
MicroRNA-138 Increases Chemo-Sensitivity of Glioblastoma through Downregulation of Survivin

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Cell Cultures
2.2. Cell Proliferation Assay
2.3. Apoptosis Analysis
2.4. Western Blotting
2.5. Dual Luciferase Reporter Assay
2.6. Animal Studies
2.7. Statistical Analysis
3. Results
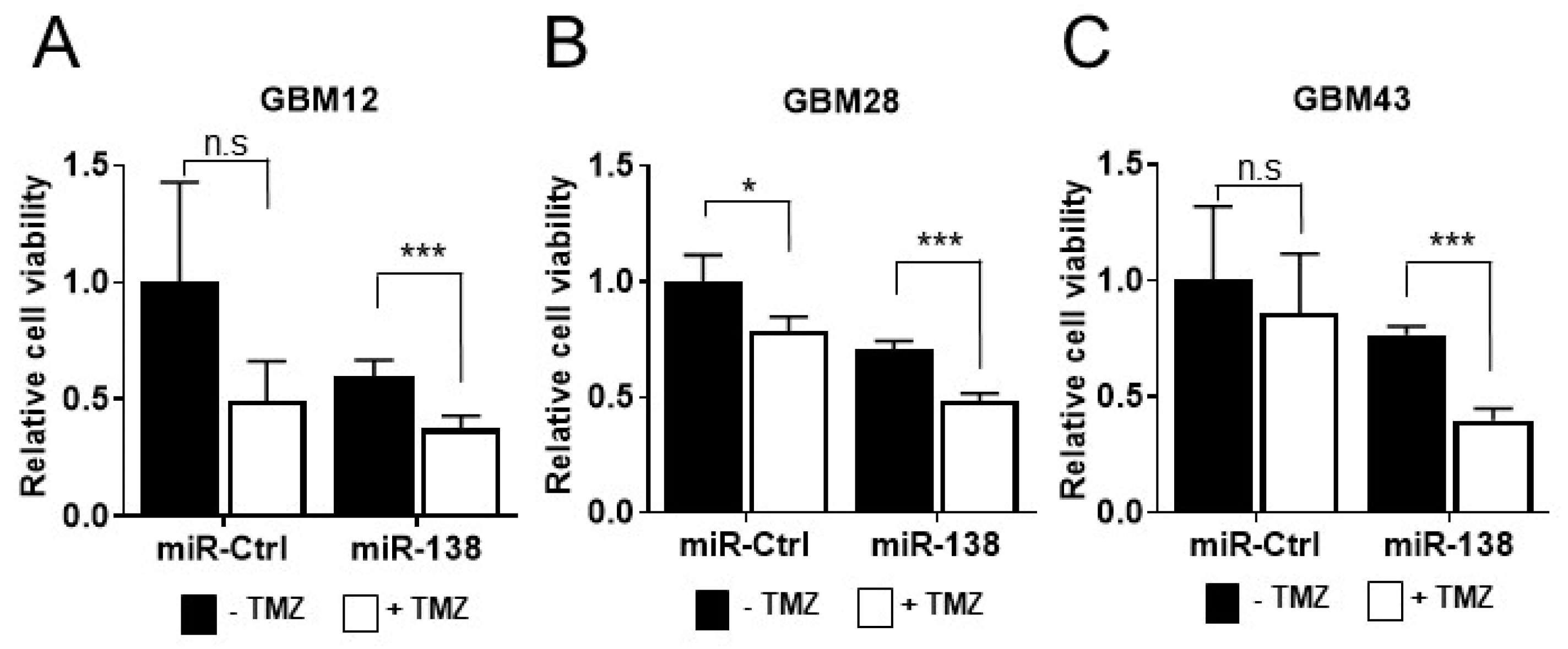
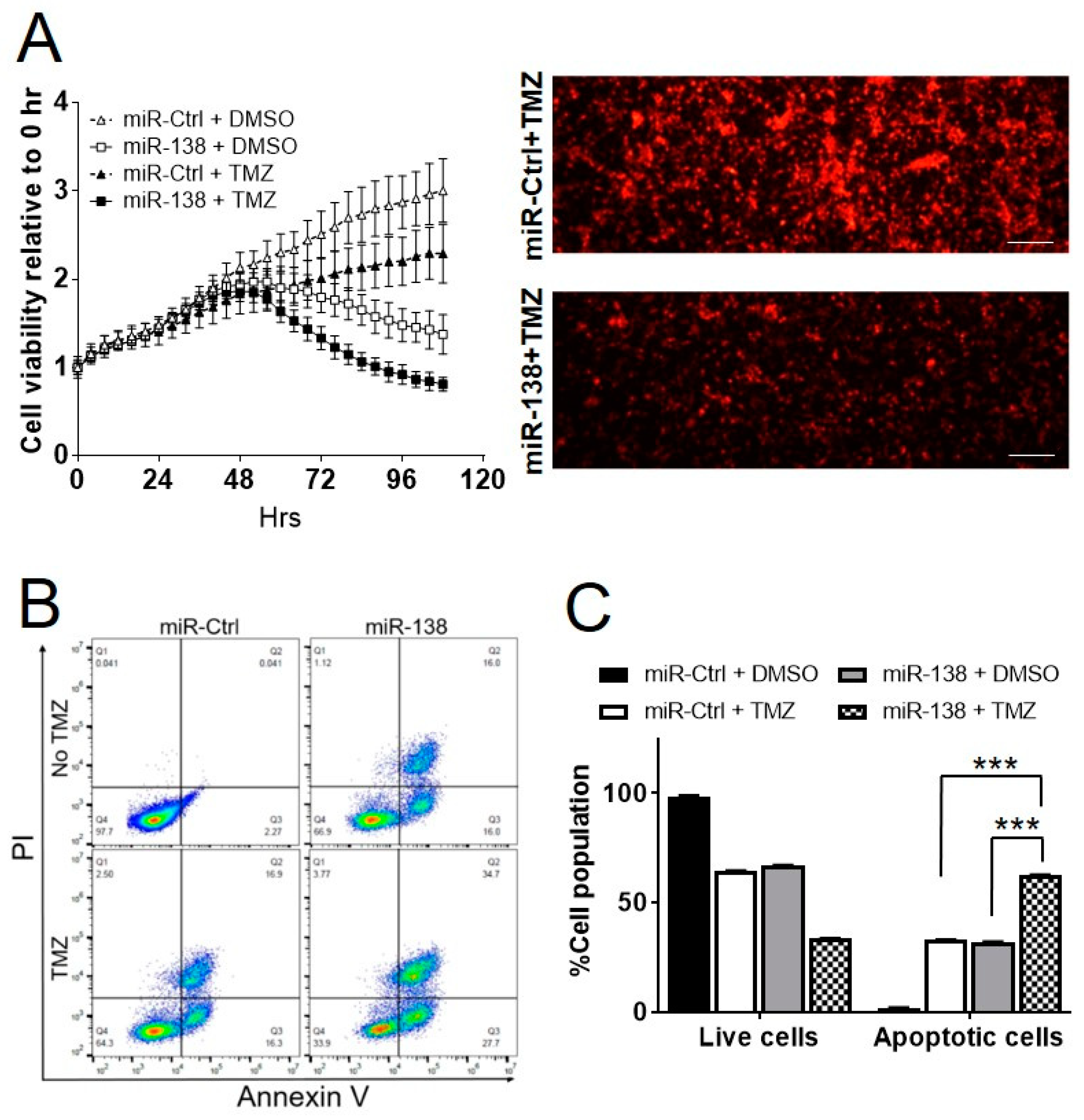
3.1. miR-138 Sensitizes GBM to Chemotherapeutic Drug Temozolomide
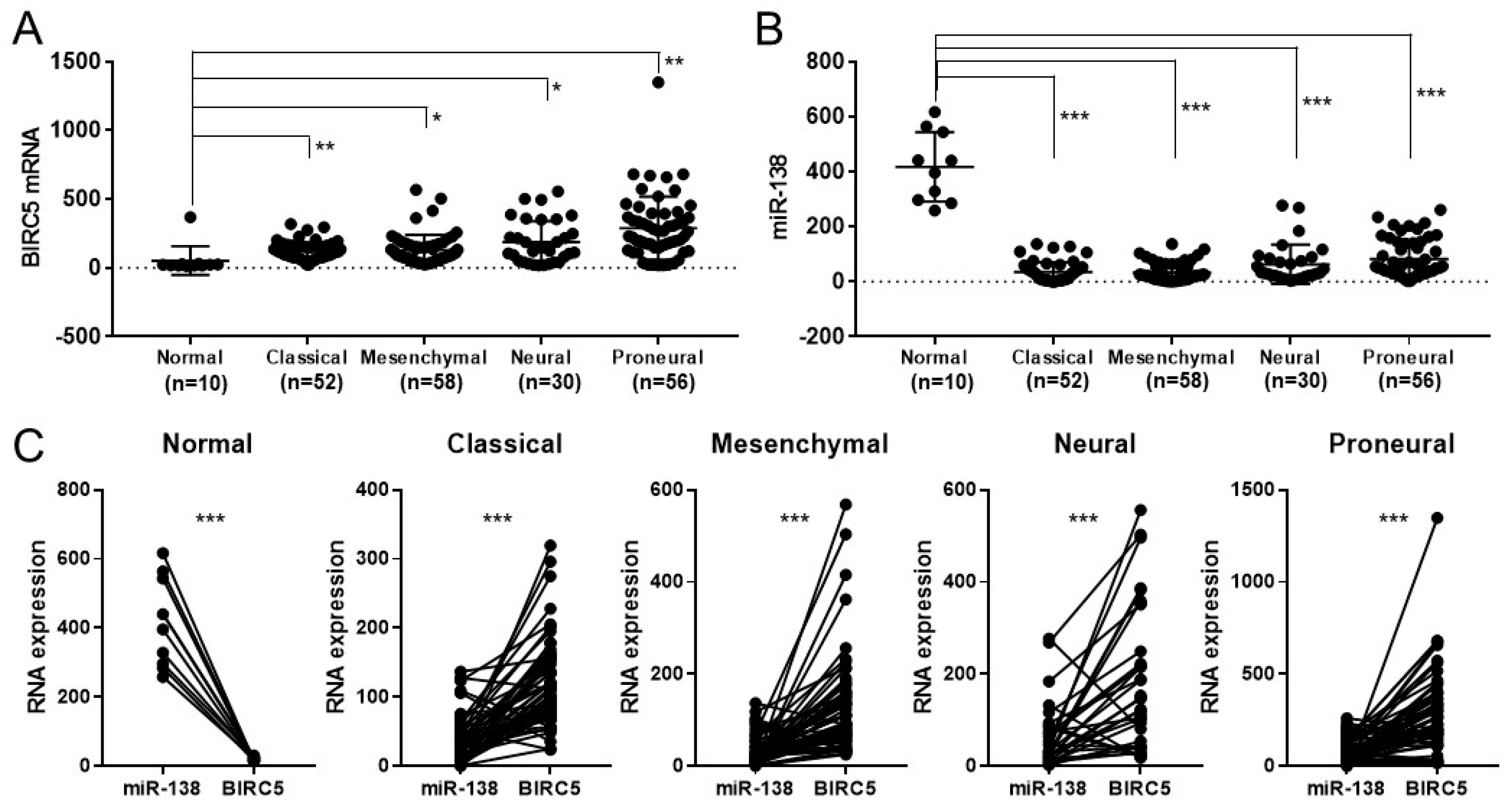
3.2. BICR5 Is Downregulated in Glioblastoma
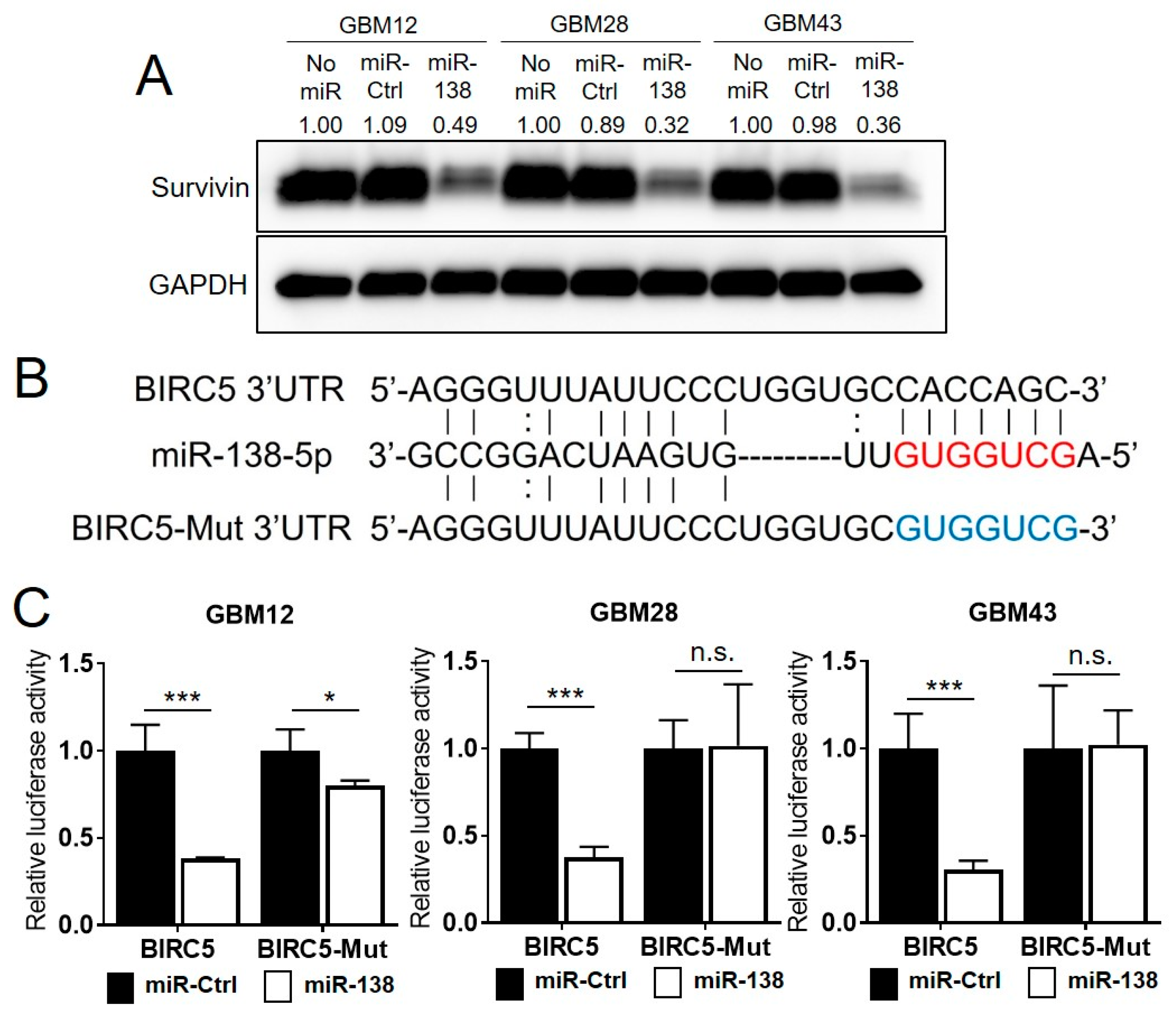
3.3. Survivin Is a Direct Target of miR-138
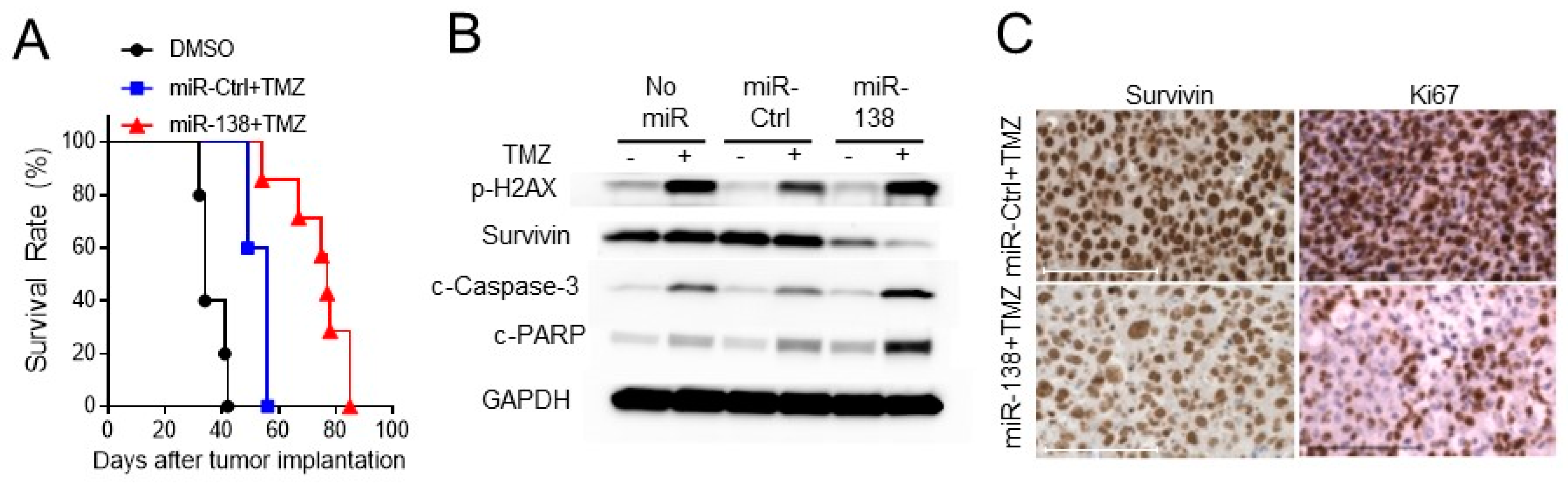
3.4. Ectopic Expression of miR-138 Improves Survival Rates in Intracranial Tumor Bearing Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lesniak, M.S.; Brem, H. Targeted therapy for brain tumours. Nat. Rev. Drug Discov. 2004, 3, 499–508. [Google Scholar] [CrossRef]
- Stupp, R.; Taillibert, S.; Kanner, A.; Read, W.; Steinberg, D.; Lhermitte, B.; Toms, S.; Idbaih, A.; Ahluwalia, M.S.; Fink, K.; et al. Effect of Tumor-Treating Fields Plus Maintenance Temozolomide vs Maintenance Temozolomide Alone on Survival in Patients With Glioblastoma: A Randomized Clinical Trial. JAMA 2017, 318, 2306–2316. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Lahiri, D.; Maji, T.; Biswas, J. Recurrent Glioblastoma: Where we stand. South Asian J. Cancer 2015, 4, 163–173. [Google Scholar] [CrossRef]
- Lee, C.Y. Strategies of temozolomide in future glioblastoma treatment. Onco Targets Ther. 2017, 10, 265–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Anthiya, S.; Griveau, A.; Loussouarn, C.; Baril, P.; Garnett, M.; Issartel, J.P.; Garcion, E. MicroRNA-Based Drugs for Brain Tumors. Trends Cancer 2018, 4, 222–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, M.; Wang, Y.Y.; Yoo, J.Y.; Oh, C.; Otani, Y.; Kang, J.M.; Park, E.S.; Kim, E.; Chung, S.W.; Jeon, Y.J.; et al. MicroRNA-138 Suppresses Glioblastoma Proliferation through Downregulation of CD44. Sci. Rep. 2021, 11, 9219. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Woo, S.J.; Park, J.S.; Kim, H.S.; Park, M.Y.; Park, S.D.; Hong, Y.K.; Kim, T.G. Enhanced antitumour immunity by combined use of temozolomide and TAT-survivin pulsed dendritic cells in a murine glioma. Immunology 2007, 122, 615–622. [Google Scholar] [CrossRef]
- Nitschke, N.J.; Bjoern, J.; Iversen, T.Z.; Andersen, M.H.; Svane, I.M. Indoleamine 2,3-dioxygenase and survivin peptide vaccine combined with temozolomide in metastatic melanoma. Stem Cell Investig. 2017, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Pan, Y.; Ling, G.; Wang, S.; Huang, M.; Jiang, X.; Ke, Y. Escape of U251 glioma cells from temozolomide-induced senescence was modulated by CDK1/survivin signaling. Am. J. Transl. Res. 2017, 9, 2163–2180. [Google Scholar]
- Tang, J.H.; Yang, L.; Chen, J.X.; Li, Q.R.; Zhu, L.R.; Xu, Q.F.; Huang, G.H.; Zhang, Z.X.; Xiang, Y.; Du, L.; et al. Bortezomib inhibits growth and sensitizes glioma to temozolomide (TMZ) via down-regulating the FOXM1-Survivin axis. Cancer Commun. 2019, 39, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, B.L.; Zhou, Y.; Lv, H.; Sun, S.H.; Tang, W.X. MS-275 potentiates the effect of YM-155 in lung adenocarcinoma via survivin downregulation induced by miR-138 and miR-195. Thorac. Cancer 2019, 10, 1355–1368. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wang, L.; Du, X.; Sun, Q.; Wang, Y.; Li, M.; Zang, W.; Liu, K.; Zhao, G. alpha-solanine enhances the chemosensitivity of esophageal cancer cells by inducing microRNA138 expression. Oncol. Rep. 2018, 39, 1163–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Liu, M.; Liang, H.; Guo, S.; Guo, X.; Yuan, M.; Lian, H.; Yan, X.; Zhang, S.; Chen, X.; et al. miR-138-5p contributes to cell proliferation and invasion by targeting Survivin in bladder cancer cells. Mol. Cancer 2016, 15, 82. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.J.; Haque, F.; Shu, D.; Yoo, J.Y.; Li, H.; Yokel, R.A.; Horbinski, C.; Kim, T.H.; Kim, S.H.; Kwon, C.H.; et al. RNA nanoparticle as a vector for targeted siRNA delivery into glioblastoma mouse model. Oncotarget 2015, 6, 14766–14776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.J.; Haque, F.; Vieweger, M.; Yoo, J.Y.; Kaur, B.; Guo, P.; Croce, C.M. Functional assays for specific targeting and delivery of RNA nanoparticles to brain tumor. Methods Mol. Biol. 2015, 1297, 137–152. [Google Scholar] [CrossRef] [Green Version]
- Yeh, M.; Oh, C.S.; Yoo, J.Y.; Kaur, B.; Lee, T.J. Pivotal role of microRNA-138 in human cancers. Am. J. Cancer Res. 2019, 9, 1118–1126. [Google Scholar]
- Sha, H.H.; Wang, D.D.; Chen, D.; Liu, S.W.; Wang, Z.; Yan, D.L.; Dong, S.C.; Feng, J.F. miR-138: A promising therapeutic target for cancer. Tumour Biol. 2017, 39, 1010428317697575. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Chen, D.; Cabay, R.J.; Wang, A.; Crowe, D.L.; Zhou, X. Role of microRNA-138 as a potential tumor suppressor in head and neck squamous cell carcinoma. Int. Rev. Cell Mol. Biol. 2013, 303, 357–385. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, P.K.; Goel, A.; Mittal, R.D. Survivin: A molecular biomarker in cancer. Indian J. Med. Res. 2015, 141, 389–397. [Google Scholar] [CrossRef]
- Varughese, R.K.; Torp, S.H. Survivin and gliomas: A literature review. Oncol. Lett. 2016, 12, 1679–1686. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Nduom, E.K.; Kong, L.Y.; Hashimoto, Y.; Xu, S.; Gabrusiewicz, K.; Ling, X.; Huang, N.; Qiao, W.; Zhou, S.; et al. miR-138 exerts anti-glioma efficacy by targeting immune checkpoints. Neuro-Oncology 2016, 18, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, S.; Marrinan, J.; Frishman, C.; Sampath, P. Impact of temozolomide on immune response during malignant glioma chemotherapy. Clin. Dev. Immunol. 2012, 2012, 831090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, D.; Shu, Y.; Haque, F.; Abdelmawla, S.; Guo, P. Thermodynamically stable RNA three-way junction for constructing multifunctional nanoparticles for delivery of therapeutics. Nat. Nanotechnol. 2011, 6, 658–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pi, F.; Binzel, D.W.; Lee, T.J.; Li, Z.; Sun, M.; Rychahou, P.; Li, H.; Haque, F.; Wang, S.; Croce, C.M.; et al. Nanoparticle orientation to control RNA loading and ligand display on extracellular vesicles for cancer regression. Nat. Nanotechnol. 2018, 13, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.J.; Yoo, J.Y.; Shu, D.; Li, H.; Zhang, J.; Yu, J.G.; Jaime-Ramirez, A.C.; Acunzo, M.; Romano, G.; Cui, R.; et al. RNA Nanoparticle-Based Targeted Therapy for Glioblastoma through Inhibition of Oncogenic miR-21. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 1544–1555. [Google Scholar] [CrossRef] [Green Version]
- Stojcheva, N.; Schechtmann, G.; Sass, S.; Roth, P.; Florea, A.M.; Stefanski, A.; Stuhler, K.; Wolter, M.; Muller, N.S.; Theis, F.J.; et al. MicroRNA-138 promotes acquired alkylator resistance in glioblastoma by targeting the Bcl-2-interacting mediator BIM. Oncotarget 2016, 7, 12937–12950. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, J.-Y.; Yeh, M.; Wang, Y.-Y.; Oh, C.; Zhao, Z.-M.; Kaur, B.; Lee, T.-J. MicroRNA-138 Increases Chemo-Sensitivity of Glioblastoma through Downregulation of Survivin. Biomedicines 2021, 9, 780. https://doi.org/10.3390/biomedicines9070780
Yoo J-Y, Yeh M, Wang Y-Y, Oh C, Zhao Z-M, Kaur B, Lee T-J. MicroRNA-138 Increases Chemo-Sensitivity of Glioblastoma through Downregulation of Survivin. Biomedicines. 2021; 9(7):780. https://doi.org/10.3390/biomedicines9070780
Chicago/Turabian StyleYoo, Ji-Young, Margaret Yeh, Yin-Ying Wang, Christina Oh, Zhong-Ming Zhao, Balveen Kaur, and Tae-Jin Lee. 2021. "MicroRNA-138 Increases Chemo-Sensitivity of Glioblastoma through Downregulation of Survivin" Biomedicines 9, no. 7: 780. https://doi.org/10.3390/biomedicines9070780