F.B. Targeting the Toll-like receptor pathway as a therapeutic strategy for neonatal infection. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 321, R879–R902. [Google Scholar] [CrossRef]
- Wynn, J.L.; Levy, O. Role of innate host defenses in susceptibility to early-onset neonatal sepsis. Clin. Perinatol. 2010, 37, 307–337. [Google Scholar] [CrossRef] [Green Version]
- Ng, P.C.; Li, K.; Wong, R.P.; Chui, K.; Wong, E.; Li, G.; Fok, T.F. Proinflammatory and anti-inflammatory cytokine responses in preterm infants with systemic infections. Arch. Dis. Child Fetal Neonatal. 2003, 88, F209–F213. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.D.; Ji, T.T.; Dong, J.R.; Feng, H.; Chen, F.Q.; Chen, X.; Zhao, H.Y.; Chen, D.K.; Ma, W.T. Pathogenesis and Treatment of Cytokine Storm Induced by Infectious Diseases. Int. J. Mol. Sci. 2021, 22, 13009. [Google Scholar] [CrossRef]
- Sadeghi, K.; Berger, A.; Langgartner, M.; Prusa, A.R.; Hayde, M.; Herkner, K.; Pollak, A.; Spittler, A.; Forster-Waldl, E. Immaturity of infection control in preterm and term newborns is associated with impaired toll-like receptor signaling. J. Infect. Dis. 2007, 195, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.M.; Osuchowski, M.F.; Wang, J.H.; Pan, Z.K. Editorial: Immune Dysfunction: An Update of New Immune Cell Subsets and Cytokines in Sepsis. Front. Immunol. 2021, 12, 822068. [Google Scholar] [CrossRef]
- Hickey, M.J.; Kubes, P. Intravascular immunity: The host-pathogen encounter in blood vessels. Nat. Rev. Immunol. 2009, 9, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Drossou, V.; Kanakoudi, F.; Diamanti, E.; Tzimouli, V.; Konstantinidis, T.; Germenis, A.; Kremenopoulos, G.; Katsougiannopoulos, V. Concentrations of main serum opsonins in early infancy. Arch. Dis. Child Fetal Neonatal. 1995, 72, F172–F175. [Google Scholar] [CrossRef] [Green Version]
- Koenig, J.M.; Stegner, J.J.; Schmeck, A.C.; Saxonhouse, M.A.; Kenigsberg, L.E. Neonatal neutrophils with prolonged survival exhibit enhanced inflammatory and cytotoxic responsiveness. Pediatr. Res. 2005, 57, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosmann, T.R. Cytokine secretion patterns and cross-regulation of T cell subsets. Immunol. Res. 1991, 10, 183–188. [Google Scholar] [CrossRef]
- Sriskandan, S.; Altmann, D.M. The immunology of sepsis. J. Pathol. 2008, 214, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Levy, O. Innate immunity of the newborn: Basic mechanisms and clinical correlates. Nat. Rev. Immunol. 2007, 7, 379–390. [Google Scholar] [CrossRef]
- Wong, H.R.; Shanley, T.P.; Sakthivel, B.; Cvijanovich, N.; Lin, R.; Allen, G.L.; Thomas, N.J.; Doctor, A.; Kalyanaraman, M.; Tofil, N.M.; et al. Genomics of Pediatric SIRS/Septic Shock Investigators. Genome-level expression profiles in pediatric septic shock indicate a role for altered zinc homeostasis in poor outcome. Physiol. Genom. 2007, 30, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.R.; Cvijanovich, N.; Lin, R.; Allen, G.L.; Thomas, N.J.; Willson, D.F.; Freishtat, R.J.; Anas, N.; Meyer, K.; Checchia, P.A.; et al. Identification of pediatric septic shock subclasses based on genome-wide expression profiling. BMC Med. 2009, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Cernada, M.; Serna, E.; Bauerl, C.; Collado, M.C.; Pérez-Martínez, G.; Vento, M. Genome-wide expression profiles in very low birth weight infants with neonatal sepsis. Pediatrics 2014, 133, e1203–e1211. [Google Scholar] [CrossRef] [Green Version]
- Wynn, J.L.; Guthrie, S.O.; Wong, H.R.; Lahni, P.; Ungaro, R.; Lopez, M.C.; Baker, H.V.; Moldawer, L.L. Postnatal Age Is a Critical Determinant of the Neonatal Host Response to Sepsis. Mol. Med. Camb. Mass. 2015, 21, 496–504. [Google Scholar] [CrossRef]
- Smith, C.L.; Dickinson, P.; Forster, T.; Craigon, M.; Ross, A.; Khondoker, M.R.; France, R.; Ivens, A.; Lynn, D.J.; Orme, J.; et al. Identification of a human neonatal immune-metabolic network associated with bacterial infection. Nat. Commun. 2014, 5, 4649. [Google Scholar] [CrossRef] [Green Version]
- Raymond, S.L.; López, M.C.; Baker, H.V.; Larson, S.D.; Efron, P.A.; Sweeney, T.E.; Khatri, P.; Moldawer, L.L.; Wynn, J.L. Unique transcriptomic response to sepsis is observed among patients of different age groups. PLoS ONE 2017, 12, e0184159. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, T.E.; Wynn, J.L.; Cernada, M.; Serna, E.; Wong, H.R.; Baker, H.V.; Vento, M.; Khatri, P. Validation of the Sepsis MetaScore for Diagnosis of Neonatal Sepsis. J. Pediatr. Infect. Dis. Soc. 2018, 7, 129–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollmann, T.R.; Levy, O.; Montgomery, R.R.; Goriely, S. Innate immune function by Toll-like receptors: Distinct responses in newborns and the elderly. Immunity 2012, 37, 771–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webber, R.J.; Sweet, R.M.; Webber, D.S. Circulating Microvesicle-Associated Inducible Nitric Oxide Synthase Is a Novel Therapeutic Target to Treat Sepsis: Current Status and Future Considerations. Int. J. Mol. Sci. 2021, 22, 13371. [Google Scholar] [CrossRef]
- Vance, J.K.; Rawson, T.W.; Povroznik, J.M.; Brundage, K.M.; Robinson, C.M. Myeloid-Derived Suppressor Cells Gain Suppressive Function during Neonatal Bacterial Sepsis. Int. J. Mol. Sci. 2021, 22, 7047. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.; Strunk, T.; Lee, A.H.; Gill, E.E.; Falsafi, R.; Woodman, T.; Hibbert, J.; Hancock, R.E.W.; Currie, A. Whole blood transcriptional responses of very preterm infants during late-onset sepsis. PLoS ONE 2020, 15, e0233841. [Google Scholar] [CrossRef]
- Talaei, K.; Garan, S.A.; Quintela, B.M.; Olufsen, M.S.; Cho, J.; Jahansooz, J.R.; Bhullar, P.K.; Suen, E.K.; Piszker, W.J.; Martins, N.R.B.; et al. A Mathematical Model of the Dynamics of Cytokine Expression and Human Immune Cell Activation in Response to the Pathogen Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2021, 11, 711153. [Google Scholar] [CrossRef] [PubMed]
- Cernada, M.; Pinilla-González, A.; Kuligowski, J.; Morales, J.M.; Lorente- Pozo, S.; Piñeiro-Ramos, J.D.; Parra-Lloraca, A.; Lara-Cantón, I.; Vento, M.; Serna, E. Transcriptome profiles discriminate between Gram-positive and Gram-negative sepsis in preterm neonates. Pediatr. Res. 2022, 91, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Feezor, R.J.; Oberholzer, C.; Baker, H.V.; Novick, D.; Rubinstein, M.; Moldawer, L.L.; Pribble, J.; Souza, S.; Dinarello, C.A.; Ertel, W.; et al. Molecular characterization of the acute inflammatory response to infections with gram-negative versus gram-positive bacteria. Infect. Immun. 2003, 71, 5803–5813. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Y.; Liu, Y.; Liu, Y.; Li, Y. Identification of key genes in Gram-positive and Gram-negative sepsis using stochastic perturbation. Mol. Med. Rep. 2017, 16, 3133–3146. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, X.; Tang, W.; Guan, X.; Xiong, Z.; Zhu, Y.; Gong, J.; Hu, B. Differential Gene Sets Profiling in Gram-Negative and Gram-Positive Sepsis. Front. Cell. Infect. Microbiol. 2022, 12, 801232. [Google Scholar] [CrossRef]
- Franco, J.H.; Chen, X.; Pan, Z.K. Novel Treatments Targeting the Dysregulated Cell Signaling Pathway during Sepsis. J. Cell Signal. 2021, 2, 228–234. [Google Scholar]
- Kwiatkowski, P.; Kurzawski, M.; Łopusiewicz, Ł.; Pruss, A.; Sienkiewicz, M.; Wojciechowska-Koszko, I.; Dołęgowska, B. Preliminary evaluation of selected inflammatory cytokine gene expression in lymphocytes isolated from whole human blood infected with trans-anethole-treated Staphylococcus aureus Newman strain. Lett. Appl. Microbiol. 2022, 74, 513–518. [Google Scholar] [CrossRef]
- Cao, I.; Lippmann, N.; Thome, U.H. The Value of Perinatal Factors, Blood Biomarkers and Microbiological Colonization Screening in Predicting Neonatal Sepsis. J. Clin. Med. 2022, 11, 5837. [Google Scholar] [CrossRef]
- Eichberger, J.; Resch, E.; Resch, B. Diagnosis of Neonatal Sepsis: The Role of Inflammatory Markers. Front. Pediatr. 2022, 10, 840288. [Google Scholar] [CrossRef]
- Li, T.; Li, X.; Liu, X.; Zhu, Z.; Zhang, M.; Xu, Z.; Wei, Y.; Feng, Y.; Qiao, X.; Yang, J.; et al. Association of Procalcitonin to Albumin Ratio with the Presence and Severity of Sepsis in Neonates. J. Inflamm. Res. 2022, 15, 2313–2321. [Google Scholar] [CrossRef]
- Mathur, N.B.; Behera, B. Blood Procalcitonin Levels and Duration of Antibiotics in Neonatal Sepsis. J. Trop. Pediatr. 2019, 65, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Fugit, R.V.; McCoury, J.B.M.; Bessesen, M.T. Procalcitonin for sepsis management: Implementation within an antimicrobial stewardship program. Am. J. Health Syst. Pharm. 2022, zxac341. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Torres-Cuevas, I.; Parra-Llorca, A.; Sánchez-Illana, A.; Nuñez-Ramiro, A.; Kuligowski, J.; Cháfer-Pericás, C.; Cernada, M.; Escobar, J.; Vento, M. Oxygen and oxidative stress in the perinatal period. Redox Biol. 2017, 12, 674–681. [Google Scholar] [CrossRef]
- Vera, S.; Martínez, R.; Gormaz, J.G.; Gajardo, A.; Galleguillos, F.; Rodrigo, R. Novel relationships between oxidative stress and angiogenesis-related factors in sepsis: New biomarkers and therapies. Ann. Med. 2015, 47, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive Oxygen Species and Neutrophil Function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef]
- Neri, M.; Riezzo, I.; Pomara, C.; Schiavone, S.; Turillazzi, E. Oxidative-Nitrosative Stress and Myocardial Dysfunctions in Sepsis: Evidence from the Literature and Postmortem Observations. Mediat. Inflamm. 2016, 2016, 3423450. [Google Scholar] [CrossRef] [Green Version]
- Reczek, C.R.; Chandel, N.S. ROS-dependent signal transduction. Curr. Opin. Cell Biol. 2015, 33, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Cebula, M.; Schmidt, E.E.; Arnér, E.S.J. TrxR1 as a potent regulator of the Nrf2-Keap1 response system. Antioxid. Redox Signal. 2015, 23, 823–853. [Google Scholar] [CrossRef] [Green Version]
- Marinho, H.S.; Real, C.; Cyrne, L.; Soares, H.; Antunes, F. Hydrogen peroxide sensing, signaling and regulation of transcription factors. Redox Biol. 2014, 2, 535–562. [Google Scholar] [CrossRef] [Green Version]
- Zambrano, S.; De Toma, I.; Piffer, A.; Bianchi, M.E.; Agresti, A. NF-κB oscillations translate into functionally related patterns of gene expression. eLife 2016, 5, e09100. [Google Scholar] [CrossRef] [PubMed]
- Vento, M.; Aguar, M.; Escobar, J.; Arduini, A.; Escrig, R.; Brugada, M.; Izquierdo, I.; Asensi, M.A.; Sastre, J.; Saenz, P.; et al. Antenatal steroids and antioxidant enzyme activity in preterm infants: Influence of gender and timing. Antioxid. Redox Signal. 2009, 11, 2945–2955. [Google Scholar] [CrossRef]
- Luce, W.A.; Hoffman, T.M.; Bauer, J.A. Bench-to-bedside review: Developmental influences on the mechanisms, treatment and outcomes of cardiovascular dysfunction in neonatal versus adult sepsis. Crit. Care 2007, 11, 228. [Google Scholar] [CrossRef] [Green Version]
- Bajčetić, M.; Spasić, S.; Spasojević, I. Redox therapy in neonatal sepsis: Reasons, targets, strategy, and agents. Shock (Augusta Ga.) 2014, 42, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Cernada, M.; Bäuerl, C.; Serna, E.; Collado, M.C.; Martínez, G.P.; Vento, M. Sepsis in preterm infants causes alterations in mucosal gene expression and microbiota profiles compared to non-septic twins. Sci. Rep. 2016, 6, 25497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asci, A.; Surmeli-Onay, O.; Erkekoglu, P.; Yigit, S.; Yurdakok, M.; Kocer-Gumusel, B. Oxidant and antioxidant status in neonatal proven and clinical sepsis according to selenium status. Pediatr. Int. J. Jpn. Pediatr. Soc. 2015, 57, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Cancelier, A.C.; Petronilho, F.; Reinke, A.; Constantino, L.; Machado, R.; Ritter, C.; Dal-Pizzol, F. Inflammatory and oxidative parameters in cord blood as diagnostic of early-onset neonatal sepsis: A case-control study. Pediatr. Crit. Care Med. 2009, 10, 467–471. [Google Scholar] [CrossRef]
- Kuligowski, J.; Torres-Cuevas, I.; Quintás, G.; Rook, D.; van Goudoever, J.B.; Cubells, E.; Asensi, M.; Lliso, I.; Nuñez, A.; Vento, M.; et al. Assessment of oxidative damage to proteins and DNA in urine of newborn infants by a validated UPLC-MS/MS approach. PLoS ONE 2014, 9, e93703. [Google Scholar] [CrossRef] [Green Version]
- Pinilla-Gonzalez, A.; Lara-Cantón, I.; Torrejón-Rodríguez, L.; Parra-Llorca, A.; Aguar, M.; Kuligowski, J.; Piñeiro-Ramos, J.D.; Sánchez-Illana, Á.; Navarro, A.G.; Vento, M.; et al. Early molecular markers of ventilator-associated pneumonia in bronchoalveolar lavage in preterm infants. Pediatr. Res. 2022, 7, 1–7. [Google Scholar] [CrossRef]
- Kuligowski, J.; Escobar, J.; Quintás, G.; Lliso, I.; Torres-Cuevas, I.; Nuñez, A.; Cubells, E.; Rook, D.; van Goudoever, J.B.; Vento, M. Analysis of lipid peroxidation biomarkers in extremely low gestational age neonate urines by UPLC-MS/MS. Anal. Bioanal. Chem. 2014, 406, 4345–4356. [Google Scholar] [CrossRef]
- Kuligowski, J.; Aguar, M.; Rook, D.; Lliso, I.; Torres-Cuevas, I.; Escobar, J.; Quintás, G.; Brugada, M.; Sánchez-Illana, A.; van Goudoever, J.B.; et al. Urinary Lipid Peroxidation Byproducts: Are They Relevant for Predicting Neonatal Morbidity in Preterm Infants? Antioxid. Redox Signal. 2015, 23, 178–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, G.; Williams, T.C. Human milk for preterm infants: Why, what, when and how? Arch. Dis. Child Fetal Neonatal. 2013, 98, F559–F562. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, P. Clinical Benefits of Lactoferrin for Infants and Children. J. Pediatr. 2016, 173, S43–S52. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, J. Human milk and the premature infant. Ann. Nutr. Metab. 2013, 62 (Suppl. 3), 8–14. [Google Scholar] [CrossRef] [PubMed]
- Quigley, M.; Embleton, N.D.; McGuire, W. Formula versus donor breast milk for feeding preterm or low birth weight infants. Cochrane Database Syst. Rev. 2019, 7, CD002971. [Google Scholar] [CrossRef] [Green Version]
- Ballard, O.; Morrow, A.L. Human Milk Composition: Nutrients and Bioactive Factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Păduraru, L.; Dimitriu, D.C.; Avasiloaiei, A.L.; Moscalu, M.; Zonda, G.I.; Stamatin, M. Total antioxidant status in fresh and stored human milk from mothers of term and preterm neonates. Pediatr. Neonatol. 2018, 59, 600–605. [Google Scholar] [CrossRef] [Green Version]
- Parra-Llorca, A.; Gormaz, M.; Sánchez-Illana, A.; Piñeiro-Ramos, J.D.; Collado, M.C.; Serna, E.; Cernada, M.; Nuñez-Ramiro, A.; Ramón-Beltrán, A.; Oger, C.; et al. Does Pasteurized Donor Human Milk Efficiently Protect Preterm Infants Against Oxidative Stress? Antioxid. Redox. Signal. 2019, 31, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Ledo, A.; Arduini, A.; Asensi, M.A.; Sastre, J.; Escrig, R.; Brugada, M.; Aguar, M.; Saenz, P.; Vento, M. Human milk enhances antioxidant defenses against hydroxyl radical aggression in preterm infants. Am. J. Clin. Nutr. 2009, 89, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Pammi, M.; Suresh, G. Enteral lactoferrin supplementation for prevention of sepsis and necrotizing enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2017, 6, CD007137. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Health Dis. 2015, 26, 26050. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [Green Version]
- Song, S.J.; Dominguez-Bello, M.G.; Knight, R. How delivery mode and feeding can shape the bacterial community in the infant gut. CMAJ 2013, 185, 373–374. [Google Scholar] [CrossRef] [Green Version]
- Cortese, R.; Lu, L.; Yu, Y.; Ruden, D.; Claud, E.C. Epigenome-Microbiome crosstalk: A potential new paradigm influencing neonatal susceptibility to disease. Epigenetics 2016, 11, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Claud, E.C. Intrauterine Inflammation, Epigenetics, and Microbiome Influences on Preterm Infant Health. Curr. Pathobiol. Rep. 2018, 6, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Penders, J.; Stobberingh, E.E.; van den Brandt, P.A.; Thijs, C. The role of the intestinal microbiota in the development of atopic disorders. Allergy 2007, 62, 1223–1236. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, K.E.; Slusher, N.A.; Cabana, M.D.; Lynch, S.V. Role of the gut microbiota in defining human health. Expert Rev. Anti-Infect. Ther. 2010, 8, 435–454. [Google Scholar] [CrossRef] [Green Version]
- Mshvildadze, M.; Neu, J. The infant intestinal microbiome: Friend or foe? Early Hum. Dev. 2010, 86 (Suppl. 1), 67–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muglia, L.J.; Katz, M. The enigma of spontaneous preterm birth. N. Engl. J. Med. 2010, 362, 529–535. [Google Scholar] [CrossRef]
- Ruiz, L.; Moles, L.; Gueimonde, M.; Rodriguez, J.M. Perinatal Microbiomes’ Influence on Preterm Birth and Preterms’ Health: Influencing Factors and Modulation Strategies. J. Pediatr. Gastroenterol. Nutr. 2016, 63, e193–e203. [Google Scholar] [CrossRef] [PubMed]
- Mshvildadze, M.; Neu, J.; Shuster, J.; Theriaque, D.; Li, N.; Mai, V. Intestinal microbial ecology in premature infants assessed with non-culture-based techniques. J. Pediatr. 2010, 156, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Selma-Royo, M.; Tarrazó, M.; García-Mantrana, I.; Gómez-Gallego, C.; Salminen, S.; Collado, M.C. Shaping Microbiota During the First 1000 Days of Life. Adv. Exp. Med. Biol. 2019, 1125, 3–24. [Google Scholar]
- Ardissone, A.N.; de la Cruz, D.M.; Davis-Richardson, A.G.; Rechcigl, K.T.; Li, N.; Drew, J.C.; Murgas-Torrazza, R.; Sharma, R.; Hudak, M.L.; Triplett, E.W.; et al. Meconium microbiome analysis identifies bacteria correlated with premature birth. PLoS ONE 2014, 9, e90784. [Google Scholar] [CrossRef] [Green Version]
- Moles, L.; Gómez, M.; Heilig, H.; Bustos, G.; Fuentes, S.; de Vos, W.; Fernández, L.; Rodríguez, J.M.; Jiménez, E. Bacterial diversity in meconium of preterm neonates and evolution of their fecal microbiota during the first month of life. PLoS ONE 2013, 8, e66986. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.J.; Embleton, N.D.; Clements, E.; Luna, P.N.; Smith, D.P.; Fofanova, T.Y.; Nelson, A.; Taylor, G.; Orr, C.H.; Petrosino, J.F.; et al. Cesarean or Vaginal Birth Does Not Impact the Longitudinal Development of the Gut Microbiome in a Cohort of Exclusively Preterm Infants. Front. Microbiol. 2017, 8, 1008. [Google Scholar] [CrossRef] [Green Version]
- Arboleya, S.; Binetti, A.; Salazar, N.; Fernández, N.; Solís, G.; Hernández-Barranco, A.; Margolles, A.; de Los Reyes-Gavilán, C.G.; Gueimonde, M. Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol. Ecol. 2012, 79, 763–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arboleya, S.; Solís, G.; Fernández, N.; de los Reyes-Gavilán, C.G.; Gueimonde, M. Facultative to strict anaerobes ratio in the preterm infant microbiota: A target for intervention? Gut Microbes 2012, 3, 583–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquot, A.; Neveu, D.; Aujoulat, F.; Mercier, G.; Marchandin, H.; Jumas-Bilak, E.; Picaud, J.C. Dynamics and clinical evolution of bacterial gut microflora in extremely premature patients. J. Pediatr. 2011, 158, 390–396. [Google Scholar] [CrossRef]
- LaTuga, M.S.; Ellis, J.C.; Cotton, C.M.; Goldberg, R.N.; Wynn, J.L.; Jackson, R.B.; Seed, P.C. Beyond bacteria: A study of the enteric microbial consortium in extremely low birth weight infants. PLoS ONE 2011, 6, e27858. [Google Scholar] [CrossRef]
- Mai, V.; Torrazza, R.M.; Ukhanova, M.; Wang, X.; Sun, Y.; Li, N.; Shuster, J.; Sharma, R.; Hudak, M.L.; Neu, J. Distortions in development of intestinal microbiota associated with late onset sepsis in preterm infants. PLoS ONE 2013, 8, e52876. [Google Scholar] [CrossRef]
- Sherman, M.P. New concepts of microbial translocation in the neonatal intestine: Mechanisms and prevention. Clin. Perinatol. 2010, 37, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Deitch, E.A. Gut-origin sepsis: Evolution of a concept. Surg. J. R. Coll. Surg. Edinb. Irel. 2012, 10, 350–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemarkt, H.J.; De Meij, T.G.; van Ganzewinkel, C.J.; de Boer, N.K.H.; Andriessen, P.; Hütten, M.C.; Kramer, B.W. Necrotizing Enterocolitis, Gut Microbiota, and Brain Development: Role of the Brain-Gut Axis. Neonatology 2019, 115, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Pronovost, G.N.; Hsiao, E.Y. Perinatal interactions between the microbiome, immunity and neurodevelopment. Immunity 2019, 50, 18–36. [Google Scholar] [CrossRef]
- Möhle, L.; Mattei, D.; Heimesaat, M.M.; Bereswill, S.; Fischer, A.; Alutis, M.; French, T.; Hambardzumyan, D.; Matzinger, P.; Dunay, I.R.; et al. Ly6C(hi) Monocytes Provide a Link between Antibiotic-Induced Changes in Gut Microbiota and Adult Hippocampal Neurogenesis. Cell Rep. 2016, 15, 1945–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuong, H.E.; Yano, J.M.; Fung, T.C.; Hsiao, E.Y. The Microbiome and Host Behavior. Annu. Rev. Neurosci. 2017, 40, 21–49. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minter, M.R.; Hinterleitner, R.; Meisel, M.; Zhang, C.; Leone, V.; Zhang, X.; Oyler-Castrillo, P.; Zhang, X.; Musch, M.W.; Shen, X.; et al. Antibiotic-induced perturbations in microbial diversity during post-natal development alters amyloid pathology in an aged APPSWE/PS1ΔE9 murine model of Alzheimer’s disease. Sci. Rep. 2017, 7, 10411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackos, A.R.; Maltz, R.; Bailey, M.T. The role of the commensal microbiota in adaptive and maladaptive stressor-induced immunomodulation. Horm. Behav. 2017, 88, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Jašarević, E.; Howard, C.D.; Morrison, K.; Misic, A.; Weinkopff, T.; Scott, P.; Hunter, C.; Beiting, D.; Bale, T.L. The maternal vaginal microbiome partially mediates the effects of prenatal stress on offspring gut and hypothalamus. Nat. Neurosci. 2018, 21, 1061–1071. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef]
- Stewart, C.J.; Marrs, E.C.L.; Nelson, A.; Lanyon, C.; Perry, J.D.; Embleton, N.D.; Cummings, S.P.; Berrington, J.E. Development of the preterm gut microbiome in twins at risk of necrotising enterocolitis and sepsis. PLoS ONE 2013, 8, e73465. [Google Scholar] [CrossRef] [PubMed]
- Madan, J.C.; Salari, R.C.; Saxena, D.; Davidson, L.; O’Toole, G.A.; Moore, J.H.; Sogin, M.L.; Foster, J.A.; Edwards, W.H.; Palumbo, P.; et al. Gut microbial colonisation in premature neonates predicts neonatal sepsis. Arch. Dis. Child Fetal Neonatal. 2012, 97, F456–F462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pammi, M.; Cope, J.; Tarr, P.I.; Warner, B.B.; Morrow, A.L.; Mai, V.; Gregory, K.E.; Kroll, J.S.; McMurtry, V.; Ferris, M.J.; et al. Intestinal dysbiosis in preterm infants preceding necrotizing enterocolitis: A systematic review and meta-analysis. Microbiome 2017, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Taft, D.H.; Ambalavanan, N.; Schibler, K.R.; Yu, Z.; Newburg, D.S.; Deshmukh, H.; Ward, D.V.; Morrow, A.L. Center Variation in Intestinal Microbiota Prior to Late-Onset Sepsis in Preterm Infants. PLoS ONE 2015, 10, e0130604. [Google Scholar] [CrossRef] [PubMed]
- Raymond, F.; Déraspe, M.; Boissinot, M.; Bergeron, M.G.; Corbeil, J. Partial recovery of microbiomes after antibiotic treatment. Gut Microbes 2016, 7, 428–434. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, S.E.; Tobin, J.M.; Opie, G.F.; Donath, S.; Tabrizi, S.N.; Pirotta, M.; Morley, C.J.; Garland, S.M. ProPrems Study Group. Probiotic effects on late-onset sepsis in very preterm infants: A randomized controlled trial. Pediatrics 2013, 132, 1055–1062. [Google Scholar] [CrossRef] [Green Version]
- Robertson, C.; Savva, G.M.; Clapuci, R.; Jones, J.; Maimouni, H.; Brown, E.; Minocha, A.; Hall, L.J.; Clarke, P. Incidence of necrotising enterocolitis before and after introducing routine prophylactic Lactobacillus and Bifidobacterium probiotics. Arch. Dis. Child Fetal Neonatal. Ed. 2020, 105, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Granger, C.; Dermyshi, E.; Roberts, E.; Beck, L.C.; Embleton, N.; Berrington, J. Necrotising enterocolitis, late-onset sepsis and mortality after routine probiotic introduction in the UK. Arch. Dis. Child Fetal Neonatal. Ed. 2022, 107, 352–358. [Google Scholar] [CrossRef] [PubMed]
- van den Akker, C.H.P.; van Goudoever, J.B.; Szajewska, H.; Embleton, N.D.; Hojsak, I.; Reid, D.; Shamir, R.; ESPGHAN Working Group for Probiotics, Prebiotics & Committee on Nutrition. Probiotics for Preterm Infants: A Strain-Specific Systematic Review and Network Meta-analysis. J. Pediatr. Gastroenterol. Nutr. 2018, 67, 103–122. [Google Scholar] [CrossRef]
- Wandro, S.; Osborne, S.; Enriquez, C.; Bixby, C.; Arrieta, A.; Whiteson, K. The Microbiome and Metabolome of Preterm Infant Stool Are Personalized and Not Driven by Health Outcomes, Including Necrotizing Enterocolitis and Late-Onset Sepsis. mSphere 2018, 3, e00104–e00118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitkin, S.I.; Tkachenko, E.I.; Vakhitov, T.Y. Metabolic dysbiosis of the gut microbiota and its biomarkers. Eksp Klin Gastroenterol. 2016, 12, 6–29. [Google Scholar] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parra-Llorca, A.; Pinilla-Gonzlez, A.; Torrejón-Rodríguez, L.; Lara-Cantón, I.; Kuligowski, J.; Collado, M.C.; Gormaz, M.; Aguar, M.; Vento, M.; Serna, E.; et al. Effects of Sepsis on Immune Response, Microbiome and Oxidative Metabolism in Preterm Infants. Children 2023, 10, 602. https://doi.org/10.3390/children10030602
Parra-Llorca A, Pinilla-Gonzlez A, Torrejón-Rodríguez L, Lara-Cantón I, Kuligowski J, Collado MC, Gormaz M, Aguar M, Vento M, Serna E, et al. Effects of Sepsis on Immune Response, Microbiome and Oxidative Metabolism in Preterm Infants. Children. 2023; 10(3):602. https://doi.org/10.3390/children10030602
Chicago/Turabian StyleParra-Llorca, Anna, Alejandro Pinilla-Gonzlez, Laura Torrejón-Rodríguez, Inmaculada Lara-Cantón, Julia Kuligowski, María Carmen Collado, María Gormaz, Marta Aguar, Máximo Vento, Eva Serna, and et al. 2023. "Effects of Sepsis on Immune Response, Microbiome and Oxidative Metabolism in Preterm Infants" Children 10, no. 3: 602. https://doi.org/10.3390/children10030602