
Psychiatric Neural Networks and Precision Therapeutics by Machine Learning


Abstract
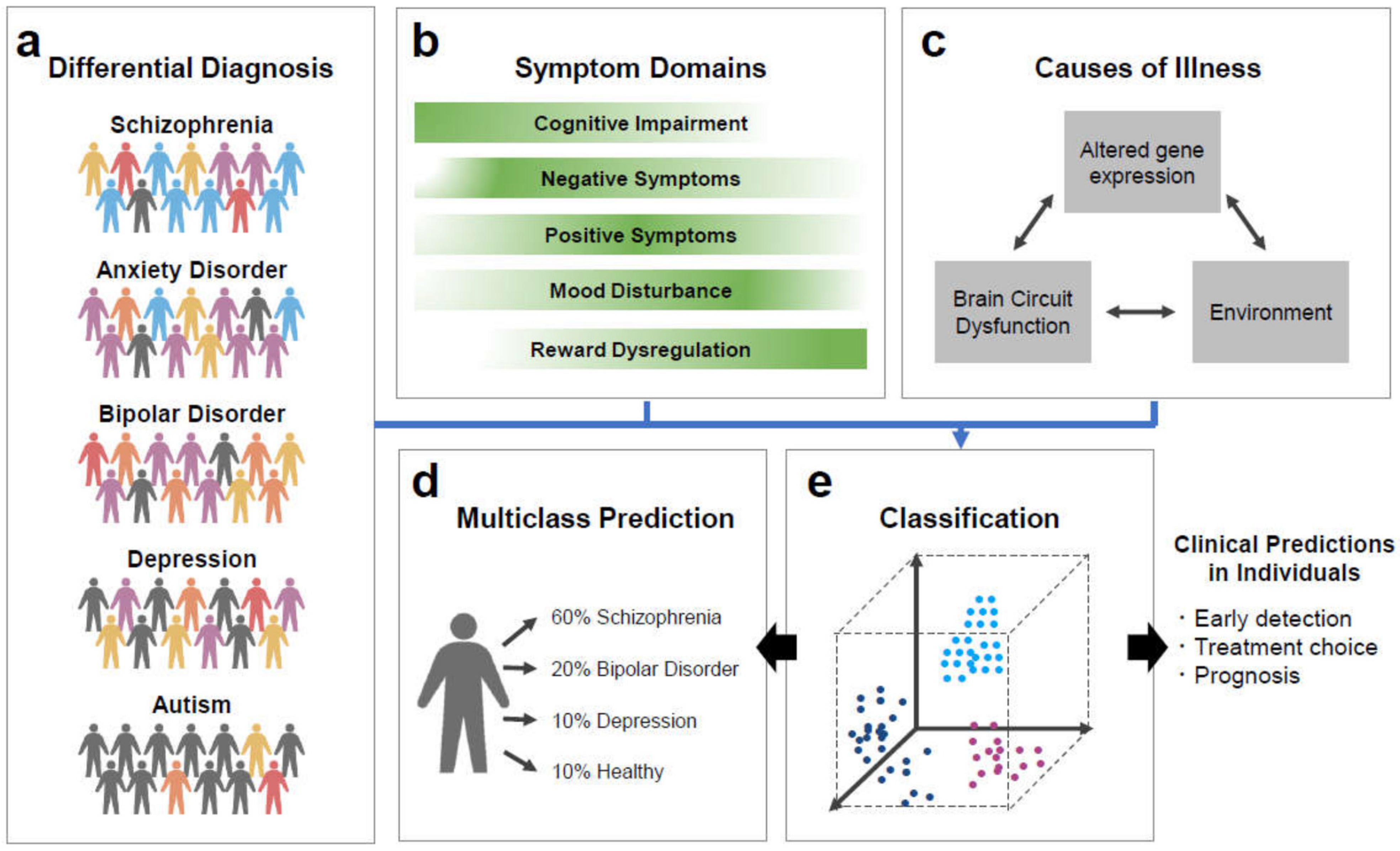
:1. Introduction
2. Decision-Making in Real Life and Psychiatric Disorders
2.1. Schizophrenia
2.2. Bipolar Disorder
2.3. Depression and Anxiety Disorders
2.4. Autism Spectrum Disorder
3. Psychiatric Neural Networks
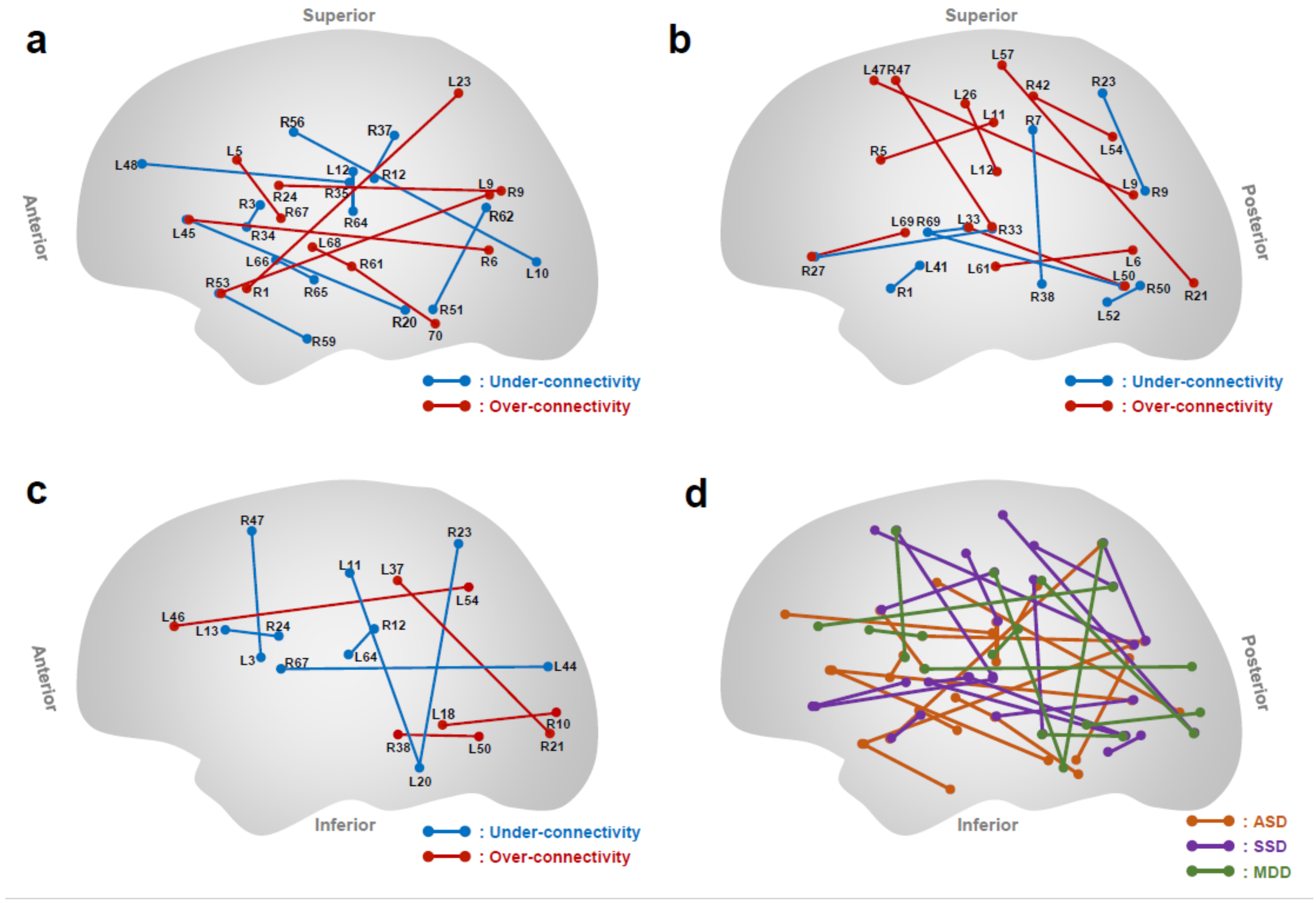
3.1. Functional Connectivity as ASD Classifier
3.2. Functional Connectivity as Schizophrenia Spectrum Disorder Classifier
3.3. Functional Connectivity as MDD Classifier
4. Machine Learning Approach to Predict Therapeutic Outcomes in Psychiatric Disorders
4.1. Prediction of Therapeutic Outcomes in Schizophrenia
4.2. Prediction of Therapeutic Outcomes in Depression
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pontzer, H.; Brown, M.H.; Raichlen, D.A.; Dunsworth, H.; Hare, B.; Walker, K.; Luke, A.; Dugas, L.R.; Durazo-Arvizu, R.; Schoeller, D.; et al. Metabolic acceleration and the evolution of human brain size and life history. Nat. Cell Biol. 2016, 533, 390–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Marin, A.; Ghazanfar, A.A. The Life of Behavior. Neuron 2019, 104, 25–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, B.R.; Gao, W.-J. PV Interneurons: Critical Regulators of E/I Balance for Prefrontal Cortex-Dependent Behavior and Psychiatric Disorders. Front. Neural Circuits 2018, 12, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.M. The attack and defense mechanisms: Perspectives from behavioral economics and game theory. Behav. Brain Sci. 2019, 42, e121. [Google Scholar] [CrossRef]
- Robalino, N.; Robson, A. The economic approach to ‘theory of mind’. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 2224–2233. [Google Scholar] [CrossRef] [Green Version]
- Brosnan, S.F.; Jones, O.D.; Lambeth, S.P.; Mareno, M.C.; Richardson, A.S.; Schapiro, S.J. Endowment Effects in Chimpanzees. Curr. Biol. 2007, 17, 1704–1707. [Google Scholar] [CrossRef] [Green Version]
- Lakshminaryanan, V.; Chen, M.K.; Santos, L.R. Endowment effect in capuchin monkeys. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 3837–3844. [Google Scholar] [CrossRef] [Green Version]
- Santos, L.R.; Hughes, K.D. Economic cognition in humans and animals: The search for core mechanisms. Curr. Opin. Neurobiol. 2009, 19, 63–66. [Google Scholar] [CrossRef]
- Camerer, C.F. Behavioural studies of strategic thinking in games. Trends Cogn. Sci. 2003, 7, 225–231. [Google Scholar] [CrossRef]
- Camerer, C.F. Psychology and economics. Strategizing in the brain. Science 2003, 300, 1673–1675. [Google Scholar] [CrossRef]
- Mackintosh, N.J. Varieties of perceptual learning. Learn. Behav. 2009, 37, 119–125. [Google Scholar] [CrossRef]
- Laurent, P.A. The emergence of saliency and novelty responses from Reinforcement Learning principles. Neural Netw. 2008, 21, 1493–1499. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.M. Neuroscience and Ethics. Exp. Psychol. 2017, 64, 82–92. [Google Scholar] [CrossRef]
- Jones, O.D.; Marois, R.; Farah, M.J.; Greely, H.T. Law and Neuroscience. J. Neurosci. 2013, 33, 17624–17630. [Google Scholar] [CrossRef] [Green Version]
- Arciniegas, D.B.; Anderson, C.A. Toward a Neuroscience of Politics. J. Neuropsychiatry Clin. Neurosci. 2017, 29, 84–85. [Google Scholar] [CrossRef]
- Appleton, K.M.; Bray, J.; Price, S.; Liebchen, G.; Jiang, N.; Mavridis, I.; Saulais, L.; Giboreau, A.; Perez-Cueto, F.J.A.; Coolen, R.; et al. A Mobile Phone App for the Provision of Personalized Food-Based Information in an Eating-Out Situation: Development and Initial Evaluation. JMIR Form. Res. 2019, 3, e12966. [Google Scholar] [CrossRef]
- Frydman, C.; Camerer, C.F. The Psychology and Neuroscience of Financial Decision Making. Trends Cogn. Sci. 2016, 20, 661–675. [Google Scholar] [CrossRef] [Green Version]
- Padoa-Schioppa, C.; Conen, K.E. Orbitofrontal Cortex: A Neural Circuit for Economic Decisions. Neuron 2017, 96, 736–754. [Google Scholar] [CrossRef]
- Rosenberg, A.; Patterson, J.S.; Angelaki, D.E. A computational perspective on autism. Proc. Natl. Acad. Sci. USA 2015, 112, 9158–9165. [Google Scholar] [CrossRef] [Green Version]
- Dayan, P.; Niv, Y.; Seymour, B.; Daw, N.D. The misbehavior of value and the discipline of the will. Neural Netw. 2006, 19, 1153–1160. [Google Scholar] [CrossRef]
- Rangel, A.; Camerer, C.F.; Montague, P.R. A framework for studying the neurobiology of value-based decision making. Nat. Rev. Neurosci. 2008, 9, 545–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Seo, H.; Jung, M.W. Neural Basis of Reinforcement Learning and Decision Making. Annu. Rev. Neurosci. 2012, 35, 287–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Meer, M.; Kurth-Nelson, Z.; Redish, A.D. Information Processing in Decision-Making Systems. Neuroscience 2012, 18, 342–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, M.R.; Dickerson, K.C. Reward-Related Learning via Multiple Memory Systems. Biol. Psychiatry 2012, 72, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Hyman, S.E. Can neuroscience be integrated into the DSM-V? Nat. Rev. Neurosci. 2007, 8, 725–732. [Google Scholar] [CrossRef]
- Sharp, C.; Monterosso, J.; Montague, P.R. Neuroeconomics: A Bridge for Translational Research. Biol. Psychiatry 2012, 72, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Kelly, J.R.; Clarke, G.; Cryan, J.F.; Dinan, T.G. Dimensional thinking in psychiatry in the era of the Research Domain Criteria (RDoC). Ir. J. Psychol. Med. 2017, 35, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, B.N. The RDoC framework: Facilitating transition from ICD/DSM to dimensional approaches that integrate neuroscience and psychopathology. World Psychiatry 2014, 13, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Insel, T.; Cuthbert, B.; Garvey, M.; Heinssen, R.; Pine, D.S.; Quinn, K.; Sanislow, C.; Wang, P. Research Domain Criteria (RDoC): Toward a New Classification Framework for Research on Mental Disorders. Am. J. Psychiatry 2010, 167, 748–751. [Google Scholar] [CrossRef] [Green Version]
- Tai, A.M.; Albuquerque, A.; Carmona, N.E.; Subramanieapillai, M.; Cha, D.S.; Sheko, M.; Lee, Y.; Mansur, R.; McIntyre, R.S. Machine learning and big data: Implications for disease modeling and therapeutic discovery in psychiatry. Artif. Intell. Med. 2019, 99, 101704. [Google Scholar] [CrossRef]
- Galatzer-Levy, I.R.; Ruggles, K.V.; Chen, Z. Data Science in the Research Domain Criteria Era: Relevance of Machine Learning to the Study of Stress Pathology, Recovery, and Resilience. Chronic Stress 2018, 2. [Google Scholar] [CrossRef]
- Koutsouleris, N.; Riecher-Rössler, A.; Meisenzahl, E.M.; Smieskova, R.; Studerus, E.; Kambeitz-Ilankovic, L.; Von Saldern, S.; Cabral, C.; Reiser, M.; Falkai, P.; et al. Detecting the Psychosis Prodrome Across High-Risk Populations Using Neuroanatomical Biomarkers. Schizophr. Bull. 2014, 41, 471–482. [Google Scholar] [CrossRef]
- Chekroud, A.M.; Zotti, R.J.; Shehzad, Z.; Gueorguieva, R.; Johnson, M.K.; Trivedi, M.H.; Cannon, T.D.; Krystal, J.H.; Corlett, P.R. Cross-trial prediction of treatment outcome in depression: A machine learning approach. Lancet Psychiatry 2016, 3, 243–250. [Google Scholar] [CrossRef]
- Koutsouleris, N.; Kahn, R.S.; Chekroud, A.M.; Leucht, S.; Falkai, P.; Wobrock, T.; Derks, E.M.; Fleischhacker, W.W.; Hasan, A. Multisite prediction of 4-week and 52-week treatment outcomes in patients with first-episode psychosis: A machine learning approach. Lancet Psychiatry 2016, 3, 935–946. [Google Scholar] [CrossRef]
- Cáceda, R.; Nemeroff, C.B.; Harvey, P.D. Toward an Understanding of Decision Making in Severe Mental Illness. J. Neuropsychiatry Clin. Neurosci. 2014, 26, 196–213. [Google Scholar] [CrossRef]
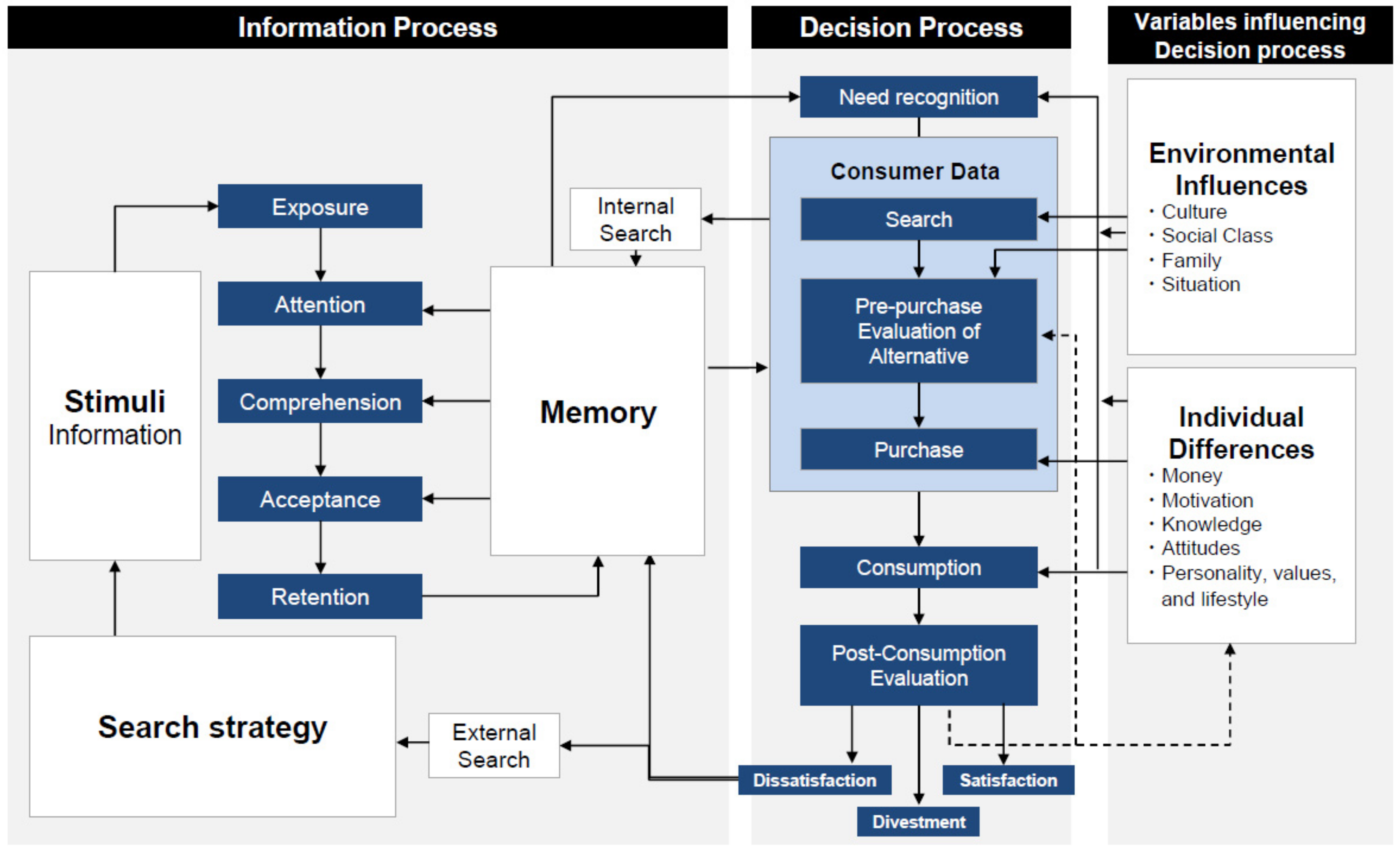
- Blackwell, R.D.; Miniard, P.W.; Engel, J.F. Consumer Behavior, 9th ed.; Harcourt College Publishers: Ft. Worth, TX, USA, 2001; p. 570. [Google Scholar]
- Ammerman, W. The Invisible Brand: Marketing in the Age of Automation, Big Data, and Machine Learning; McGraw-Hill Education: New York, NY, USA, 2019; p. xiv. [Google Scholar]
- Gandal, M.J.; Haney, J.R.; Parikshak, N.N.; Leppa, V.; Ramaswami, G.; Hartl, C.; Schork, A.J.; Appadurai, V.; Buil, A.; Werge, T.M.; et al. Shared molecular neuropathology across major psychiatric disorders parallels polygenic overlap. Science 2018, 359, 693–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishida, K.T.; King-Casas, B.; Montague, P.R. Neuroeconomic Approaches to Mental Disorders. Neuron 2010, 67, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Hasler, G. Can the neuroeconomics revolution revolutionize psychiatry? Neurosci. Biobehav. Rev. 2012, 36, 64–78. [Google Scholar] [CrossRef]
- Montague, P.R.; Dolan, R.J.; Friston, K.J.; Dayan, P. Computational psychiatry. Trends Cogn. Sci. 2012, 16, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Morningstar, J. The Mind Within the Brain: How We Make Decisions and How Those Decisions Go Wrong. Libr. J. 2013, 138, 94. [Google Scholar]
- Andreasen, N.C.; Carpenter, W.T. Diagnosis and Classification of Schizophrenia. Schizophr. Bull. 1993, 19, 199–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, D.A. Cortical circuit dysfunction and cognitive deficits in schizophrenia—implications for preemptive interventions. Eur. J. Neurosci. 2012, 35, 1871–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barch, D.M.; Ceaser, A. Cognition in schizophrenia: Core psychological and neural mechanisms. Trends Cogn. Sci. 2012, 16, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberger, D.R.; Berman, K.F.; Zec, R.F. Physiologic dysfunction of dorsolateral prefrontal cortex in schizophrenia. I. Regional cerebral blood flow evidence. Arch. Gen. Psychiatry 1986, 43, 114–124. [Google Scholar] [CrossRef]
- Heerey, E.A.; Bell-Warren, K.R.; Gold, J.M. Decision-Making Impairments in the Context of Intact Reward Sensitivity in Schizophrenia. Biol. Psychiatry 2008, 64, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Waltz, J.A.; Frank, M.J.; Robinson, B.M.; Gold, J.M. Selective Reinforcement Learning Deficits in Schizophrenia Support Predictions from Computational Models of Striatal-Cortical Dysfunction. Biol. Psychiatry 2007, 62, 756–764. [Google Scholar] [CrossRef] [Green Version]
- Strauss, G.P.; Frank, M.J.; Waltz, J.A.; Kasanova, Z.; Herbener, E.S.; Gold, J.M. Deficits in Positive Reinforcement Learning and Uncertainty-Driven Exploration Are Associated with Distinct Aspects of Negative Symptoms in Schizophrenia. Biol. Psychiatry 2011, 69, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Corlett, P.R.; Murray, G.K.; Honey, G.D.; Aitken, M.R.F.; Shanks, D.R.; Robbins, T.; Bullmore, E.; Dickinson, A.; Fletcher, P.C. Disrupted prediction-error signal in psychosis: Evidence for an associative account of delusions. Brain 2007, 130, 2387–2400. [Google Scholar] [CrossRef]
- Gradin, V.B.; Kumar, P.; Waiter, G.; Ahearn, T.; Stickle, C.; Milders, M.; Reid, I.; Hall, J.; Steele, J.D. Expected value and prediction error abnormalities in depression and schizophrenia. Brain 2011, 134, 1751–1764. [Google Scholar] [CrossRef] [Green Version]
- Volkow, N.D. Substance Use Disorders in Schizophrenia—Clinical Implications of Comorbidity. Schizophr. Bull. 2009, 35, 469–472. [Google Scholar] [CrossRef] [Green Version]
- Goff, D.C.; Henderson, D.C.; Amico, E. Cigarette smoking in schizophrenia: Relationship to psychopathology and med-ication side effects. Am. J. Psychiatry 1992, 149, 1189–1194. [Google Scholar] [PubMed]
- De Leon, J.; Dadvand, M.; Canuso, C.; White, A.O.; Stanilla, J.K.; Simpson, G.M. Schizophrenia and smoking: An epi-demiological survey in a state hospital. Am. J. Psychiatry 1995, 152, 453–455. [Google Scholar] [PubMed]
- Krystal, J.H.; D’Souza, D.C.; Gallinat, J.; Driesen, N.; Abi-Dargham, A.; Petrakis, I.; Heinz, A.; Pearlson, G. The vulner-ability to alcohol and substance abuse in individuals diagnosed with schizophrenia. Neurotox. Res. 2006, 10, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Harvey, P.D.; Wingo, A.P.; Burdick, K.E.; Baldessarini, R.J. Cognition and disability in bipolar disorder: Lessons from schizophrenia research. Bipolar Disord. 2010, 12, 364–375. [Google Scholar] [CrossRef]
- Martino, D.J.; Strejilevich, S.A.; Scápola, M.; Igoa, A.; Marengo, E.; Ais, E.D.; Perinot, L. Heterogeneity in cognitive functioning among patients with bipolar disorder. J. Affect. Disord. 2008, 109, 149–156. [Google Scholar] [CrossRef]
- Gruber, J. A Review and Synthesis of Positive Emotion and Reward Disturbance in Bipolar Disorder. Clin. Psychol. Psychother. 2011, 18, 356–365. [Google Scholar] [CrossRef]
- Samame, C.; Martino, D.J.; Strejilevich, S.A. Social cognition in euthymic bipolar disorder: Systematic review and me-ta-analytic approach. Acta Psychiatr. Scand. 2012, 125, 266–280. [Google Scholar] [CrossRef]
- Robinson, L.J.; Thompson, J.M.; Gallagher, P.; Goswami, U.; Young, A.H.; Ferrier, I.N.; Moore, P.B. A meta-analysis of cognitive deficits in euthymic patients with bipolar disorder. J. Affect. Disord. 2006, 93, 105–115. [Google Scholar] [CrossRef]
- Thompson, J.M.; Gallagher, P.; Hughes, J.H.; Watson, S.; Gray, J.M.; Ferrier, I.N.; Young, A.H. Neurocognitive im-pairment in euthymic patients with bipolar affective disorder. Br. J. Psychiatry 2005, 186, 32–40. [Google Scholar] [CrossRef]
- Torres, I.J.; Boudreau, V.G.; Yatham, L.N. Neuropsychological functioning in euthymic bipolar disorder: A meta-analysis. Acta Psychiatr. Scand. 2007, 116, 17–26. [Google Scholar] [CrossRef]
- Rau, G.; Blair, K.S.; Berghorst, L.; Knopf, L.; Skup, M.; Luckenbaugh, D.A.; Pine, D.S.; Blair, R.J.; Leibenluft, E. Processing of Differentially Valued Rewards and Punishments in Youths with Bipolar Disorder or Severe Mood Dysregulation. J. Child. Adolesc. Psychopharmacol. 2008, 18, 185–196. [Google Scholar] [CrossRef]
- Ernst, M.; Dickstein, D.P.; Munson, S.; Eshel, N.; Pradella, A.; Jazbec, S.; Pine, D.S.; Leibenluft, E. Reward-related pro-cesses in pediatric bipolar disorder: A pilot study. J. Affect. Disord. 2004, 82 (Suppl. 1), S89–S101. [Google Scholar] [CrossRef]
- Kovacs, M.; Devlin, B. Internalizing disorders in childhood. J. Child Psychol. Psychiatry 1998, 39, 47–63. [Google Scholar] [CrossRef]
- Krueger, R.F. Internalization and Externalization and the Structure of Common Mental Disorders. Psycextra Dataset 2004, 56, 921–926. [Google Scholar] [CrossRef]
- Mineka, S.; Watson, D.; Clark, L.A. Comorbidity of anxiety and unipolar mood disorders. Annu. Rev. Psychol. 1998, 49, 377–412. [Google Scholar] [CrossRef] [Green Version]
- Paulus, M.P.; Yu, A.J. Emotion and decision-making: Affect-driven belief systems in anxiety and depression. Trends Cogn. Sci. 2012, 16, 476–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolen-Hoeksema, S. The role of rumination in depressive disorders and mixed anxiety/depressive symptoms. J. Abnorm. Psychol. 2000, 109, 504–511. [Google Scholar] [CrossRef]
- Eshel, N.; Roiser, J.P. Reward and Punishment Processing in Depression. Biol. Psychiatry 2010, 68, 118–124. [Google Scholar] [CrossRef]
- Craske, M.G.; Rauch, S.L.; Ursano, R.; Prenoveau, J.; Pine, D.S.; Zinbarg, R.E. What is an anxiety disorder? Depress. Anxiety 2009, 26, 1066–1085. [Google Scholar] [CrossRef]
- Hartley, C.A.; Phelps, E.A. Anxiety and Decision-Making. Biol. Psychiatry 2012, 72, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Sheline, Y.I.; Barch, D.M.; Price, J.L.; Rundle, M.M.; Vaishnavi, S.N.; Snyder, A.Z.; Mintun, M.A.; Wang, S.; Coalson, R.S.; Raichle, M.E. The default mode network and self-referential processes in depression. Proc. Natl. Acad. Sci. USA 2009, 106, 1942–1947. [Google Scholar] [CrossRef] [Green Version]
- Mayberg, H.S.; Lozano, A.M.; Voon, V.; McNeely, H.E.; Seminowicz, D.; Hamani, C.; Schwalb, J.M.; Kennedy, S.H. Deep brain stimulation for treatment-resistant depression. Neuron 2005, 45, 651–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayan, P.; Huys, Q.J.M. Serotonin in Affective Control. Annu. Rev. Neurosci. 2009, 32, 95–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doya, K. Metalearning and neuromodulation. Neural Netw. 2002, 15, 495–506. [Google Scholar] [CrossRef]
- Daw, N.D.; Kakade, S.; Dayan, P. Opponent interactions between serotonin and dopamine. Neural Netw. 2002, 15, 603–616. [Google Scholar] [CrossRef]
- Dayan, P.; Huys, Q.J. Serotonin, inhibition, and negative mood. PLoS Comput. Biol. 2008, 4, e4. [Google Scholar] [CrossRef] [Green Version]
- Huys, Q.J.M.; Eshel, N.; O’Nions, E.; Sheridan, L.; Dayan, P.; Roiser, J.P. Bonsai Trees in Your Head: How the Pavlovian System Sculpts Goal-Directed Choices by Pruning Decision Trees. PLoS Comput. Biol. 2012, 8, e1002410. [Google Scholar] [CrossRef] [Green Version]
- Geschwind, D.H.; Levitt, P. Autism spectrum disorders: Developmental disconnection syndromes. Curr. Opin. Neurobiol. 2007, 17, 103–111. [Google Scholar] [CrossRef]
- Baron-Cohen, S.; Leslie, A.M.; Frith, U. Does the autistic child have a “theory of mind”? Cognition 1985, 21, 37–46. [Google Scholar] [CrossRef]
- Frith, U. Mind Blindness and the Brain in Autism. Neuron 2001, 32, 969–979. [Google Scholar] [CrossRef] [Green Version]
- Wiggins, L.D.; Rice, C.E.; Barger, B.; Soke, G.N.; Lee, L.-C.; Moody, E.; Edmondson-Pretzel, R.; Levy, S.E. DSM-5 criteria for autism spectrum disorder maximizes diagnostic sensitivity and specificity in preschool children. Soc. Psychiatry Psychiatr. Epidemiol. 2019, 54, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Hadjikhani, N.; Joseph, R.M.; Snyder, J.; Tager-Flusberg, H. Abnormal activation of the social brain during face perception in autism. Hum. Brain Mapp. 2006, 28, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Nebel, M.B.; Joel, S.E.; Muschelli, J.; Barber, A.D.; Caffo, B.S.; Pekar, J.J.; Mostofsky, S.H. Disruption of functional organization within the primary motor cortex in children with autism. Hum. Brain Mapp. 2012, 35, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staples, K.L.; Reid, G. Fundamental Movement Skills and Autism Spectrum Disorders. J. Autism Dev. Disord. 2010, 40, 209–217. [Google Scholar] [CrossRef]
- Breveglieri, R.; Galletti, C.; Gamberini, M.; Passarelli, L.; Fattori, P. Somatosensory Cells in Area PEc of Macaque Posterior Parietal Cortex. J. Neurosci. 2006, 26, 3679–3684. [Google Scholar] [CrossRef] [Green Version]
- Courchesne, E. Brainstem, cerebellar and limbic neuroanatomical abnormalities in autism. Curr. Opin. Neurobiol. 1997, 7, 269–278. [Google Scholar] [CrossRef]
- Frith, U. Autism: Explaining the Enigma, 2nd ed.; Blackwell Pub.: New York, NY, USA, 2003. [Google Scholar]
- Müller, R.-A.; Kleinhans, N.; Kemmotsu, N.; Pierce, K.; Courchesne, E. Abnormal Variability and Distribution of Functional Maps in Autism: An fMRI Study of Visuomotor Learning. Am. J. Psychiatry 2003, 160, 1847–1862. [Google Scholar] [CrossRef] [Green Version]
- Hadjikhani, N.; Joseph, R.M.; Snyder, J.; Tager-Flusberg, H. Anatomical Differences in the Mirror Neuron System and Social Cognition Network in Autism. Cereb. Cortex 2005, 16, 1276–1282. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.; Whiten, A.; Suddendorf, T.; Perrett, D. Imitation, mirror neurons and autism. Neurosci. Biobehav. Rev. 2001, 25, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Baron-Cohen, S.; Ring, H.; Bullmore, E.; Wheelwright, S.; Ashwin, C.; Williams, S. The amygdala theory of autism. Neurosci. Biobehav. Rev. 2000, 24, 355–364. [Google Scholar] [CrossRef]
- Brothers, L. Brain mechanisms of social cognition. J. Psychopharmacol. 1996, 10, 2–8. [Google Scholar] [CrossRef]
- Ernst, M.; Bolla, K.; Mouratidis, M.; Contoreggi, C.; Matochik, J.A.; Kurian, V.; Cadet, J.-L.; Kimes, A.S.; London, E.D. Decision-making in a Risk-taking Task A PET Study. Neuropsychopharmacol. 2002, 26, 682–691. [Google Scholar] [CrossRef]
- Bar-On, R.; Tranel, D.; Denburg, N.L.; Bechara, A. Exploring the neurological substrate of emotional and social intelli-gence. Brain 2003, 126, 1790–1800. [Google Scholar] [CrossRef]
- Bechara, A. The role of emotion in decision-making: Evidence from neurological patients with orbitofrontal damage. Brain Cogn. 2004, 55, 30–40. [Google Scholar] [CrossRef]
- Cao, H.; Plichta, M.M.; Schafer, A.; Haddad, L.; Grimm, O.; Schneider, M.; Esslinger, C.; Kirsch, P.; Meyer-Lindenberg, A.; Tost, H. Test-retest reliability of fMRI-based graph theoretical properties during working memory, emotion processing, and resting state. NeuroImage 2014, 84, 888–900. [Google Scholar] [CrossRef]
- Drysdale, A.T.; Grosenick, L.; Downar, J.; Dunlop, K.; Mansouri, F.; Meng, Y.; Fetcho, R.N.; Zebley, B.; Oathes, D.J.; Etkin, A.; et al. Resting-state connectivity biomarkers define neurophysio-logical subtypes of depression. Nat. Med. 2017, 23, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Sundermann, B.; Feder, S.; Wersching, H.; Teuber, A.; Schwindt, W.; Kugel, H.; Heindel, W.; Arolt, V.; Berger, K.; Pfleiderer, B. Diagnostic classification of unipolar depression based on resting-state functional connectivity MRI: Effects of generali-zation to a diverse sample. J. Neural. Transm. 2017, 124, 589–605. [Google Scholar] [CrossRef]
- Kupfer, D.J.; Frank, E.; Phillips, M.L. Major depressive disorder: New clinical, neurobiological, and treatment perspectives. Lancet 2012, 379, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.T.; Holmes, A.J.; Masters, G.A.; Yeo, B.T.T.; Krienen, F.; Buckner, R.L.; Öngür, D. Disruption of Cortical Association Networks in Schizophrenia and Psychotic Bipolar Disorder. JAMA Psychiatry 2014, 71, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Harrison, B.J.; Soriano-Mas, C.; Pujol, J.; Ortiz, H.; Lopez-Sola, M.; Hernandez-Ribas, R.; Deus, J.; Alonso, P.; Yucel, M.; Pantelis, C.; et al. Altered corticostriatal functional connectivity in obsessive-compulsive disorder. Arch. Gen. Psychiatry 2009, 66, 1189–1200. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, R.H.; Andrews-Hanna, J.R.; Wager, T.D.; Pizzagalli, D.A. Large-Scale Network Dysfunction in Major Depressive Disorder: A Meta-analysis of Resting-State Functional Connectivity. JAMA Psychiatry 2015, 72, 603–611. [Google Scholar] [CrossRef]
- Yahata, N.; Morimoto, J.; Hashimoto, R.; Lisi, G.; Shibata, K.; Kawakubo, Y.; Kuwabara, H.; Kuroda, M.; Yamada, T.; Megumi, F.; et al. A small number of abnormal brain connections predicts adult autism spectrum disorder. Nat. Commun. 2016, 7, 11254. [Google Scholar] [CrossRef] [Green Version]
- Shaposhnyk, V.; Villa, A.E. Reciprocal projections in hierarchically organized evolvable neural circuits affect EEG-like signals. Brain Res. 2012, 1434, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Baldassarre, A.; Lewis, C.M.; Committeri, G.; Snyder, A.Z.; Romani, G.L.; Corbetta, M. Individual variability in functional connectivity predicts performance of a perceptual task. Proc. Natl. Acad. Sci. USA 2012, 109, 3516–3521. [Google Scholar] [CrossRef] [Green Version]
- Dosenbach, N.U.F.; Nardos, B.; Cohen, A.L.; Fair, D.A.; Power, J.D.; Church, J.A.; Nelson, S.M.; Wig, G.S.; Vogel, A.C.; Lessov-Schlaggar, C.N.; et al. Prediction of Individual Brain Maturity Using fMRI. Science 2010, 329, 1358–1361. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.M.; Baldassarre, A.; Committeri, G.; Romani, G.L.; Corbetta, M. Learning sculpts the spontaneous activity of the resting human brain. Proc. Natl. Acad. Sci. USA 2009, 106, 17558–17563. [Google Scholar] [CrossRef] [Green Version]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable Intrinsic Connectivity Networks for Salience Processing and Executive Control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef]
- Finn, E.S.; Shen, X.; Scheinost, D.; Rosenberg, M.D.; Huang, J.; Chun, M.M.; Papademetris, X.; Constable, R.T. Functional connectome fingerprinting: Identifying individuals using patterns of brain connectivity. Nat. Neurosci. 2015, 18, 1664–1671. [Google Scholar] [CrossRef]
- Rosenberg, M.D.; Finn, E.S.; Scheinost, D.; Papademetris, X.; Shen, X.; Constable, R.T.; Chun, M.M. A neuromarker of sustained attention from whole-brain functional connectivity. Nat. Neurosci. 2016, 19, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.M.; Nichols, T.E.; Vidaurre, D.; Winkler, A.M.; Behrens, T.E.J.; Glasser, M.F.; Ugurbil, K.; Barch, D.M.; Van Essen, D.C.; Miller, K.L. A positive-negative mode of population covariation links brain connectivity, demographics and behavior. Nat. Neurosci. 2015, 18, 1565–1567. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, M.; Kawato, M.; Imamizu, H. Predicting learning plateau of working memory from whole-brain intrinsic network connectivity patterns. Sci. Rep. 2015, 5, 7622. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Yoshihara, Y.; Hashimoto, R.; Yahata, N.; Ichikawa, N.; Sakai, Y.; Yamada, T.; Matsukawa, N.; Okada, G.; Tanaka, S.C.; et al. A prediction model of working memory across health and psychiatric disease using whole-brain functional connectivity. eLife 2018, 7, e38844. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, Y.; Lisi, G.; Yahata, N.; Fujino, J.; Matsumoto, Y.; Miyata, J.; Sugihara, G.-I.; Urayama, S.-I.; Kubota, M.; Yamashita, M.; et al. Overlapping but Asymmetrical Relationships Between Schizophrenia and Autism Revealed by Brain Connectivity. Schizophr. Bull. 2020, 46, 1210–1218. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, N.; Lisi, G.; Yahata, N.; Okada, G.; Takamura, M.; Hashimoto, R.I.; Yamada, T.; Yamada, M.; Suhara, T.; Moriguchi, S.; et al. Primary functional brain connections associated with melancholic major depressive disorder and modulation by antidepressants. Sci. Rep. 2020, 10, 3542. [Google Scholar] [CrossRef] [Green Version]
- Ecker, C.; Marquand, A.; Mourao-Miranda, J.; Johnston, P.; Daly, E.M.; Brammer, M.J.; Maltezos, S.; Murphy, C.M.; Robertson, D.; Williams, S.C.; et al. Describing the brain in autism in five dimensions--magnetic resonance im-aging-assisted diagnosis of autism spectrum disorder using a multiparameter classification approach. J. Neurosci. 2010, 30, 10612–10623. [Google Scholar] [CrossRef] [Green Version]
- Ecker, C.; Rocha-Rego, V.; Johnston, P.; Mourao-Miranda, J.; Marquand, A.; Daly, E.M.; Brammer, M.J.; Murphy, C.; Murphy, D.G. Investigating the predictive value of whole-brain structural MR scans in autism: A pattern classification approach. NeuroImage 2010, 49, 44–56. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Menon, V.; Young, C.B.; Ryali, S.; Chen, T.; Khouzam, A.; Minshew, N.J.; Hardan, A.Y. Multivariate Searchlight Classification of Structural Magnetic Resonance Imaging in Children and Adolescents with Autism. Biol. Psychiatry 2011, 70, 833–841. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.S.; Nielsen, J.A.; Froehlich, A.L.; DuBray, M.B.; Druzgal, T.J.; Cariello, A.N.; Cooperrider, J.R.; Zielinski, B.A.; Ravichandran, C.; Fletcher, P.T.; et al. Functional connectivity magnetic resonance imaging classification of autism. Brain 2011, 134, 3742–3754. [Google Scholar] [CrossRef]
- Ingalhalikar, M.; Parker, D.; Bloy, L.; Roberts, T.P.; Verma, R. Diffusion based abnormality markers of pathology: Toward learned diagnostic prediction of ASD. NeuroImage 2011, 57, 918–927. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Chen, C.; Fushing, H. Extracting Multiscale Pattern Information of fMRI Based Functional Brain Connectivity with Application on Classification of Autism Spectrum Disorders. PLoS ONE 2012, 7, e45502. [Google Scholar] [CrossRef]
- Edeshpande, G.; Libero, L.E.; Sreenivasan, K.R.; Deshpande, H.D.; Kana, R.K. Identification of neural connectivity signatures of autism using machine learning. Front. Hum. Neurosci. 2013, 7, 670. [Google Scholar] [CrossRef] [Green Version]
- Tomasi, D.; Volkow, N.D. Gender differences in brain functional connectivity density. Hum. Brain Mapp. 2011, 33, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Klaassens, B.L.; Van Gorsel, H.C.; Khalili-Mahani, N.; Van Der Grond, J.; Wyman, B.T.; Whitcher, B.; Rombouts, S.A.; Van Gerven, J.M. Single-dose serotonergic stimulation shows widespread effects on functional brain connectivity. NeuroImage 2015, 122, 440–450. [Google Scholar] [CrossRef]
- Friedman, L.; Glover, G.H.; Fbirn, C. Reducing interscanner variability of activation in a multicenter fMRI study: Con-trolling for signal-to-fluctuation-noise-ratio (SFNR) differences. NeuroImage 2006, 33, 471–481. [Google Scholar] [CrossRef]
- Thulborn, K.R.; Chang, S.Y.; Shen, G.X.; Voyvodic, J.T. High-resolution echo-planar fMRI of human visual cortex at 3.0 tesla. Nmr Biomed. 1997, 10, 183–190. [Google Scholar] [CrossRef]
- Van Dijk, K.R.A.; Hedden, T.; Venkataraman, A.; Evans, K.C.; Lazar, S.W.; Buckner, R.L. Intrinsic Functional Connectivity as a Tool for Human Connectomics: Theory, Properties, and Optimization. J. Neurophysiol. 2010, 103, 297–321. [Google Scholar] [CrossRef]
- Whelan, R.; Garavan, H. When Optimism Hurts: Inflated Predictions in Psychiatric Neuroimaging. Biol. Psychiatry 2014, 75, 746–748. [Google Scholar] [CrossRef]
- The IMAGEN Consortium; Whelan, R.J.; Watts, R.; Orr, C.A.; Althoff, R.R.; Artiges, E.; Banaschewski, T.; Barker, G.J.; Bokde, A.L.W.; Büchel, C.; et al. Neuropsychosocial profiles of current and future adolescent alcohol misusers. Nat. Cell Biol. 2014, 512, 185–189. [Google Scholar] [CrossRef]
- Tu, P.-C.; Hsieh, J.-C.; Li, C.-T.; Bai, Y.-M.; Su, T.-P. Cortico-striatal disconnection within the cingulo-opercular network in schizophrenia revealed by intrinsic functional connectivity analysis: A resting fMRI study. NeuroImage 2012, 59, 238–247. [Google Scholar] [CrossRef]
- Cross-Disorder Group of the Psychiatric Genomics Consortium. Identification of risk loci with shared effects on five major psychiatric disorders: A genome-wide analysis. Lancet 2013, 381, 1371–1379. [Google Scholar]
- Geschwind, D.H.; Flint, J. Genetics and genomics of psychiatric disease. Science 2015, 349, 1489–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, B.H.; Lord, C. Is schizophrenia on the autism spectrum? Brain Res. 2011, 1380, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Pinkham, A.E.; Hopfinger, J.B.; Pelphrey, K.A.; Piven, J.; Penn, D.L. Neural bases for impaired social cognition in schizophrenia and autism spectrum disorders. Schizophr. Res. 2008, 99, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Witten, D.M.; Tibshirani, R.; Hastie, T. A penalized matrix decomposition, with applications to sparse principal compo-nents and canonical correlation analysis. Biostatistics 2009, 10, 515–534. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, O.; Sato, M.-A.; Yoshioka, T.; Tong, F.; Kamitani, Y. Sparse estimation automatically selects voxels relevant for the decoding of fMRI activity patterns. NeuroImage 2008, 42, 1414–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaugler, T.; Klei, L.; Sanders, S.J.; Bodea, C.A.; Goldberg, A.P.; Lee, A.B.; Mahajan, M.C.; Manaa, D.; Pawitan, Y.; Reichert, J.G.; et al. Most genetic risk for autism resides with common variation. Nat. Genet. 2014, 46, 881–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, P.F.; Magnusson, C.; Reichenberg, A.; Boman, M.; Dalman, C.; Davidson, M.; Fruchter, E.; Hultman, C.M.; Lundberg, M.; Långström, N.; et al. Family History of Schizophrenia and Bipolar Disorder as Risk Factors for Autism. Arch. Gen. Psychiatry 2012, 69, 1099–1103. [Google Scholar] [CrossRef] [Green Version]
- Arbabshirani, M.R.; Plis, S.; Sui, J.; Calhoun, V.D. Single subject prediction of brain disorders in neuroimaging: Promises and pitfalls. NeuroImage 2017, 145, 137–165. [Google Scholar] [CrossRef] [Green Version]
- Akil, H.; Gordon, J.; Hen, R.; Javitch, J.; Mayberg, H.; McEwen, B.; Meaney, M.J.; Nestler, E.J. Treatment resistant de-pression: A multi-scale, systems biology approach. Neurosci. Biobehav. Rev. 2018, 84, 272–288. [Google Scholar] [CrossRef]
- Kendler, K.S. The Diagnostic Validity of Melancholic Major Depression in a Population-Based Sample of Female Twins. Arch. Gen. Psychiatry 1997, 54, 299–304. [Google Scholar] [CrossRef]
- Sun, N.; Li, Y.; Cai, Y.; Chen, J.; Shen, Y.; Sun, J.; Zhang, Z.; Zhang, J.; Wang, L.; Guo, L.; et al. A comparison of melancholic and nonmelan-cholic recurrent major depression in Han Chinese women. Depress. Anxiety 2012, 29, 4–9. [Google Scholar] [CrossRef] [Green Version]
- Consortium, C. Sparse whole-genome sequencing identifies two loci for major depressive disorder. Nature 2015, 523, 588–591. [Google Scholar] [CrossRef]
- Hyett, M.P.; Breakspear, M.J.; Friston, K.J.; Guo, C.C.; Parker, G.B. Disrupted Effective Connectivity of Cortical Systems Supporting Attention and Interoception in Melancholia. JAMA Psychiatry 2015, 72, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Greening, S.G.; Finger, E.C.; Mitchell, D.G. Parsing decision making processes in prefrontal cortex: Response inhibition, overcoming learned avoidance, and reversal learning. NeuroImage 2011, 54, 1432–1441. [Google Scholar] [CrossRef]
- Dombrovski, A.Y.; Szanto, K.; Clark, L.; Aizenstein, H.J.; Chase, H.W.; Reynolds, C.F., 3rd; Siegle, G.J. Corticostria-tothalamic reward prediction error signals and executive control in late-life depression. Psychol. Med. 2015, 45, 1413–1424. [Google Scholar] [CrossRef] [Green Version]
- Mascaro, J.S.; Rilling, J.K.; Negi, L.T.; Raison, C.L. Compassion meditation enhances empathic accuracy and related neural activity. Soc. Cogn. Affect. Neurosci. 2013, 8, 48–55. [Google Scholar] [CrossRef]
- LeWinn, K.Z.; Strigo, I.A.; Connolly, C.G.; Ho, T.C.; Tymofiyeva, O.; Sacchet, M.D.; Weng, H.Y.; Blom, E.H.; Simmons, A.N.; Yang, T.T. An exploratory examination of reappraisal success in depressed adolescents: Preliminary evidence of functional differences in cognitive control brain regions. J. Affect. Disord. 2018, 240, 155–164. [Google Scholar] [CrossRef]
- Ferrari, C.; Lega, C.; Vernice, M.; Tamietto, M.; Mende-Siedlecki, P.; Vecchi, T.; Todorov, A.; Cattaneo, Z. The Dorsomedial Prefrontal Cortex Plays a Causal Role in Integrating Social Impressions from Faces and Verbal Descriptions. Cereb. Cortex 2014, 26, 156–165. [Google Scholar] [CrossRef]
- Mattavelli, G.; Cattaneo, Z.; Papagno, C. Transcranial magnetic stimulation of medial prefrontal cortex modulates face expressions processing in a priming task. Neuropsychology 2011, 49, 992–998. [Google Scholar] [CrossRef]
- Jonides, J.; Nee, D. Brain mechanisms of proactive interference in working memory. Neuroscience 2006, 139, 181–193. [Google Scholar] [CrossRef]
- Engels, A.S.; Heller, W.; Spielberg, J.M.; Warren, S.L.; Sutton, B.P.; Banich, M.T.; Miller, G.A. Co-occurring anxiety influences patterns of brain activity in depression. Cogn. Affect. Behav. Neurosci. 2010, 10, 141–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardila, A.; Bernal, B.; Rosselli, M. How Localized are Language Brain Areas? A Review of Brodmann Areas Involvement in Oral Language. Arch. Clin. Neuropsychol. 2016, 31, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Leone, A.; Rubio, B.; Pallardó, F.; Catalá, M.D. Rapid-rate transcranial magnetic stimulation of left dorsolateral prefrontal cortex in drug-resistant depression. Lancet 1996, 348, 233–237. [Google Scholar] [CrossRef]
- Brzezicka, A.; Kamiński, J.; Kamińska, O.K.; Wołyńczyk-Gmaj, D.; Sedek, G. Frontal EEG alpha band asymmetry as a predictor of reasoning deficiency in depressed people. Cogn. Emot. 2016, 31, 868–878. [Google Scholar] [CrossRef]
- Mennella, R.; Patron, E.; Palomba, D. Frontal alpha asymmetry neurofeedback for the reduction of negative affect and anxiety. Behav. Res. 2017, 92, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Zotev, V.; Yuan, H.; Misaki, M.; Phillips, R.; Young, K.D.; Feldner, M.T.; Bodurka, J. Correlation between amygdala BOLD activity and frontal EEG asymmetry during real-time fMRI neurofeedback training in patients with depression. NeuroImage Clin. 2016, 11, 224–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, G.; Okamoto, Y.; Morinobu, S.; Yamawaki, S.; Yokota, N. Attenuated Left Prefrontal Activation during a Verbal Fluency Task in Patients with Depression. Neuropsychobiology 2003, 47, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Takamura, M.; Okamoto, Y.; Okada, G.; Toki, S.; Yamamoto, T.; Yamamoto, O.; Jitsuiki, H.; Yokota, N.; Tamura, T.; Kurata, A.; et al. Disrupted Brain Activation and Deactivation Pattern during Semantic Verbal Fluency Task in Patients with Major Depression. Neuropsychobiology 2016, 74, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.C.; Oathes, D.J.; Chang, C.; Bradley, T.; Zhou, Z.-W.; Williams, L.M.; Glover, G.H.; Deisseroth, K.; Etkin, A. Causal interactions between fronto-parietal central executive and default-mode networks in humans. Proc. Natl. Acad. Sci. USA 2013, 110, 19944–19949. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.S.; Strafella, A.P. rTMS of the Left Dorsolateral Prefrontal Cortex Modulates Dopamine Release in the Ipsilateral Anterior Cingulate Cortex and Orbitofrontal Cortex. PLoS ONE 2009, 4, e6725. [Google Scholar] [CrossRef] [Green Version]
- Fox, M.D.; Buckner, R.L.; White, M.P.; Greicius, M.D.; Pascual-Leone, A. Efficacy of Transcranial Magnetic Stimulation Targets for Depression Is Related to Intrinsic Functional Connectivity with the Subgenual Cingulate. Biol. Psychiatry 2012, 72, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Bassett, D.S.; Khambhati, A.N. A network engineering perspective on probing and perturbing cognition with neu-rofeedback. Ann. N. Y. Acad. Sci. 2017, 1396, 126–143. [Google Scholar] [CrossRef] [Green Version]
- Orndorff-Plunkett, F.; Singh, F.; Aragón, O.R.; Pineda, J.A. Assessing the Effectiveness of Neurofeedback Training in the Context of Clinical and Social Neuroscience. Brain Sci. 2017, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Hashimoto, R.I.; Yahata, N.; Ichikawa, N.; Yoshihara, Y.; Okamoto, Y.; Kato, N.; Takahashi, H.; Kawato, M. Resting-State Functional Connectivity-Based Biomarkers and Functional MRI-Based Neurofeedback for Psychiatric Dis-orders: A Challenge for Developing Theranostic Biomarkers. Int. J. Neuropsychopharmacol. 2017, 20, 769–781. [Google Scholar] [CrossRef]
- Rush, A.J.; Trivedi, M.H.; Wisniewski, S.R.; Nierenberg, A.A.; Stewart, J.W.; Warden, D.; Niederehe, G.; Thase, M.E.; Lavori, P.W.; Lebowitz, B.D.; et al. Acute and longer-term outcomes in depressed outpatients requiring one or several treatment steps: A STAR*D report. Am. J. Psychiatry 2006, 163, 1905–1917. [Google Scholar] [CrossRef]
- Arroll, B.; Macgillivray, S.; Ogston, S.; Reid, I.; Sullivan, F.; Williams, B.; Crombie, I. Efficacy and tolerability of tricyclic antidepressants and SSRIs compared with placebo for treatment of depression in primary care: A meta-analysis. Ann. Fam. Med. 2005, 3, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Gabrieli, J.D.; Ghosh, S.S.; Whitfield-Gabrieli, S. Prediction as a Humanitarian and Pragmatic Contribution from Human Cognitive Neuroscience. Neuron 2015, 85, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Stephan, K.; Schlagenhauf, F.; Huys, Q.; Raman, S.; Aponte, E.; Brodersen, K.; Rigoux, L.; Moran, R.; Daunizeau, J.; Dolan, R.; et al. Computational neuroimaging strategies for single patient predictions. NeuroImage 2017, 145, 180–199. [Google Scholar] [CrossRef] [Green Version]
- Freyhan, F.A. Course and outcome of schizophrenia. Am. J. Psychiatry 1955, 112, 161–169. [Google Scholar] [CrossRef]
- Häfner, H.; Der Heiden, W.A. The course of schizophrenia in the light of modern follow-up studies: The ABC and WHO studies. Eur. Arch. Psychiatry Clin. Neurosci. 1999, 249, S14–S26. [Google Scholar] [CrossRef]
- Kane, J.M.; Correll, C.U. Past and Present Progress in the Pharmacologic Treatment of Schizophrenia. J. Clin. Psychiatry 2010, 71, 1115–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourao-Miranda, J.; Reinders, A.A.T.S.; Rocha-Rego, V.; Lappin, J.; Rondina, J.; Morgan, C.; Morgan, K.D.; Fearon, P.; Jones, P.B.; Doody, G.A.; et al. Individualized prediction of illness course at the first psychotic episode: A support vector machine MRI study. Psychol. Med. 2011, 42, 1037–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, H.W.H.; Leung, A.Y.; Chung, R.C.K.; Bell, M.; Cheung, W.-M. Review on vocational predictors: A systematic review of predictors of vocational outcomes among individuals with schizophrenia: An update since 1998. Aust. N. Z. J. Psychiatry 2010, 44, 495–504. [Google Scholar]
- Jääskeläinen, E.; Juola, P.; Hirvonen, N.; McGrath, J.J.; Saha, S.; Isohanni, M.; Veijola, J.; Miettunen, J. A Systematic Review and Meta-Analysis of Recovery in Schizophrenia. Schizophr. Bull. 2013, 39, 1296–1306. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Zalesky, A.; Yue, W.; Howes, O.; Yan, H.; Liu, Y.; Fan, L.; Whitaker, K.J.; Xu, K.; Rao, G.; et al. A neu-roimaging biomarker for striatal dysfunction in schizophrenia. Nat. Med. 2020, 26, 558–565. [Google Scholar] [CrossRef]
- Patel, M.J.; Andreescu, C.; Price, J.C.; Edelman, K.L.; Reynolds, C.F., 3rd; Aizenstein, H.J. Machine learning approaches for integrating clinical and imaging features in late-life depression classification and response prediction. Int. J. Geriatr. Psychiatry 2015, 30, 1056–1067. [Google Scholar] [CrossRef]
- Redlich, R.; Opel, N.; Grotegerd, D.; Dohm, K.; Zaremba, D.; Bürger, C.; Münker, S.; Mühlmann, L.; Wahl, P.; Heindel, W.; et al. Prediction of Individual Response to Electroconvulsive Therapy via Machine Learning on Structural Magnetic Resonance Imaging Data. JAMA Psychiatry 2016, 73, 557–564. [Google Scholar] [CrossRef]
- Wade, B.S.C.; Joshi, S.H.; Njau, S.; Leaver, A.M.; Vasavada, M.; Woods, R.P.; A Gutman, B.; Thompson, P.M.; Espinoza, R.; Narr, K.L. Effect of Electroconvulsive Therapy on Striatal Morphometry in Major Depressive Disorder. Neuropsychopharmacology 2016, 41, 2481–2491. [Google Scholar] [CrossRef] [Green Version]
- Iosifescu, D.V. Electroencephalography-Derived Biomarkers of Antidepressant Response. Harv. Rev. Psychiatry 2011, 19, 144–154. [Google Scholar] [CrossRef]
- Baskaran, A.; Milev, R.; McIntyre, R.S. The neurobiology of the EEG biomarker as a predictor of treatment response in depression. Neuropharmacology 2012, 63, 507–513. [Google Scholar] [CrossRef]
- Leuchter, A.F.; Cook, I.A.; Uijtdehaage, S.H.; Dunkin, J.; Lufkin, R.B.; Anderson-Hanley, C.; Abrams, M.; Rosenberg-Thompson, S.; O’Hara, R.; Simon, S.L.; et al. Brain structure and function and the outcomes of treatment for depression. J. Clin. Psychiatry 1997, 58, 22–31. [Google Scholar]
- Rabinoff, M.; Kitchen, C.; Cook, I.; Leuchter, A. Evaluation of Quantitative EEG by Classification and Regression Trees to Characterize Responders to Antidepressant and Placebo Treatment. Open Med. Inform. J. 2011, 5, 1–8. [Google Scholar] [CrossRef]
- Bareš, M.; Brunovsky, M.; Kopecek, M.; Novak, T.; Stopkova, P.; Kožený, J.; Šóš, P.; Krajca, V.; Höschl, C. Early reduction in prefrontal theta QEEG cordance value predicts response to venlafaxine treatment in patients with resistant depressive disorder. Eur. Psychiatry 2008, 23, 350–355. [Google Scholar] [CrossRef]
- Cook, I.A.; Leuchter, A.F.; Morgan, M.L.; Stubbeman, W.; Siegman, B.; Abrams, M. Changes in prefrontal activity characterize clinical response in SSRI nonresponders: A pilot study. J. Psychiatr. Res. 2005, 39, 461–466. [Google Scholar] [CrossRef]
- Knott, V.J.; I Telner, J.; Lapierre, Y.D.; Browne, M.; Horn, E.R. Quantitative EEG in the prediction of antidepressant response to imipramine. J. Affect. Disord. 1996, 39, 175–184. [Google Scholar] [CrossRef]
- Bruder, G.E.; Sedoruk, J.P.; Stewart, J.W.; McGrath, P.J.; Quitkin, F.M.; Tenke, C.E. Electroencephalographic Alpha Measures Predict Therapeutic Response to a Selective Serotonin Reuptake Inhibitor Antidepressant: Pre- and Post-Treatment Findings. Biol. Psychiatry 2008, 63, 1171–1177. [Google Scholar] [CrossRef] [Green Version]
- Ulrich, G.; Renfordt, E.; Zeller, G.; Frick, K. Interrelation between Changes in the EEG and Psychopathology under Pharmacotherapy for Endogenous Depression. Pharmacopsychiatry 1984, 17, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Iosifescu, D.V.; Greenwald, S.; Devlin, P.; Mischoulon, D.; Denninger, J.W.; Alpert, J.E.; Fava, M. Frontal EEG predictors of treatment outcome in major depressive disorder. Eur. Neuropsychopharmacol. 2009, 19, 772–777. [Google Scholar] [CrossRef]
- Iosifescu, D.V.; Nierenberg, A.A.; Mischoulon, D.; Perlis, R.H.; Papakostas, G.I.; Ryan, J.L.; Alpert, J.E.; Fava, M. An open study of triiodothyronine augmentation of selective serotonin reuptake inhibitors in treatment-resistant major de-pressive disorder. J. Clin. Psychiatry 2005, 66, 1038–1042. [Google Scholar] [CrossRef]
- Bruder, G.E.; Stewart, J.W.; Tenke, C.E.; McGrath, P.J.; Leite, P.; Bhattacharya, N.; Quitkin, F.M. Electroencephalo-graphic and perceptual asymmetry differences between responders and nonresponders to an SSRI antidepressant. Biol. Psychiatry 2001, 49, 416–425. [Google Scholar] [CrossRef]
- Knott, V.; Mahoney, C.; Kennedy, S.; Evans, K. Pre-treatment EEG and it’s relationship to depression severity and par-oxetine treatment outcome. Pharmacopsychiatry 2000, 33, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Korb, A.S.; Hunter, A.M.; Cook, I.A.; Leuchter, A.F. Rostral anterior cingulate cortex theta current density and response to antidepressants and placebo in major depression. Clin. Neurophysiol. 2009, 120, 1313–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzagalli, D.; Pascual-Marqui, R.D.; Nitschke, J.B.; Oakes, T.R.; Larson, C.L.; Abercrombie, H.C.; Schaefer, S.M.; Koger, J.V.; Benca, R.M.; Davidson, R.J. Anterior cingulate activity as a predictor of degree of treatment response in major de-pression: Evidence from brain electrical tomography analysis. Am. J. Psychiatry 2001, 158, 405–415. [Google Scholar] [CrossRef]
- Méndez, M.A.; Zuluaga, P.; Hornero, R.; Gómez, C.; Escudero, J.; Rodríguez-Palancas, A.; Ortiz, T.; Fernández, A. Complexity analysis of spontaneous brain activity: Effects of depression and antidepressant treatment. J. Psychopharmacol. 2011, 26, 636–643. [Google Scholar] [CrossRef] [Green Version]
- Jaworska, N.; Wang, H.; Smith, D.M.; Blier, P.; Knott, V.; Protzner, A.B. Pre-treatment EEG signal variability is associated with treatment success in depression. NeuroImage Clin. 2018, 17, 368–377. [Google Scholar] [CrossRef]
- Thomasson, N.; Pezard, L. Dynamical systems and depression: A framework for theoretical perspectives. Acta Biotheor. 1999, 47, 209–218. [Google Scholar] [CrossRef]
- Zhdanov, A.; Atluri, S.; Wong, W.; Vaghei, Y.; Daskalakis, Z.J.; Blumberger, D.M.; Frey, B.N.; Giacobbe, P.; Lam, R.W.; Milev, R.; et al. Use of Machine Learning for Predicting Escitalopram Treatment Outcome From Electroencephalography Recordings in Adult Patients With Depression. JAMA Netw. Open 2020, 3, e1918377. [Google Scholar] [CrossRef]
- Hasanzadeh, F.; Mohebbi, M.; Rostami, R. Prediction of rTMS treatment response in major depressive disorder using machine learning techniques and nonlinear features of EEG signal. J. Affect. Disord. 2019, 256, 132–142. [Google Scholar] [CrossRef]
- Garg, G.; Elshimy, G.; Marwaha, R. Gender Dysphoria (Sexual Identity Disorders); StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Petry, N.M.; Rehbein, F.; Ko, C.-H.; O’Brien, C.P. Internet Gaming Disorder in the DSM-5. Curr. Psychiatry Rep. 2015, 17, 1–9. [Google Scholar] [CrossRef]
- Ross, C.A.; Margolis, R.L. Research Domain Criteria: Strengths, Weaknesses, and Potential Alternatives for Future Psy-chiatric Research. Mol. Neuropsychiatry 2019, 5, 218–236. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No | Anatomical Name | No | Anatomical Name |
|---|---|---|---|
| 1 | anterior lateral fissure | 36 | posterior sub-central ramus of the lateral fissure |
| 2 | anterior ramus of the lateral fissure | 37 | calloso-marginal posterior fissure |
| 3 | diagonal ramus of the lateral fissure | 38 | collateral fissure |
| 4 | anterior sub-central ramus of the lateral fissure | 39 | intraparietal sulcus |
| 5 | calloso-marginal anterior fissure | 40 | secondary intermediate ramus of the intraparietal sulcus |
| 6 | calcarine fissure | 41 | insula |
| 7 | superior postcentral intraparietal superior sulcus | 42 | paracentral lobule central sulcus |
| 8 | primary intermediate ramus of the intraparietal sulcus | 43 | central sylvian sulcus |
| 9 | parieto-occipital fissure | 44 | cuneal sulcus |
| 10 | lobe occipital | 45 | anterior interior frontal sulcus |
| 11 | central sulcus | 46 | intermediate frontal sulcus |
| 12 | subcallosal sulcus | 47 | median frontal sulcus |
| 13 | inferior frontal sulcus | 48 | polar frontal sulcus |
| 14 | internal frontal sulcus | 49 | sulcus of the supra-marginal gyrus |
| 15 | marginal frontal sulcus | 50 | posterior intra-lingual sulcus |
| 16 | orbital frontal sulcus | 51 | internal occipito-temporal lateral sulcus |
| 17 | superior frontal sulcus | 52 | posterior occipito-temporal lateral sulcus |
| 18 | anterior intralingual sulcus | 53 | olfactory sulcus |
| 19 | anterior occipito-temporal lateral sulcus | 54 | internal parietal sulcus |
| 20 | median occipito-temporal lateral sulcus | 55 | transverse precentral sulcus |
| 21 | occipito-polar sulcus | 56 | intermediate precentral sulcus |
| 22 | orbital sulcus | 57 | median precentral sulcus |
| 23 | superior parietal sulcus | 58 | superior postcentral sulcus |
| 24 | interior precentral sulcus | 59 | rhinal sulcus |
| 25 | marginal precentral sulcus | 60 | posterior inferior temporal sulcus |
| 26 | superior precentral sulcus | 61 | superior temporal sulcus |
| 27 | inferior rostral sulcus | 62 | superior terminal ascending branch of the superior temporal sulcus |
| 28 | anterior inferior temporal sulcus | 63 | sub-parietal sulcus |
| 29 | polar temporal sulcus | 64 | Thalamus |
| 30 | anterior terminal ascending branch of the superior temporal sulcus | 65 | Amygdala |
| 31 | paracentral sulcus | 66 | Accumbens |
| 32 | ventricle | 67 | Caudate |
| 33 | posterior lateral fissure | 68 | Pallidum |
| 34 | ascending ramus of the lateral fissure | 69 | Putamen |
| 35 | retro central transverse ramus of the lateral fissure | 70 | Vermis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komatsu, H.; Watanabe, E.; Fukuchi, M. Psychiatric Neural Networks and Precision Therapeutics by Machine Learning. Biomedicines 2021, 9, 403. https://doi.org/10.3390/biomedicines9040403
Komatsu H, Watanabe E, Fukuchi M. Psychiatric Neural Networks and Precision Therapeutics by Machine Learning. Biomedicines. 2021; 9(4):403. https://doi.org/10.3390/biomedicines9040403
Chicago/Turabian StyleKomatsu, Hidetoshi, Emi Watanabe, and Mamoru Fukuchi. 2021. "Psychiatric Neural Networks and Precision Therapeutics by Machine Learning" Biomedicines 9, no. 4: 403. https://doi.org/10.3390/biomedicines9040403