
Antisense Oligonucleotide-Based Therapeutic against Menin for Triple-Negative Breast Cancer Treatment

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Design and Synthesis of Antisense Oligonucleotides
2.3. Transfection with ASOs
2.4. Western Blot
2.5. Cell Viability with Alamar Blue
2.6. Cell Treatment with Docetaxel and MTT Assay
2.7. Cell Cycle Distribution Assay
2.8. Cell Apoptosis Assay
2.9. Assessment of Menin Silencing in TNBC Xenografted Mice
2.10. Immunoprecipitation and Mass Spectrometry Analysis
2.11. Bioinformatics Analyses
2.12. Statistical Analysis
3. Results
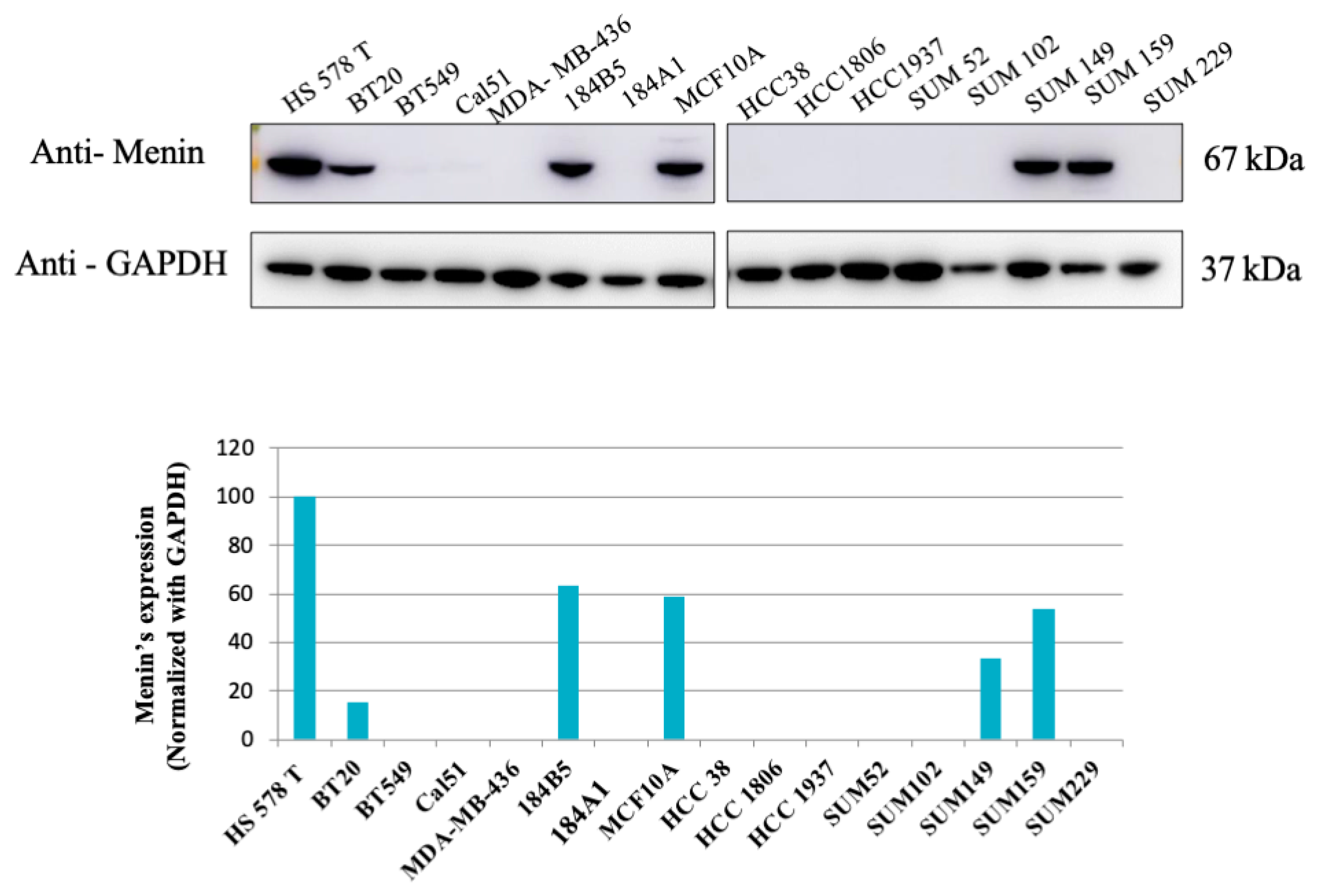
3.1. Expression of Menin in Different Breast Cancer Cell Lines
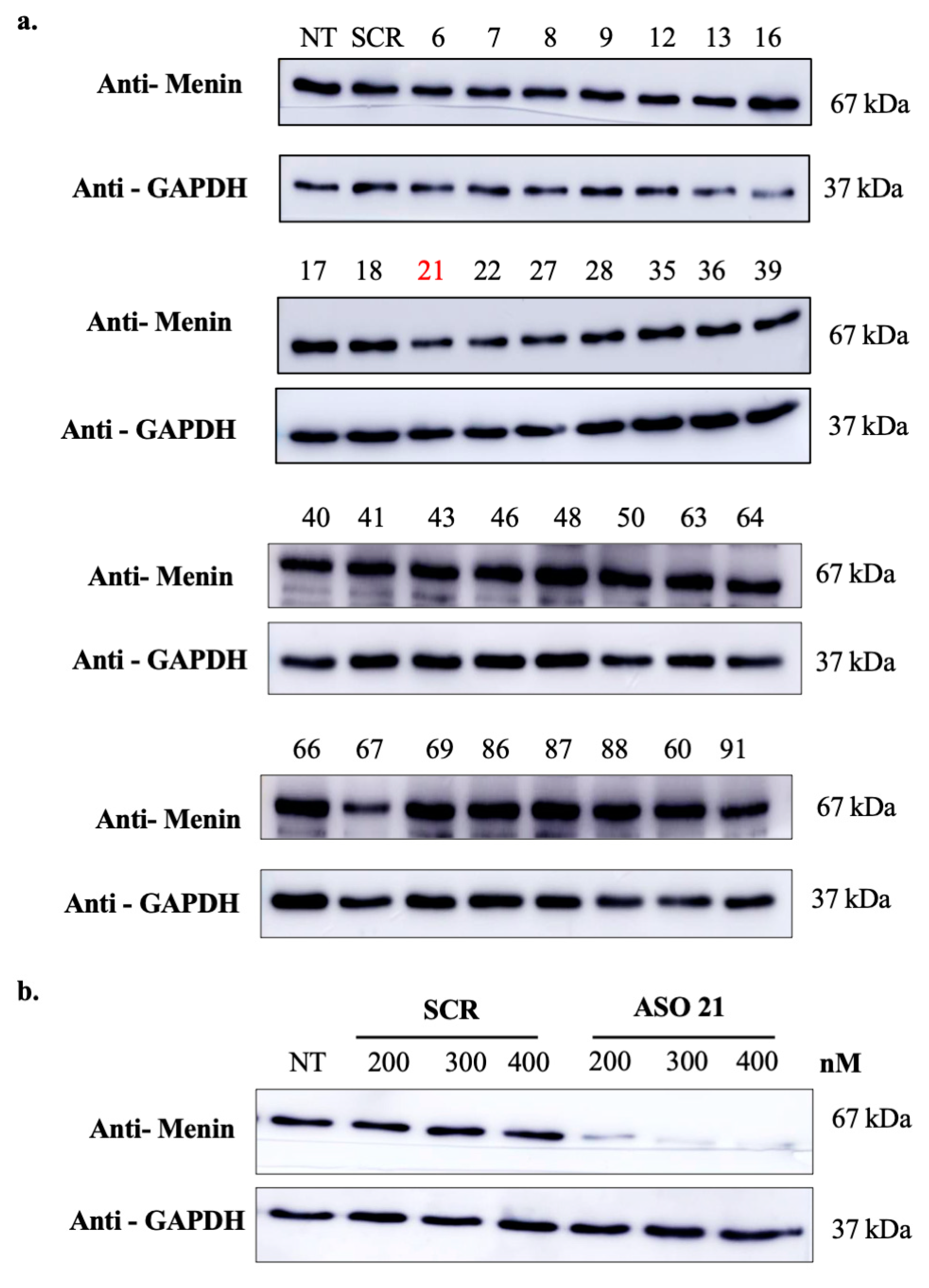
3.2. Design and Screening of Menin-ASOs
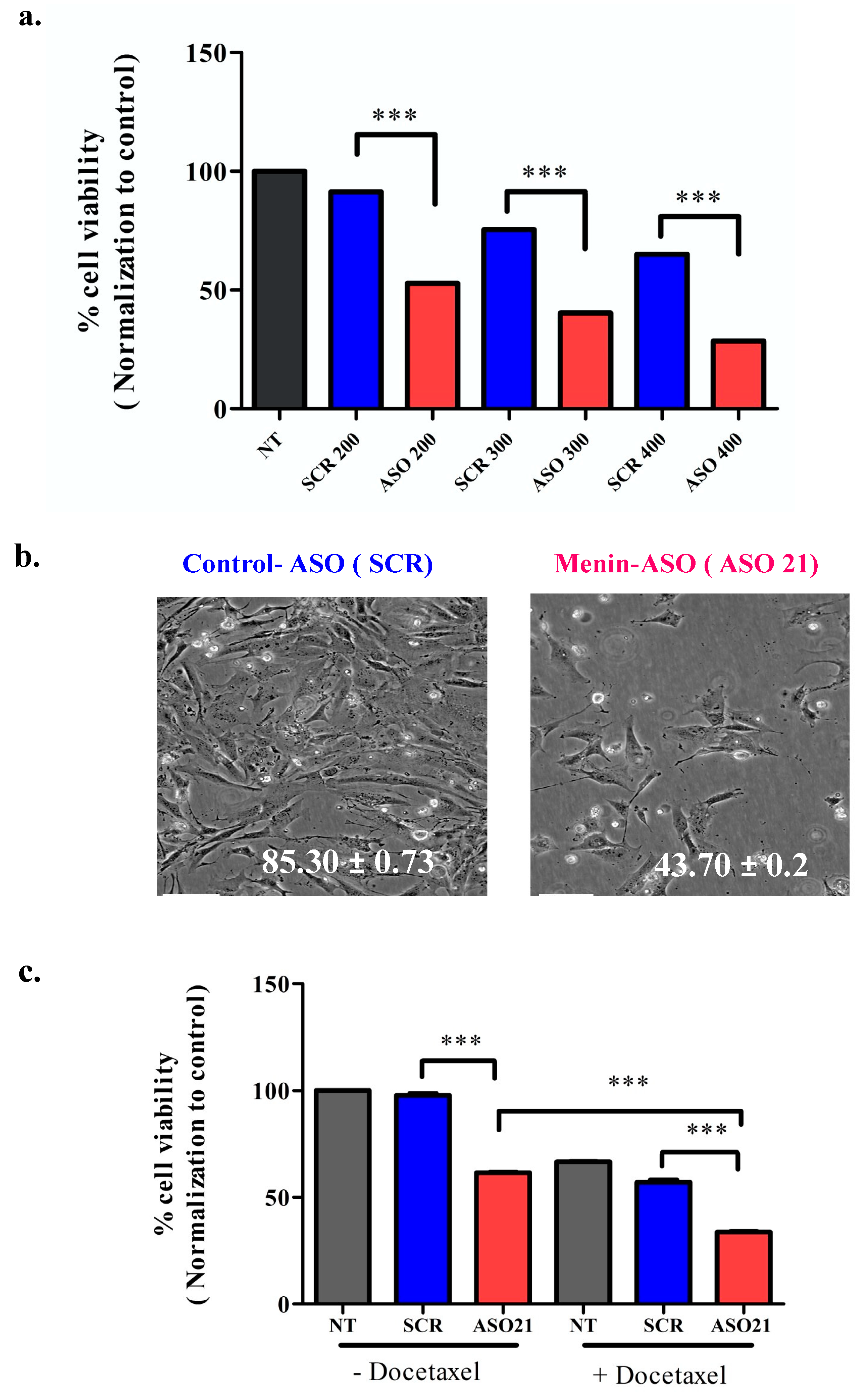
3.3. ASO21 Reduces Hs 578T Viability and Enhances Chemosensitivity of TNBC
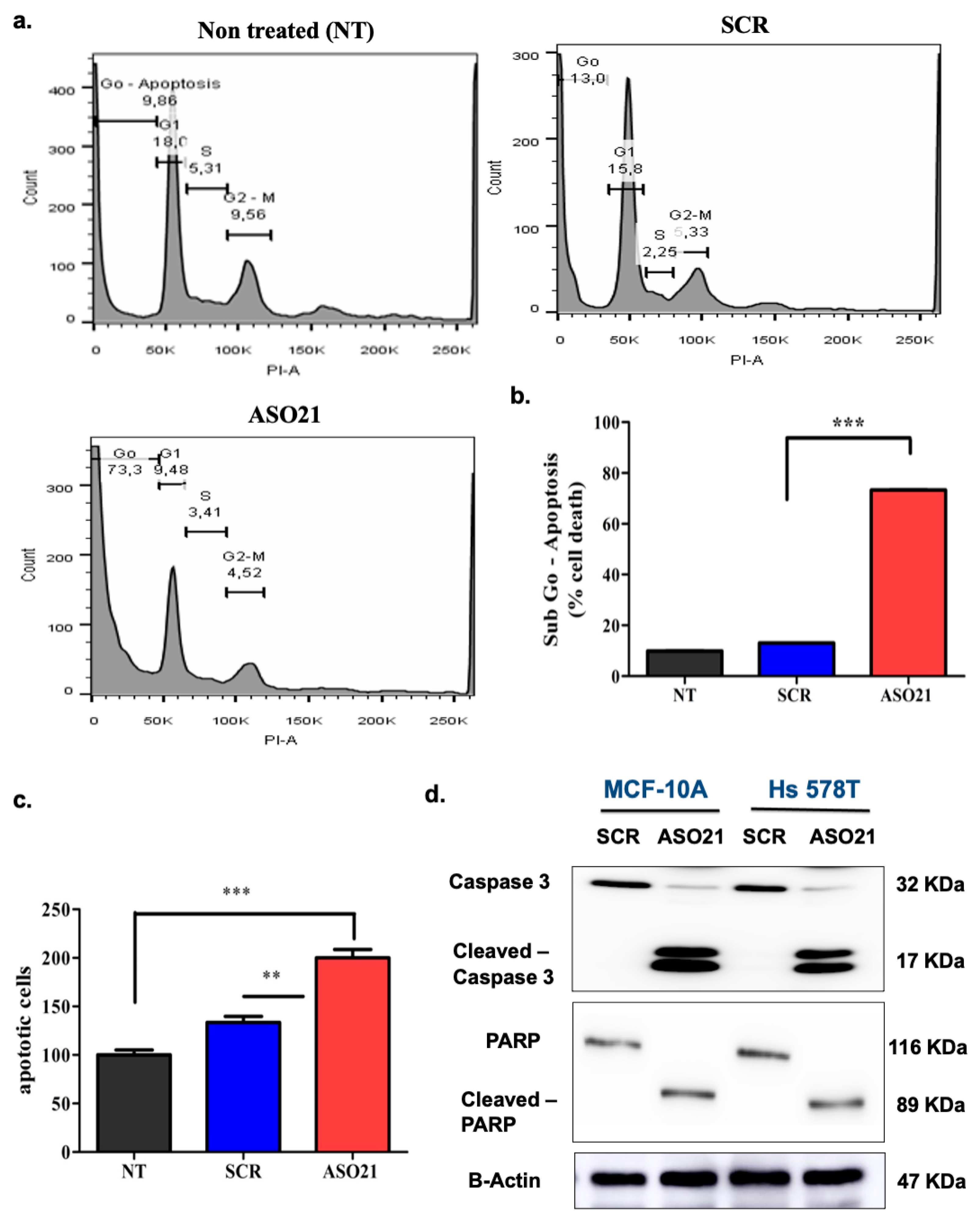
3.4. ASO21 Increases Tumor Cell Apoptosis via the Intrinsic Pathway
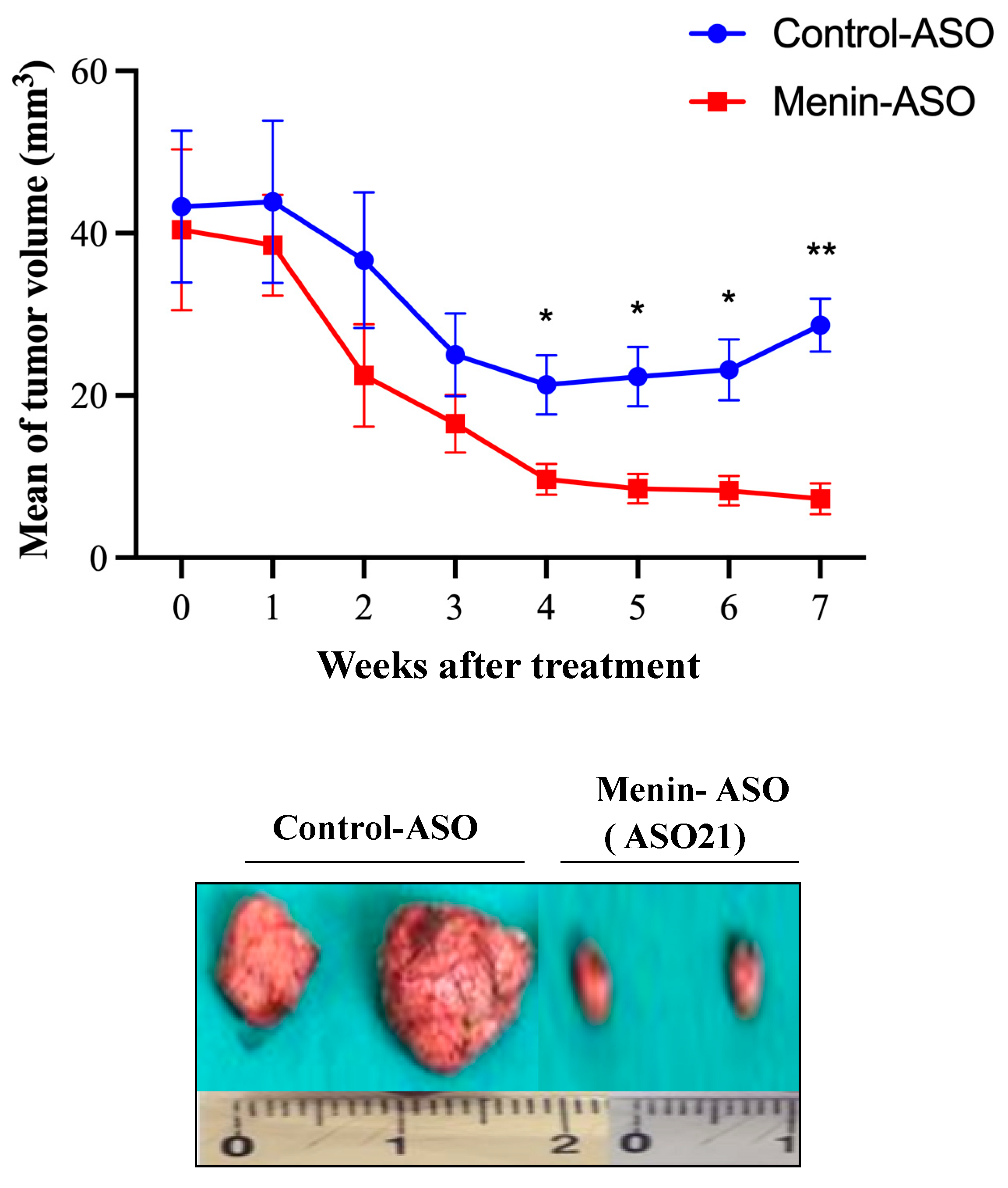
3.5. Menin-ASO (ASO21) Reduces TNBC-Derived Xenograft Tumor Growth
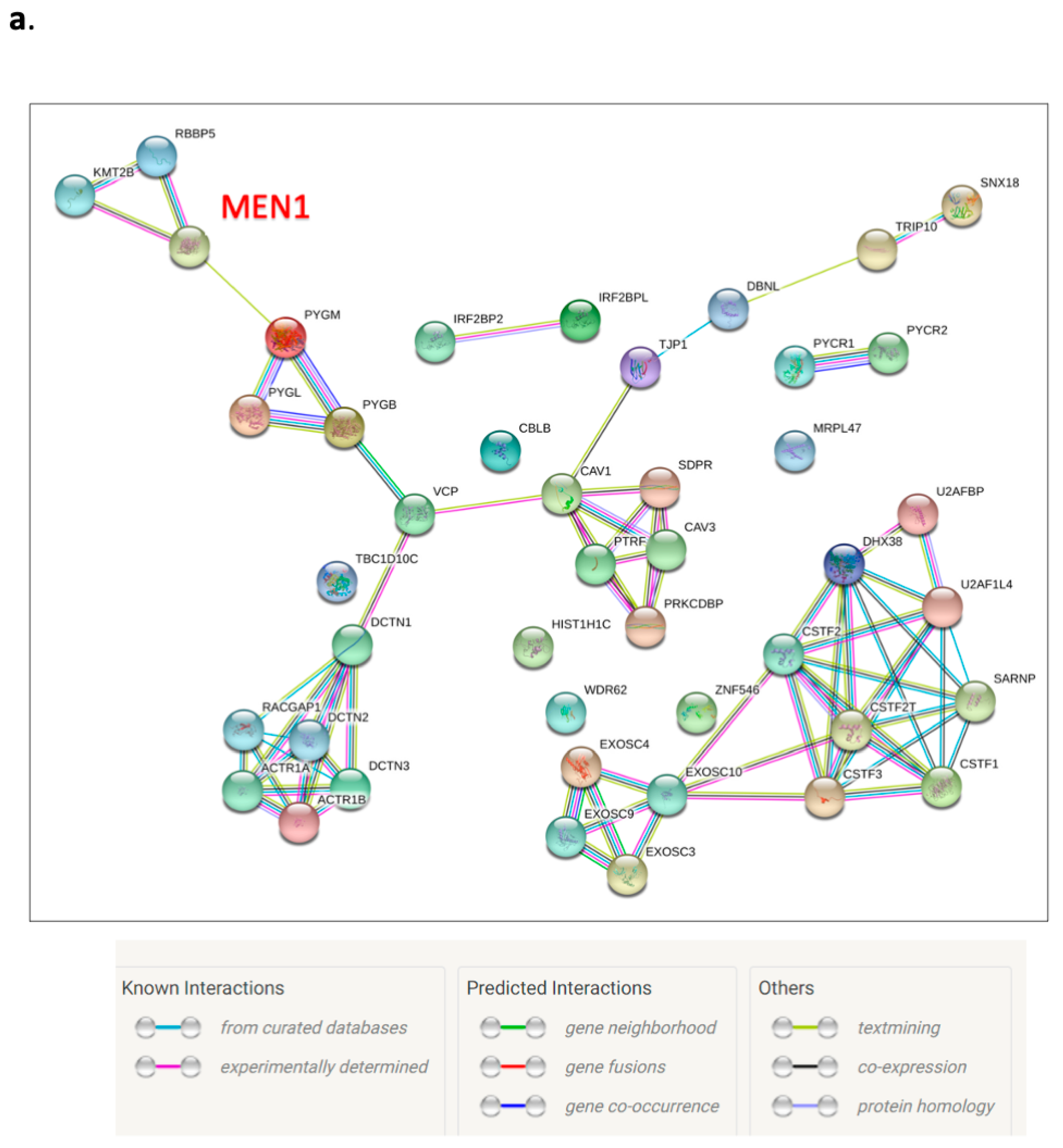
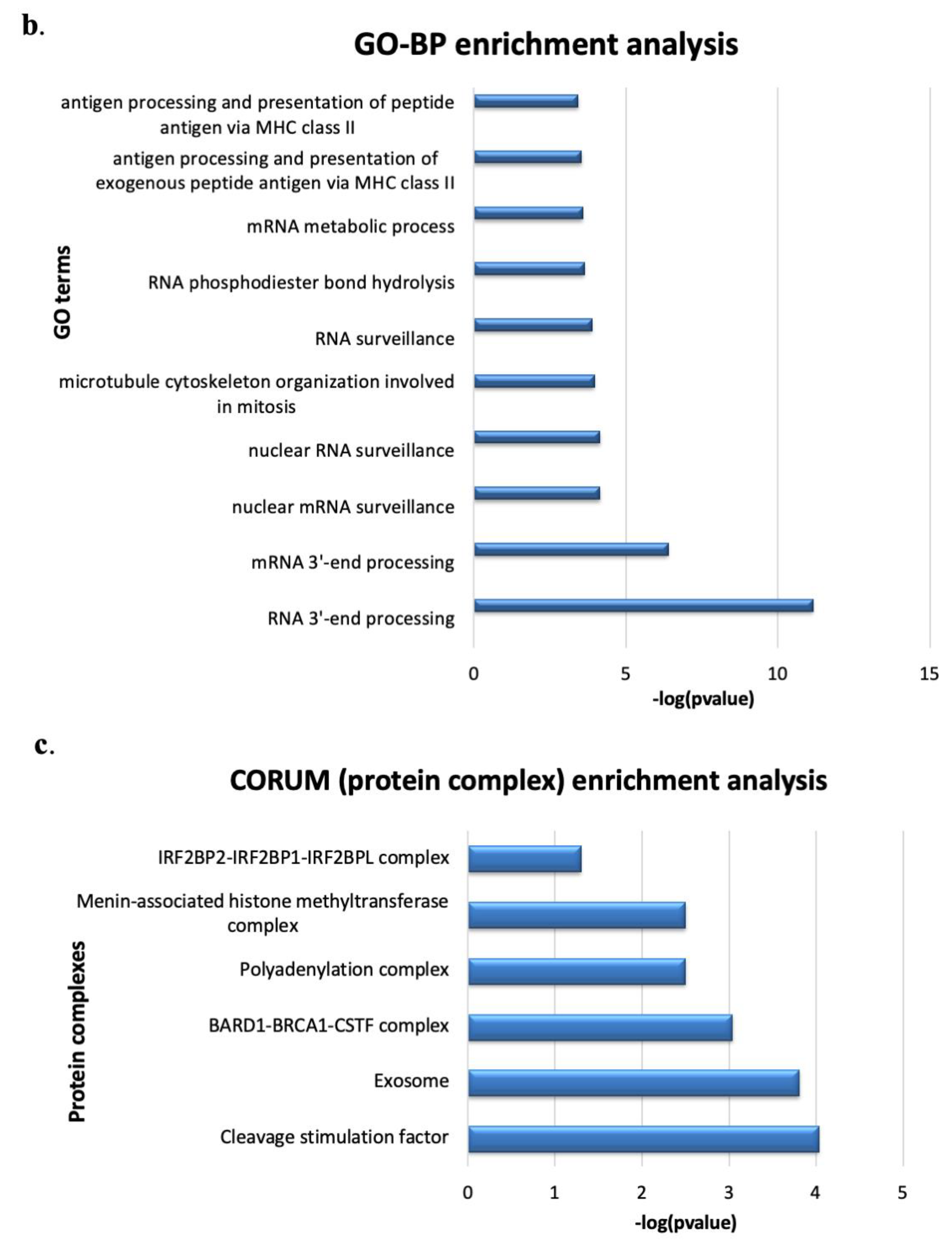
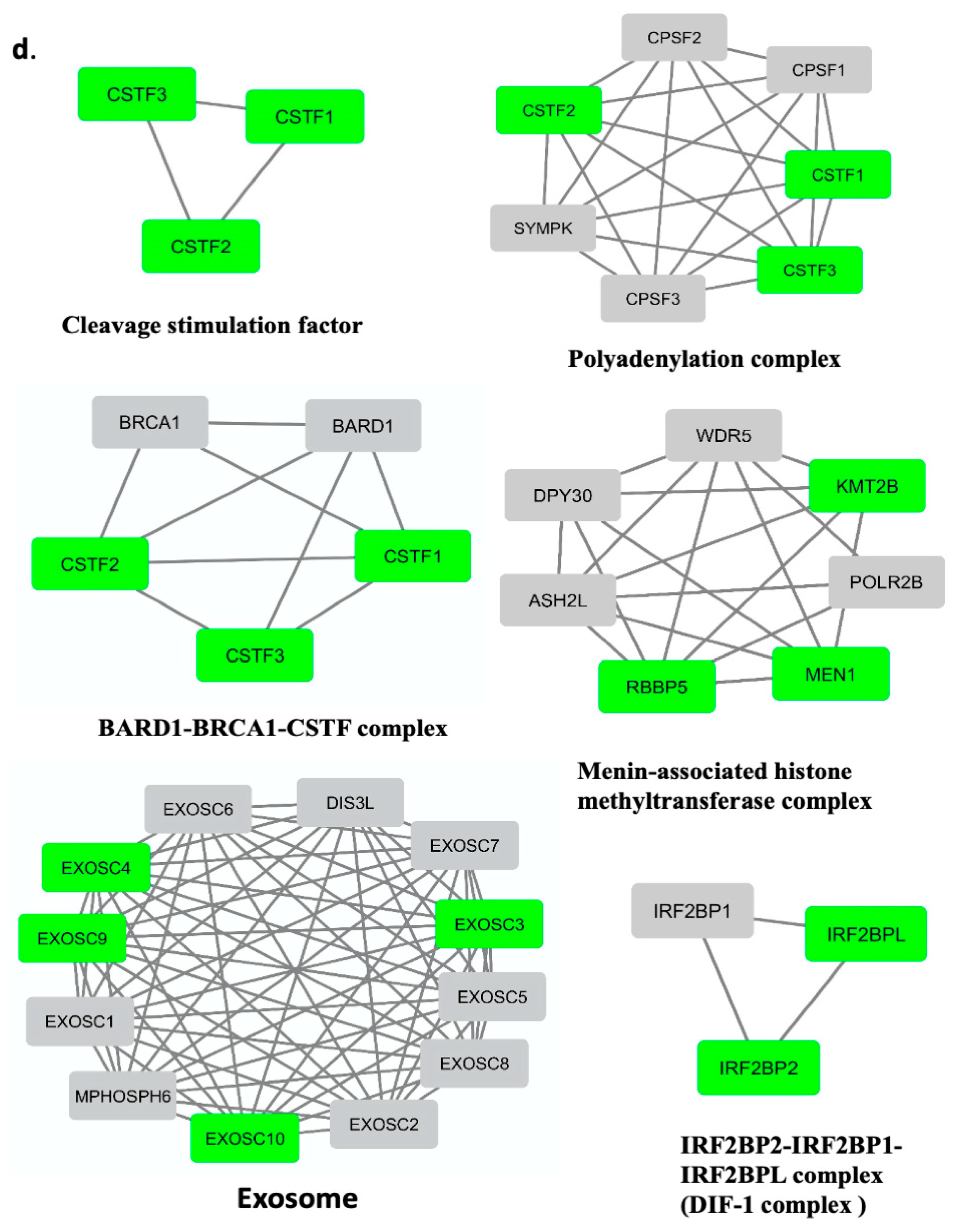
3.6. Menin’s Interactome Implicates New Putative Functions in TNBC Hs 578T
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef]
- Allred, D.C. Issues and updates: Evaluating estrogen receptor-α, progesterone receptor, and HER2 in breast cancer. Mod. Pathol. 2010, 23, S52–S59. [Google Scholar] [CrossRef] [Green Version]
- Fragomeni, S.M.; Sciallis, A.; Jeruss, J.S. Molecular Subtypes and Local-Regional Control of Breast Cancer. Surg. Oncol. Clin. N. Am. 2018, 27, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-Q.; Russo, J. ERα-negative and triple negative breast cancer: Molecular features and potential therapeutic approaches. Biochim. Biophys. Acta (BBA)-Bioenerg. 2009, 1796, 162–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S.-C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 1–13. [Google Scholar] [CrossRef]
- Foulkes, W.D.; Smith, I.E.; Reis-Filho, J.S. Triple-Negative Breast Cancer. N. Engl. J. Med. 2010, 363, 1938–1948. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Castro, A.C.; Lin, N.U.; Polyak, K. Insights into Molecular Classifications of Triple-Negative Breast Cancer: Improving Patient Selection for Treatment. Cancer Discov. 2019, 9, 176–198. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Yang, J.; Peng, L.; Sahin, A.A.; Huo, L.; Ward, K.C.; O’Regan, R.; Torres, M.A.; Meisel, J.L. Triple-negative breast cancer has worse overall survival and cause-specific survival than non-triple-negative breast cancer. Breast Cancer Res. Treat. 2017, 161, 279–287. [Google Scholar] [CrossRef]
- Cierpicki, T.; Grembecka, J. Challenges and opportunities in targeting the menin–MLL interaction. Future Med. Chem. 2014, 6, 447–462. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekharappa, S.C.; Guru, S.C.; Manickam, P.; Olufemi, S.-E.; Collins, F.S.; Emmert-Buck, M.R.; Debelenko, L.V.; Zhuang, Z.; Lubensky, I.A.; Liotta, L.A.; et al. Positional Cloning of the Gene for Multiple Endocrine Neoplasia-Type 1. Science 1997, 276, 404–407. [Google Scholar] [CrossRef]
- Lemmens, I.; Van De Ven, W.J.M.; Kas, K.; Zhang, C.X.; Giraud, S.; Wautot, V.; Buisson, N.; De Witte, K.; Salandre, J.; Lenoir, G.; et al. Identification of the multiple endocrine neoplasia type 1 (MEN1) gene. The European Consortium on MEN1. Hum. Mol. Genet. 1997, 6, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, J.S.; Scacheri, P.C.; Ward, J.M.; Garrett-Beal, L.; Emmert-Buck, M.R.; Edgemon, K.A.; Lorang, D.; Libutti, S.K.; Chandrasekharappa, S.C.; Marx, S.J.; et al. A mouse model of multiple endocrine neoplasia, type 1, develops multiple endocrine tumors. Proc. Natl. Acad. Sci. USA 2001, 98, 1118–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caslini, C.; Yang, Z.; El-Osta, M.; Milne, T.; Slany, R.K.; Hess, J.L. Interaction of MLL Amino Terminal Sequences with Menin Is Required for Transformation. Cancer Res. 2007, 67, 7275–7283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreijerink, K.; Groner, A.C.; Vos, E.; Font-Tello, A.; Gu, L.; Chi, D.; Reyes, J.; Cook, J.; Lim, E.; Lin, C.Y.; et al. Enhancer-Mediated Oncogenic Function of the Menin Tumor Suppressor in Breast Cancer. Cell Rep. 2017, 18, 2359–2372. [Google Scholar] [CrossRef] [Green Version]
- Dreijerink, K.M.; Goudet, P.; Burgess, J.; Valk, G.D. Breast-Cancer Predisposition in Multiple Endocrine Neoplasia Type 1. N. Engl. J. Med. 2014, 371, 583–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreijerink, K.; Mulder, K.W.; Winkler, S.; Höppener, J.W.; Lips, C.J.; Timmers, H.M. Menin Links Estrogen Receptor Activation to Histone H3K4 Trimethylation. Cancer Res. 2006, 66, 4929–4935. [Google Scholar] [CrossRef] [Green Version]
- Imachi, H.; Murao, K.; Dobashi, H.; Bhuyan, M.M.; Cao, X.; Kontani, K.; Niki, S.; Murazawa, C.; Nakajima, H.; Kohno, N.; et al. Menin, a product of the MENI gene, binds to estrogen receptor to enhance its activity in breast cancer cells: Possibility of a novel predictive factor for tamoxifen resistance. Breast Cancer Res. Treat. 2009, 122, 395–407. [Google Scholar] [CrossRef]
- Huang, J.; Gurung, B.; Wan, B.; Matkar, S.; Veniaminova, N.A.; Wan, K.; Merchant, J.L.; Hua, X.; Lei, M. The same pocket in menin binds both MLL and JUND but has opposite effects on transcription. Nature 2012, 482, 542–546. [Google Scholar] [CrossRef]
- Hughes, C.M.; Rozenblatt-Rosen, O.; Milne, T.; Copeland, T.D.; Levine, S.; Lee, J.C.; Hayes, D.N.; Shanmugam, K.S.; Bhattacharjee, A.; Biondi, C.A.; et al. Menin Associates with a Trithorax Family Histone Methyltransferase Complex and with the Hoxc8 Locus. Mol. Cell 2004, 13, 587–597. [Google Scholar] [CrossRef]
- Yokoyama, A.; Wang, Z.; Wysocka, J.; Sanyal, M.; Aufiero, D.J.; Kitabayashi, I.; Herr, W.; Cleary, M.L. Leukemia Proto-Oncoprotein MLL Forms a SET1-Like Histone Methyltransferase Complex with Menin To Regulate Hox Gene Expression. Mol. Cell. Biol. 2004, 24, 5639–5649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreijerink, K.; Timmers, H.T.M.; Brown, M.; Timmers, M. Twenty years of menin: Emerging opportunities for restoration of transcriptional regulation in MEN1. Endocr.-Relat. Cancer 2017, 24, T135–T145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Hu, C.; El Achkar, C.M.; Black, L.E.; Douville, J.; Larson, A.; Pendergast, M.K.; Goldkind, S.F.; Lee, E.A.; Kuniholm, A.; et al. Patient-Customized Oligonucleotide Therapy for a Rare Genetic Disease. N. Engl. J. Med. 2019, 381, 1644–1652. [Google Scholar] [CrossRef]
- Le, T.K.; Paris, C.; Khan, K.S.; Robson, F.; Ng, W.-L.; Rocchi, P. Nucleic Acid-Based Technologies Targeting Coronaviruses. Trends Biochem. Sci. 2021, 46, 351–365. [Google Scholar] [CrossRef]
- Grembecka, J.; He, S.; Shi, A.; Purohit, T.; Muntean, A.G.; Sorenson, R.J.; Showalter, H.D.; Murai, M.J.; Belcher, A.M.; Hartley, T.; et al. Menin-MLL inhibitors reverse oncogenic activity of MLL fusion proteins in leukemia. Nat. Chem. Biol. 2012, 8, 277–284. [Google Scholar] [CrossRef]
- Brzezinka, K.; Nevedomskaya, E.; Lesche, R.; Haegebarth, A.; Ter Laak, A.; Fernández-Montalván, A.E.; Eberspaecher, U.; Werbeck, N.D.; Moenning, U.; Siegel, S.; et al. Characterization of the Menin-MLL Interaction as Therapeutic Cancer Target. Cancers 2020, 12, 201. [Google Scholar] [CrossRef] [Green Version]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass Spectrometric Sequencing of Proteins from Silver-Stained Polyacrylamide Gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef]
- MaxQuant Enables High Peptide Identification Rates, Individualized p.p.b.-Range Mass Accuracies and Proteome-Wide Protein Quantification | Nature Biotechnology. Available online: https://www.nature.com/articles/nbt.1511 (accessed on 30 December 2020).
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in oligonucleotide drug delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef]
- Schnepp, R.W.; Mao, H.; Sykes, S.M.; Zong, W.-X.; Silva, A.; La, P.; Hua, X. Menin Induces Apoptosis in Murine Embryonic Fibroblasts. J. Biol. Chem. 2004, 279, 10685–10691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La, P.; Yang, Y.; Karnik, S.K.; Silva, A.C.; Schnepp, R.W.; Kim, S.K.; Hua, X. Menin-mediated Caspase 8 Expression in Suppressing Multiple Endocrine Neoplasia Type 1. J. Biol. Chem. 2007, 282, 31332–31340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazzi, W.; Renon, M.; Vercherat, C.; Hamze, Z.; Lacheretz–Bernigaud, A.; Wang, H.; Blanc, M.; Roche, C.; Calender, A.; Chayvialle, J.; et al. MEN1 Missense Mutations Impair Sensitization to Apoptosis Induced by Wild-Type Menin in Endocrine Pancreatic Tumor Cells. Gastroenterology 2008, 135, 1698–1709.e2. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Yeung, K.T.; Mahajan, M.A.; Samuels, H.H. Fas Activated Serine-Threonine Kinase Domains 2 (FASTKD2) mediates apoptosis of breast and prostate cancer cells through its novel FAST2 domain. BMC Cancer 2014, 14, 852. [Google Scholar] [CrossRef] [Green Version]
- Yeung, K.T.; Das, S.; Zhang, J.; Lomniczi, A.; Ojeda, S.R.; Xu, C.-F.; Neubert, T.A.; Samuels, H.H. A Novel Transcription Complex That Selectively Modulates Apoptosis of Breast Cancer Cells through Regulation of FASTKD2. Mol. Cell. Biol. 2011, 31, 2287–2298. [Google Scholar] [CrossRef] [Green Version]
- Ghezzi, D.; Saada, A.; D’Adamo, P.; Fernandez-Vizarra, E.; Gasparini, P.; Tiranti, V.; Elpeleg, O.; Zeviani, M. FASTKD2 Nonsense Mutation in an Infantile Mitochondrial Encephalomyopathy Associated with Cytochrome C Oxidase Deficiency. Am. J. Hum. Genet. 2008, 83, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Tinnikov, A.A.; Yeung, K.T.; Das, S.; Samuels, H.H. Identification of a novel pathway that selectively modulates apoptosis of breast cancer cells. Cancer Res. 2009, 69, 1375–1382. [Google Scholar] [CrossRef] [Green Version]
- Takagaki, Y.; Manley, J. Complex Protein Interactions within the Human Polyadenylation Machinery Identify a Novel Component. Mol. Cell. Biol. 2000, 20, 1515–1525. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hu, X.; Wang, P.; Shao, Z.-M. The 3′UTR signature defines a highly metastatic subgroup of triple-negative breast cancer. Oncotarget 2016, 7, 59834–59844. [Google Scholar] [CrossRef]
- Masamha, C.P.; Wagner, E.J. The contribution of alternative polyadenylation to the cancer phenotype. Carcinogenesis 2018, 39, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Xia, Z.; Donehower, L.A.; Cooper, T.A.; Neilson, J.R.; Wheeler, D.A.; Wagner, E.J.; Li, W. Dynamic analyses of alternative polyadenylation from RNA-seq reveal a 3′-UTR landscape across seven tumour types. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Hu, X.; Wang, P.; Shao, Z. Integrative 3′ Untranslated Region-Based Model to Identify Patients with Low Risk of Axillary Lymph Node Metastasis in Operable Triple-Negative Breast Cancer. Oncologist 2019, 24, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, D.; Xue, M.; Mi, X.; Liang, Y.; Wang, P. 3′UTR shortening identifies high-risk cancers with targeted dysregulation of the ceRNA network. Sci. Rep. 2014, 4, 5406. [Google Scholar] [CrossRef] [PubMed]
- Akman, H.B.; Oyken, M.; Tuncer, T.; Can, T.; Erson-Bensan, A.E. 3?UTR shortening and EGF signaling: Implications for breast cancer. Hum. Mol. Genet. 2015, 24, 6910–6920. [Google Scholar] [CrossRef] [PubMed]
- Staff, T.P.G.; Staff, P.G. Correction: 3’UTR Shortening Potentiates MicroRNA-Based Repression of Pro-differentiation Genes in Proliferating Human Cells. PLoS Genet. 2016, 12, e1006254. [Google Scholar] [CrossRef] [Green Version]
- Francis, J.; Lin, W.; Rozenblatt-Rosen, O.; Meyerson, M. The Menin Tumor Suppressor Protein Is Phosphorylated in Response to DNA Damage. PLoS ONE 2011, 6, e16119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Rege, M.; Peterson, C.L.; Volkert, M.R. RNA polymerase II depletion promotes transcription of alternative mRNA species. BMC Mol. Biol. 2016, 17, 20. [Google Scholar] [CrossRef] [Green Version]
- Denkert, C.; Liedtke, C.; Tutt, A.; von Minckwitz, G. Molecular alterations in triple-negative breast cancer—the road to new treatment strategies. Lancet 2017, 389, 2430–2442. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term_Name | Term_Id | Functions |
|---|---|---|
| Cleavage stimulation factor | CORUM:1146 | Cleavage step, mRNA 3′-end processing (polyadenylation) |
| BARD1-BRCA1-CSTF complex | CORUM:2211 | Interaction of BARD1 with CstF inhibits RNA polyadenylation and ensures that at DNA damage sites nascent RNAs are not erroneously polyadenylated. |
| Polyadenylation complex (CSTF1, CSTF2, CSTF3, SYMPK CPSF1, CPSF2, CPSF3) | CORUM:1147 | mRNA 3′-end processing (polyadenylation) |
| Menin-associated histone methyltransferase complex | CORUM:1254 | chromosome remodeling, epigenetic regulation |
| IRF2BP2-IRF2BP1-IRF2BPL complex | CORUM:6848 | regulation of apoptotic process, negative regulation of transcription |
| Exosome | CORUM:7443 | degradation and maturation of a wide variety of RNAs |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, D.T.; Le, T.K.; Paris, C.; Cherif, C.; Audebert, S.; Udu-Ituma, S.O.; Benizri, S.; Barthélémy, P.; Bertucci, F.; Taïeb, D.; et al. Antisense Oligonucleotide-Based Therapeutic against Menin for Triple-Negative Breast Cancer Treatment. Biomedicines 2021, 9, 795. https://doi.org/10.3390/biomedicines9070795
Nguyen DT, Le TK, Paris C, Cherif C, Audebert S, Udu-Ituma SO, Benizri S, Barthélémy P, Bertucci F, Taïeb D, et al. Antisense Oligonucleotide-Based Therapeutic against Menin for Triple-Negative Breast Cancer Treatment. Biomedicines. 2021; 9(7):795. https://doi.org/10.3390/biomedicines9070795
Chicago/Turabian StyleNguyen, Dang Tan, Thi Khanh Le, Clément Paris, Chaïma Cherif, Stéphane Audebert, Sandra Oluchi Udu-Ituma, Sébastien Benizri, Philippe Barthélémy, François Bertucci, David Taïeb, and et al. 2021. "Antisense Oligonucleotide-Based Therapeutic against Menin for Triple-Negative Breast Cancer Treatment" Biomedicines 9, no. 7: 795. https://doi.org/10.3390/biomedicines9070795