
Asiatic Acid Prevents Cognitive Deficits by Inhibiting Calpain Activation and Preserving Synaptic and Mitochondrial Function in Rats with Kainic Acid-Induced Seizure

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Experimental Design
2.4. Morris Water Maze Test
2.5. Neutral Red and FJB Staining
2.6. Synaptosomal Preparation and Transmission Electron Microscopy
2.7. Proteomics
2.8. Biological Network Analysis Using MetaCore™
2.9. Western Blotting
2.10. Statistical Analyses
3. Results
3.1. Asiatic Acid Reduced the Mortality Rate and Seizures in Rats Receiving KA
3.2. Asiatic Acid Improved Learning and Memory Deficits in Rats with KA-Induced Seizures
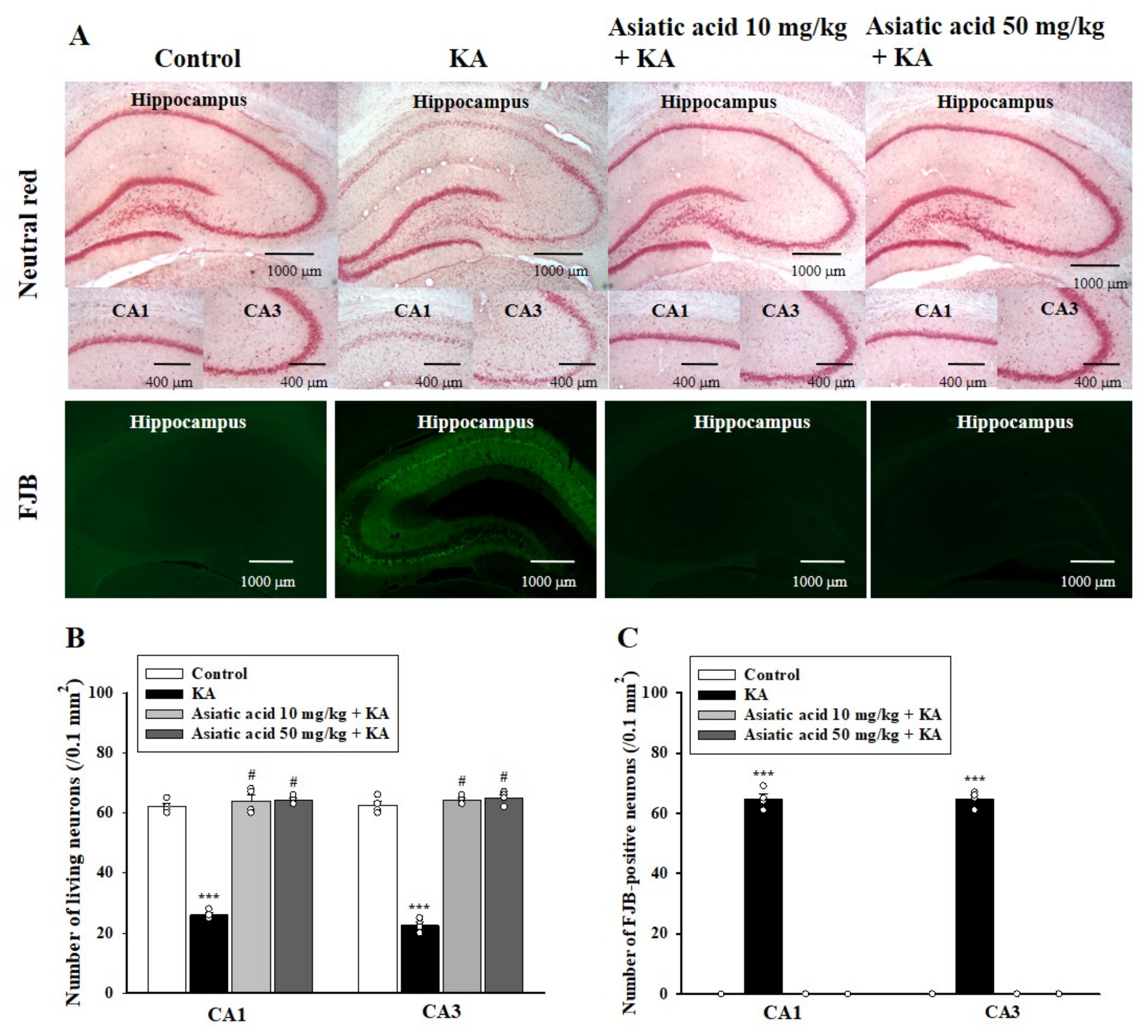
3.3. Asiatic Acid Attenuated Neuronal Damage in the Hippocampus of Rats with KA-Induced Seizures
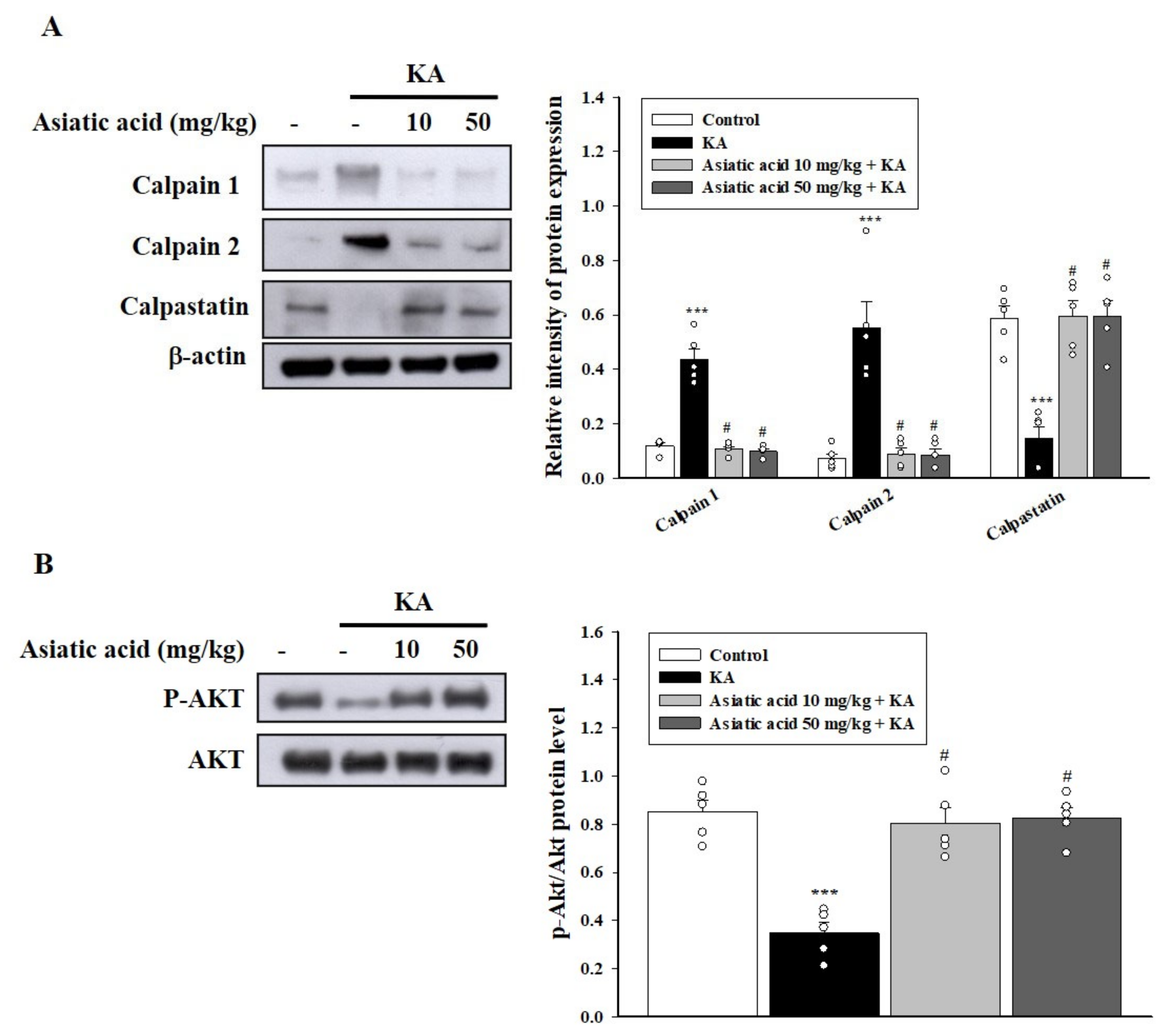
3.4. Asiatic Acid Suppressed Calpain Activation and AKT Inactivation in the Hippocampus of Rats with KA-Induced Seizures
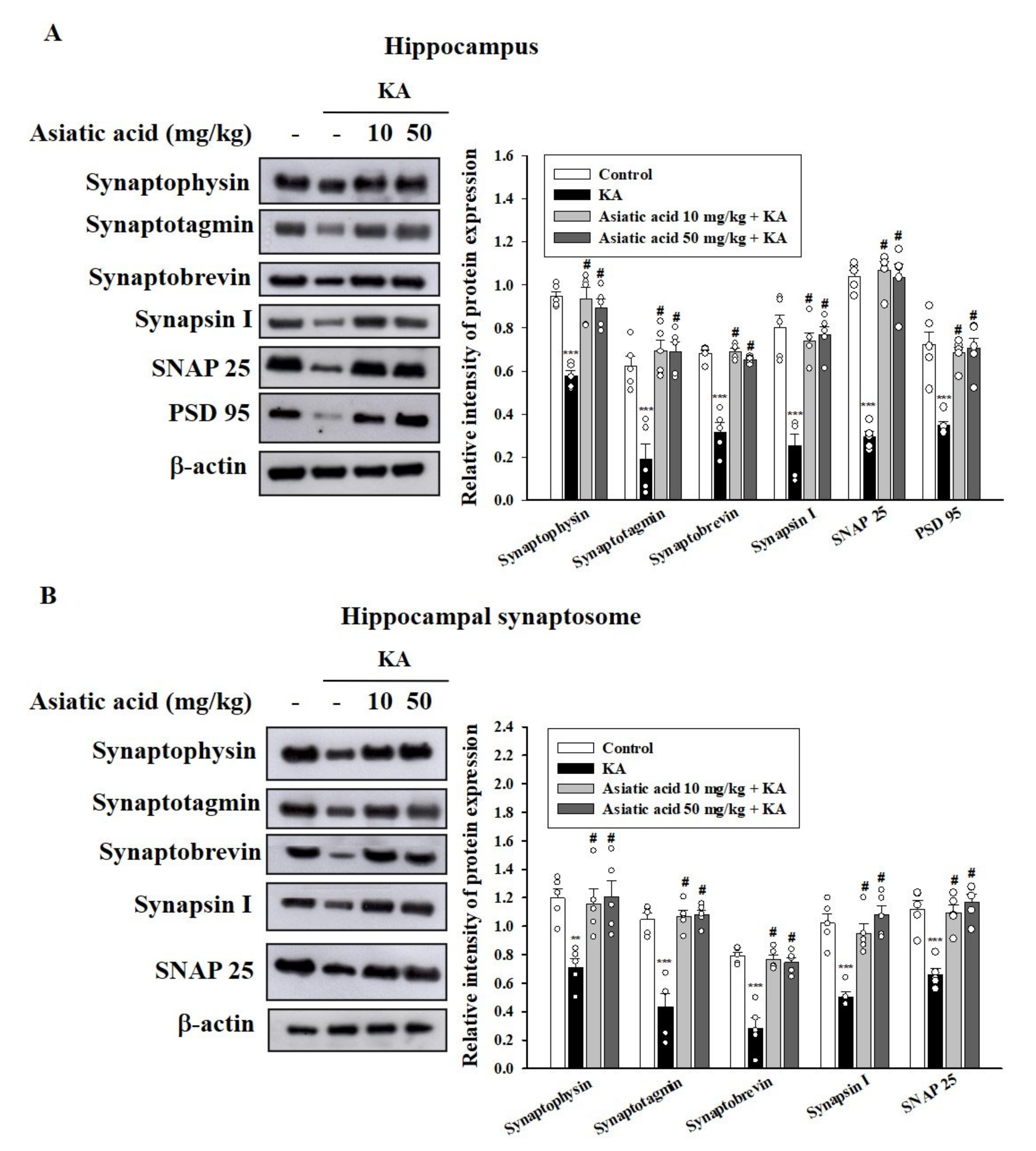
3.5. Asiatic Acid Preserved the Levels of Synaptic Proteins in the Hippocampus of Rats with KA-Induced Seizures
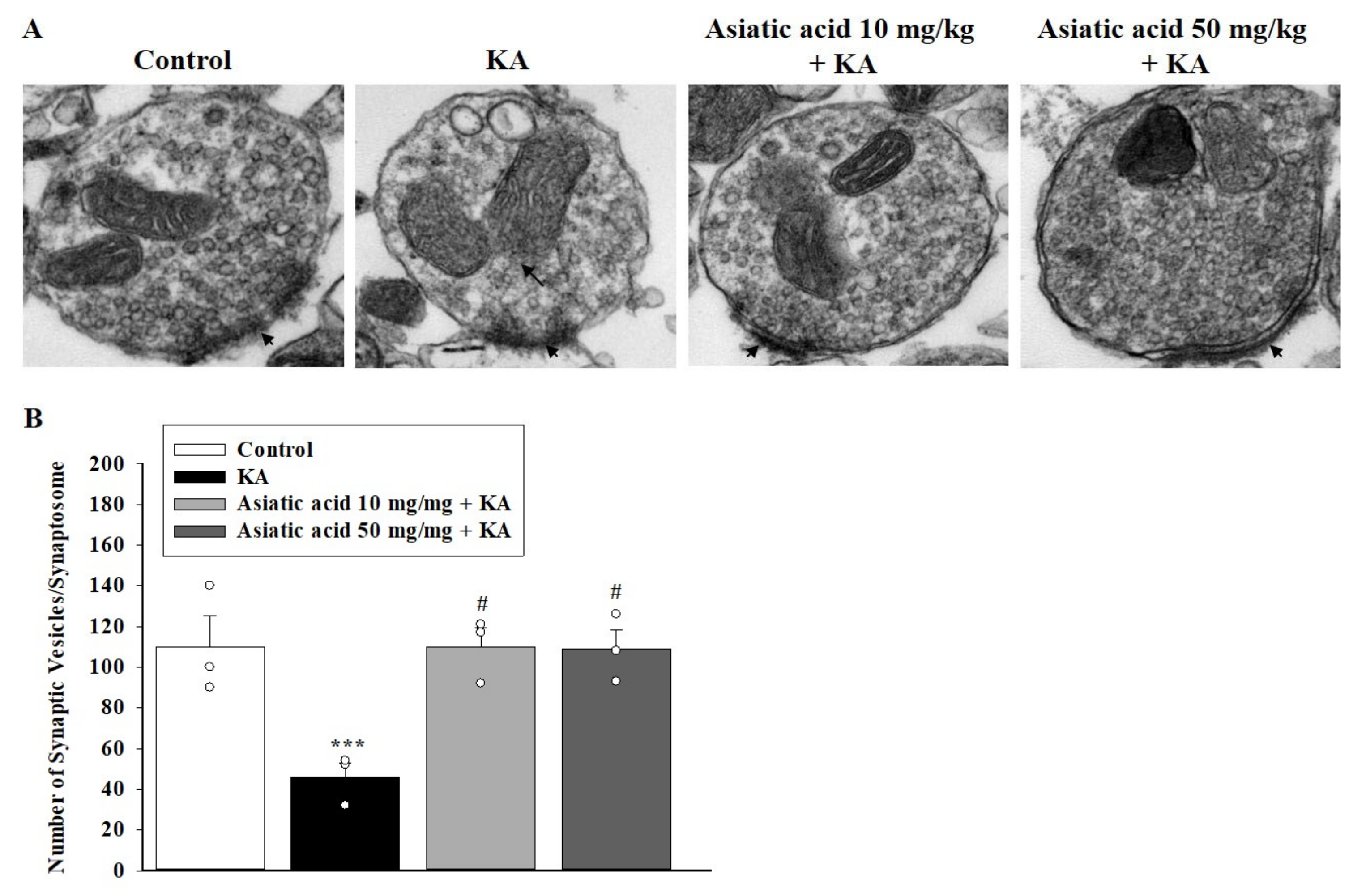
3.6. Asiatic Acid Prevented the Decline in Synaptic Vesicles and Mitochondrial Morphology Damage in the Hippocampal Synaptosomes of Rats with KA-Induced Seizures
3.7. Proteomic Analysis Identified 12 Proteins Associated with the Effect of Asiatic Acid on the Hippocampal Synaptosomes of Rats with KA-Induced Seizures
3.8. Functional Network Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moshé, S.L.; Perucca, E.; Ryvlin, P.; Tomson, T. Epilepsy: New advances. Lancet 2015, 385, 884–898. [Google Scholar] [CrossRef]
- Elger, C.E.; Helmstaedter, C.; Kurthen, M. Chronic epilepsy and cognition. Lancet Neurol. 2004, 3, 663–672. [Google Scholar] [CrossRef]
- Lin, J.J.; Mula, M.; Hermann, B.P. Uncovering the neurobehavioural comorbidities of epilepsy over the lifespan. Lancet 2012, 380, 1180–1192. [Google Scholar] [CrossRef] [Green Version]
- Witt, J.A.; Helmstaedter, C. Cognition in epilepsy: Current clinical issues of interest. Curr. Opin. Neurol. 2017, 30, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Ortinski, P.; Meador, K.J. Cognitive side effects of antiepileptic drugs. Epilepsy Behav. 2004, 5 (Suppl. 1), S60–S65. [Google Scholar] [CrossRef]
- Santulli, L.; Coppola, A.; Balestrini, S.; Striano, S. The challenges of treating epilepsy with 25 antiepileptic drugs. Pharmacol. Res. 2016, 107, 211–219. [Google Scholar] [CrossRef]
- Zhao, Z.; He, X.; Ma, C.; Wu, S.; Cuan, Y.; Sun, Y.; Bai, Y.; Huang, L.; Chen, X.; Gao, T.; et al. Excavating anticonvulsant compounds from prescriptions of traditional chinese medicine in the treatment of epilepsy. Am. J. Chin. Med. 2018, 46, 707–737. [Google Scholar] [CrossRef]
- Auditeau, E.; Chassagne, F.; Bourdy, G.; Bounlu, M.; Jost, J.; Luna, J.; Ratsimbazafy, V.; Preux, P.M.; Boumediene, F. Herbal medicine for epilepsy seizures in Asia, Africa and Latin America: A systematic review. J. Ethnopharmacol. 2019, 234, 119–153. [Google Scholar] [CrossRef]
- Hashim, P.; Sidek, H.; Helan, M.H.; Sabery, A.; Palanisamy, U.D.; Ilham, M. Triterpene composition and bioactivities of Centella asiatica. Molecules 2011, 16, 1310–1322. [Google Scholar] [CrossRef]
- Xu, M.F.; Xiong, Y.Y.; Liu, J.K.; Qian, J.J.; Zhu, L.; Gao, J. Asiatic acid, a pentacyclic triterpene in Centella asiatica, attenuates glutamate-induced cognitive deficits in mice and apoptosis in SH-SY5Y cells. Acta. Pharmacol. Sin. 2012, 33, 578–587. [Google Scholar] [CrossRef] [Green Version]
- Sirichoat, A.; Chaijaroonkhanarak, W.; Prachaney, P.; Pannangrong, W.; Leksomboon, R.; Chaichun, A.; Wigmore, P.; Welbat, J.U. Effects of asiatic acid on spatial working memory and cell proliferation in the adult rat hippocampus. Nutrients 2015, 7, 8413–8423. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Li, M.; He, F.; Bian, Z.; He, Q.; Wang, X.; Yao, W.; Zhu, L. Neuroprotective effect of asiatic acid against spinal cord injury in rats. Life Sci. 2016, 157, 45–51. [Google Scholar] [CrossRef]
- Nataraj, J.; Manivasagam, T.; Justin Thenmozhi, A.; Essa, M.M. Neuroprotective effect of asiatic acid on rotenone-induced mitochondrial dysfunction and oxidative stress-mediated apoptosis in differentiated SH-SYS5Y cells. Nutr. Neurosci. 2017, 20, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, J.; Manivasagam, T.; Justin Thenmozhi, A.; Essa, M.M. Neurotrophic effect of asiatic acid, a triterpene of Centella asiatica against chronic 1-methyl 4-phenyl 1, 2, 3, 6-tetrahydropyridine hydrochloride/probenecid mouse model of Parkinson’s disease: The role of MAPK, PI3K-Akt-GSK3β and mTOR signalling pathways. Neurochem. Res. 2017, 42, 1354–1365. [Google Scholar] [CrossRef]
- Wang, Z.H.; Mong, M.C.; Yang, Y.C.; Yin, M.C. Asiatic acid and maslinic acid attenuated kainic acid-induced seizure through decreasing hippocampal inflammatory and oxidative stress. Epilepsy Res. 2018, 139, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Loganathan, C.; Thayumanavan, P. Asiatic acid prevents the quinolinic acid-induced oxidative stress and cognitive impairment. Metab. Brain Dis. 2018, 33, 151–159. [Google Scholar] [CrossRef]
- Ahmad Rather, M.; Justin-Thenmozhi, A.; Manivasagam, T.; Saravanababu, C.; Guillemin, G.J.; Essa, M.M. Asiatic acid attenuated aluminum chloride-induced tau pathology, oxidative stress and apoptosis via AKT/GSK-3β signaling pathway in wistar rats. Neurotox. Res. 2019, 35, 955–968. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.W.; Lin, T.Y.; Wang, S.J.; Huang, S.K. Asiatic acid, an active substance of Centella asiatica, presynaptically depresses glutamate release in the rat hippocampus. Eur. J. Pharmacol. 2019, 865, 172781. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, R.G.; Senut, M.C.; Zemke, D.; Min, J.; Frenkel, M.B.; Greenberg, E.J.; Yu, S.W.; Ahn, N.; Goudreau, J.; Kassab, M.; et al. Asiatic acid, a pentacyclic triterpene from Centella asiatica, is neuroprotective in a mouse model of focal cerebral ischemia. J. Neurosci. Res. 2009, 87, 2541–2550. [Google Scholar] [CrossRef] [Green Version]
- Raval, N.; Mistry, T.; Acharya, N.; Acharya, S. Development of glutathione-conjugated asiatic acid-loaded bovine serum albumin nanoparticles for brain-targeted drug delivery. J. Pharm. Pharmacol. 2015, 67, 1503–1511. [Google Scholar] [CrossRef]
- Kim, S.; Jung, U.J.; Oh, Y.S.; Jeon, M.T.; Kim, H.J.; Shin, W.H.; Hong, J.; Kim, S.R. Beneficial effects of silibinin against kainic acid-induced neurotoxicity in the hippocampus in vivo. Exp. Neurobiol. 2017, 26, 266–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemi, P.; FahanikBabaei, J.; Vazifekhah, S.; Nikbakht, F. Evaluation of the neuroprotective, anticonvulsant, and cognition-improvement effects of apigenin in temporal lobe epilepsy: Involvement of the mitochondrial apoptotic pathway. Iran J. Basic Med. Sci. 2019, 22, 752–758. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, J.; Huang, S.; Zhu, W.; Wang, Y.; Chen, O.; Xue, J. Neuroprotective effects of isoliquiritigenin against cognitive impairment via suppression of synaptic dysfunction, neuronal injury, and neuroinflammation in rats with kainic acid-induced seizures. Int. Immunopharmacol. 2019, 72, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Nadler, J.V. Minireview. Kainic acid as a tool for the study of temporal lobe epilepsy. Life Sci. 1981, 29, 2031–2042. [Google Scholar] [CrossRef]
- Ben-Ari, Y. Limbic seizure and brain damage produced by kainic acid: Mechanisms and relevance to human temporal lobe epilepsy. Neuroscience 1985, 14, 375–403. [Google Scholar] [CrossRef]
- Lévesque, M.; Avoli, M. The kainic acid model of temporal lobe epilepsy. NeurosciBiobehavRev. 2013, 37, 2887–2899. [Google Scholar] [CrossRef] [Green Version]
- Racine, R.J. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 281–294. [Google Scholar] [CrossRef]
- Friedman, L.K.; Pellegrini-Giampietro, D.E.; Sperber, E.F.; Bennett, M.V.; Moshé, S.L.; Zukin, R.S. Kainate-induced status epilepticus alters glutamate and GABAA receptor gene expression in adult rat hippocampus: An in situ hybridization study. J. Neurosci. 1994, 14, 2697–2707. [Google Scholar] [CrossRef]
- Chang, Y.; Lu, C.W.; Chen, Y.J.; Lin, T.Y.; Huang, S.K.; Wang, S.J. Astaxanthin protects against kainic acid-induced seizures and pathological consequences. Neurochem. Int. 2018, 116, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.W.; Lin, T.Y.; Chiu, K.M.; Lee, M.Y.; Huang, J.H.; Wang, S.J. Silymarin inhibits glutamate release and prevents against kainic acid-induced excitotoxic injury in rats. Biomedicines 2020, 8. [Google Scholar] [CrossRef]
- Lin, T.Y.; Lu, C.W.; Wang, S.J. Luteolin protects the hippocampus against neuron impairments induced by kainic acid in rats. Neurotoxicology 2016, 55, 48–57. [Google Scholar] [CrossRef]
- Lin, T.Y.; Lu, C.W.; Wu, C.C.; Huang, S.K.; Wang, S.J. Palmitoylethanolamide inhibits glutamate release in rat cerebrocortical nerve terminals. Int. J. Mol. Sci. 2015, 16, 5555–5571. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.Y.; Wang, P.W.; Huang, C.H.; Yang, P.M.; Pan, T.L. Characterizing the relapse potential in different luminal subtypes of breast cancers with functional proteomics. Int. J. Mol. Sci. 2020, 21, 6077. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.W.; Hung, Y.C.; Lin, T.Y.; Fang, J.Y.; Yang, P.M.; Chen, M.H.; Pan, T.L. Comparison of the biological impact of UVA and UVB upon the skin with functional proteomics and immunohistochemistry. Antioxidants 2019, 8, 569. [Google Scholar] [CrossRef] [Green Version]
- Hung, Y.C.; Wang, P.W.; Lin, T.Y.; Yang, P.M.; You, J.S.; Pan, T.L. Functional redox proteomics reveal that Salvia miltiorrhiza aqueous extract alleviates adriamycin-induced cardiomyopathy via inhibiting ROS-dependent apoptosis. Oxid. Med. Cell Longev. 2020, 2020, 5136934. [Google Scholar] [CrossRef]
- Lu, C.W.; Lin, T.Y.; Huang, S.K.; Wang, S.J. Echinacoside Inhibits Glutamate Release by Suppressing Voltage-Dependent Ca(2+) Entry and Protein Kinase C in Rat Cerebrocortical Nerve Terminals. Int. J. Mol. Sci. 2016, 17, 1006. [Google Scholar] [CrossRef] [Green Version]
- Maia, G.H.; Quesado, J.L.; Soares, J.I.; do Carmo, J.M.; Andrade, P.A.; Andrade, J.P.; Lukoyanov, N.V. Loss of hippocampal neurons after kainate treatment correlates with behavioral deficits. PLoS ONE 2014, 9, e84722. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.Y.; Na, E.J.; Lee, E.; Kwon, Y.; Kim, H.J. Antiepileptic and neuroprotective effects of oleamide in rat striatum on kainate-induced behavioral seizure and excitotoxic damage via calpain inhibition. Front. Pharmacol. 2017, 8, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, P.M.; González, M.I. Calpain activation and neuronal death during early epileptogenesis. Neurobiol. Dis. 2019, 124, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Machado, V.M.; Lourenço, A.S.; Florindo, C.; Fernandes, R.; Carvalho, C.M.; Araújo, I.M. Calpastatin overexpression preserves cognitive function following seizures, while maintaining post-injury neurogenesis. Front. Mol. Neurosci. 2017, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Chun, W.; Kong, P.J.; Han, J.A.; Cho, B.P.; Kwon, O.Y.; Lee, H.J.; Kim, S.S. Sustained activation of Akt by melatonin contributes to the protection against kainic acid-induced neuronal death in hippocampus. J. Pineal Res. 2006, 40, 79–85. [Google Scholar] [CrossRef]
- Piermartiri, T.C.; Vandresen-Filho, S.; de Araújo Herculano, B.; Martins, W.C.; Dal’agnolo, D.; Stroeh, E.; Carqueja, C.L.; Boeck, C.R.; Tasca, C.I. Atorvastatin prevents hippocampal cell death due to quinolinic acid-induced seizures in mice by increasing Akt phosphorylation and glutamate uptake. Neurotox. Res. 2009, 16, 106–115. [Google Scholar] [CrossRef]
- Zhang, F.X.; Sun, Q.J.; Zheng, X.Y.; Lin, Y.T.; Shang, W.; Wang, A.H.; Duan, R.S.; Chi, Z.F. Abnormal expression of synaptophysin, SNAP-25, and synaptotagmin 1 in the hippocampus of kainic acid-exposed rats with behavioral deficits. Cell Mol. Neurobiol. 2014, 34, 813–824. [Google Scholar] [CrossRef]
- Shi, C.; Zeng, J.; Li, Z.; Chen, Q.; Hang, W.; Xia, L.; Wu, Y.; Chen, J.; Shi, A. Melatonin mitigates kainic acid-induced neuronal tau hyperphosphorylation and memory deficits through alleviating ER stress. Front. Mol. Neurosci. 2018, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Kundap, U.P.; Paudel, Y.N.; Kumari, Y.; Othman, I.; Shaikh, M.F. Embelin prevents seizure and associated cognitive impairments in a pentylenetetrazole-induced kindling zebrafish model. Front. Pharmacol. 2019, 10, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, C.; Han, S.; Wei, L.; Dang, X.; Niu, Q.; Chen, M.; Cao, B.; Liu, Y.; Jiao, H. Protective effect of compound Danshen (Salvia miltiorrhiza) dripping pills alone and in combination with carbamazepine on kainic acid-induced temporal lobe epilepsy and cognitive impairment in rats. Pharm. Biol. 2018, 56, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Q.; Zeng, C.; Luo, C.; Wu, Y. Ferrostatin-1 mitigates cognitive impairment of epileptic rats by inhibiting P38 MAPK activation. Epilepsy Behav. 2020, 103, 106670. [Google Scholar] [CrossRef]
- Kim, S.R.; Koo, K.A.; Lee, M.K.; Park, H.G.; Jew, S.S.; Cha, K.H.; Kim, Y.C. Asiatic acid derivatives enhance cognitive performance partly by improving acetylcholine synthesis. J. Pharm. Pharmacol. 2004, 56, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Aiyer, R.S.; Nath, J.R. Mechanisms of memory deficit in temporal lobe epilepsy. NZMSJ 2011, 22, 6052–6061. [Google Scholar]
- Titiz, A.S.; Mahoney, J.M.; Testorf, M.E.; Holmes, G.L.; Scott, R.C. Cognitive impairment in temporal lobe epilepsy: Role of online and offline processing of single cell information. Hippocampus 2014, 24, 1129–1145. [Google Scholar] [CrossRef] [Green Version]
- Xue, F.; Shi, C.; Chen, Q.; Hang, W.; Xia, L.; Wu, Y.; Tao, S.Z.; Zhou, J.; Shi, A.; Chen, J. Melatonin mediates protective effects against kainic acid-induced neuronal death through safeguarding ER stress and mitochondrial disturbance. Front. Mol. Neurosci. 2017, 10, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Han, Y.; Zhao, Y.; Li, Q.; Jin, H.; Qin, J. Inhibition of TRIB3 protects against neurotoxic injury induced by kainic acid in rats. Front. Pharmacol. 2019, 10, 585. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Chen, W.; Wang, P.; Chu, J. Asiatic acid protects differentiated PC12 cells from Aβ(25-35)-induced apoptosis and tau hyperphosphorylation via regulating PI3K/Akt/GSK-3β signaling. Life Sci. 2018, 208, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Grimwood, P.D.; Morris, R.G. Synaptic plasticity and memory: An evaluation of the hypothesis. AnnuRev. Neurosci. 2000, 23, 649–711. [Google Scholar] [CrossRef] [Green Version]
- Antonova, I.; Arancio, O.; Trillat, A.C.; Wang, H.G.; Zablow, L.; Udo, H.; Kandel, E.R.; Hawkins, R.D. Rapid increase in clusters of presynaptic proteins at onset of long-lasting potentiation. Science 2001, 294, 1547–1550. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.A.; Voss, K.L.; Rodriguez, J.; Bliss, T.V. Increase in synaptic vesicle proteins accompanies long-term potentiation in the dentate gyrus. Neuroscience 1994, 60, 1–5. [Google Scholar] [CrossRef]
- Mullany, P.; Lynch, M.A. Changes in protein synthesis and synthesis of the synaptic vesicle protein, synaptophysin, in entorhinal cortex following induction of long-term potentiation in dentate gyrus: An age-related study in the rat. Neuropharmacology 1997, 36, 973–980. [Google Scholar] [CrossRef]
- Saatman, K.E.; Bozyczko-Coyne, D.; Marcy, V.; Siman, R.; McIntosh, T.K. Prolonged calpain-mediated spectrin breakdown occurs regionally following experimental brain injury in the rat. J. Neuropathol. Exp. Neurol. 1996, 55, 850–860. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.C.; Baker, J.S.; Donti, T.; Graham, B.H.; Craigen, W.J.; Anderson, A.E. Mitochondrial Dysfunction Mediated by Poly(ADP-Ribose) Polymerase-1 Activation Contributes to Hippocampal Neuronal Damage Following Status Epilepticus. Int. J. Mol. Sci. 2017, 18, 1502. [Google Scholar] [CrossRef]
- Gano, L.B.; Liang, L.P.; Ryan, K.; Michel, C.R.; Gomez, J.; Vassilopoulos, A.; Reisdorph, N.; Fritz, K.S.; Patel, M. Altered mitochondrial acetylation profiles in a kainic acid model of temporal lobe epilepsy. Free Radic. Biol. Med. 2018, 123, 116–124. [Google Scholar] [CrossRef]
- Chuang, Y.C.; Chen, S.D.; Jou, S.B.; Lin, T.K.; Chen, S.F.; Chen, N.C.; Hsu, C.Y. Sirtuin 1 regulates mitochondrial biogenesis and provides an endogenous neuroprotective mechanism against seizure-induced neuronal cell death in the hippocampus following status epilepticus. Int. J. Mol. Sci. 2019, 20, 3588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordone, M.P.; Salman, M.M.; Titus, H.E.; Amini, E.; Andersen, J.V.; Chakraborti, B.; Diuba, A.V.; Dubouskaya, T.G.; Ehrke, E.; Espindola de Freitas, A.; et al. The energetic brain–A review from students to students. J. Neurochem. 2019, 151, 139–165. [Google Scholar] [CrossRef] [PubMed]
- Cloix, J.F.; Hevor, T. Epilepsy, regulation of brain energy metabolism and neurotransmission. Curr. Med. Chem. 2009, 16, 841–853. [Google Scholar] [CrossRef] [Green Version]
- Ivannikov, M.V.; Sugimori, M.; Llinás, R.R. Synaptic vesicle exocytosis in hippocampal synaptosomes correlates directly with total mitochondrial volume. J. Mol. Neurosci. 2013, 49, 223–230. [Google Scholar] [CrossRef]
- Gray, N.E.; Alcazar, M.A.; Lak, P.; Wright, K.M.; Quinn, J.; Stevens, J.F.; Maier, C.S.; Soumyanath, A. Centella asiatica–Phytochemistry and mechanisms of neuroprotection and cognitive enhancement. Phytochem Rev. 2018, 17, 161–194. [Google Scholar] [CrossRef]
- Binder, D.K.; Steinhauser, C. Functional changes in astroglial cells in epilepsy. Glia 2006, 54, 358–368. [Google Scholar] [CrossRef]
- Verdugo, C.D.; Myren-Svelstad, S.; Aydin, E.; Hoeymissen, E.V.; Deneubourg, C.; Vanderhaeghe, S.; Vancraeynest, J.; Pelgrims, R.; Cosacak, M.I.; Muto, A.; et al. Glia-neuron interactions underlie state transitions to generalized seizures. Nat. Commun. 2019, 10, 3830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasurin, P.; Sriariyanun, M.; Phusantisampan, T. Review: The bioavailability activity of Centella asiatica, KMUTNB. Int. J. Appl. Sci. Technol. 2016, 9, 1–9. [Google Scholar] [CrossRef]
- Salman, M.M.; Marsh, G.; Kusters, I.; Delincé, M.; Di Caprio, G.; Upadhyayula, S.; de Nola, G.; Hunt, R.; Ohashi, K.G.; Gray, T.; et al. Design and validation of a human brain endothelial microvessel-on-a-chip open microfluidic model enabling advanced optical imaging. Front. Bioeng. Biotechnol. 2020, 8, 573775. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. | Protein Name | Accession Number | Mw/pI | Score (Coverage) | Match Fragment | Subcellular Location | Function |
|---|---|---|---|---|---|---|---|
| 1 | Tubulin alpha-1A chain | P68370 | 50.788/4.94 | 69 (35%) | 9 | Cytoplasm, cytoskeleton. | Tubulin is the major constituent of microtubules. It binds two moles of GTP, one at an exchangeable site on the beta chain and one at a non-exchangeable site on the alpha chain. |
| 2 | Dihydrolipoyl dehydrogenase | Q6P6R2 | 54.574/7.96 | 113 (27%) | 12 | Mitochondrion matrix | Lipoamide dehydrogenase is a component of the glycine cleavage system as well as an E3 component of three alpha-ketoacid dehydrogenase complexes (pyruvate-, alpha-ketoglutarate-, and branched- chain amino acid-dehydrogenase complex). |
| 3 | Glutamate dehydrogenase 1 | P10860 | 61.719/8.05 | 89 (28%) | 12 | Mitochondrion Endoplasmic reticulum | Mitochondrial glutamate dehydrogenase that converts L- glutamate into alpha-ketoglutarate. Plays a key role in glutamine anaplerosis by producing alpha-ketoglutarate, an important intermediate in the tricarboxylic acid cycle (By similarity). May be involved in learning and memory reactions by increasing the turnover of the excitatory neurotransmitter glutamate (PubMed:9275181). |
| 4 | Aconitate hydratase | Q9ER34 | 86.121/7.87 | 159 (29%) | 16 | Mitochondrion | Catalyzes the isomerization of citrate to isocitrate via cis-aconitate. |
| 5 | Pyruvate dehydrogenase E1 component subunit beta | P49432 | 39.299/6.20 | 130 (46%) | 12 | Mitochondrion matrix | The pyruvate dehydrogenase complex catalyzes the overall conversion of pyruvate to acetyl-CoA and CO2, and thereby links the glycolytic pathway to the tricarboxylic cycle. |
| 6 | Alpha-enolase | P04764 | 47.44/6.16 | 223 (56%) | 22 | Cytoplasm. Cell membrane. | Glycolytic enzyme the catalyzes the conversion of 2- phosphoglycerate to phosphoenolpyruvate. In addition to glycolysis, involved in various processes such as growth control, hypoxia tolerance and allergic responses. May also function in the intravascular and pericellular fibrinolytic system due to its ability to serve as a receptor and activator of plasminogen on the cell surface of several cell-types such as leukocytes and neurons. |
| 7 | Tubulin beta-2A chain | P85108 | 50.274/4.78 | 290 (59%) | 29 | Cytoplasm, cytoskeleton | Tubulin is the major constituent of microtubules. It binds two moles of GTP, one at an exchangeable site on the beta chain and one at a non-exchangeable site on the alpha chain (By similarity). |
| 8 | Dihydropyrimidinase-related protein 2 | P47942 | 62.638/5.95 | 217 (45%) | 20 | Cytoplasm | Plays a role in neuronal development and polarity, as well as in axon growth and guidance, neuronal growth cone collapse and cell migration. Necessary for signaling by class 3 semaphorins and subsequent remodeling of the cytoskeleton. |
| 9 | ATP synthase subunit alpha | P15999 | 59.831/9.22 | 205 (45%) | 22 | Mitochondrion | Mitochondrial membrane ATP synthase (F1F0 ATP synthase or Complex V) produces ATP from ADP in the presence of a proton gradient across the membrane which is generated by electron transport complexes of the respiratory chain. |
| 10 | ATP synthase subunit beta | P10719 | 56.318/5.19 | 266 (64%) | 33 | Mitochondrion inner membrane | During catalysis, ATP synthesis in the catalytic domain of F1 is coupled via a rotary mechanism of the central stalk subunits to proton translocation. Subunits alpha and beta form the catalytic core in F1. |
| 11 | V-type proton ATPase subunit B | P62815 | 56.857/5.57 | 276 (61%) | 26 | Membrane | Non-catalytic subunit of the peripheral V1 complex of vacuolar ATPase. V-ATPase is responsible for acidifying a variety of intracellular compartments in eukaryotic cells. |
| 12 | Dihydrolipoyllysine-residue succinyltransferase component of 2-oxoglutarate dehydrogenase complex | Q01205 | 49.236/8.89 | 75 (20%) | 8 | Mitochondrion matrix | The 2-oxoglutarate dehydrogenase complex catalyzes the overall conversion of 2-oxoglutarate to succinyl-CoA and CO2 (By similarity). The 2-oxoglutarate dehydrogenase complex is mainly active in the mitochondrion. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, C.-W.; Lin, T.-Y.; Pan, T.-L.; Wang, P.-W.; Chiu, K.-M.; Lee, M.-Y.; Wang, S.-J. Asiatic Acid Prevents Cognitive Deficits by Inhibiting Calpain Activation and Preserving Synaptic and Mitochondrial Function in Rats with Kainic Acid-Induced Seizure. Biomedicines 2021, 9, 284. https://doi.org/10.3390/biomedicines9030284
Lu C-W, Lin T-Y, Pan T-L, Wang P-W, Chiu K-M, Lee M-Y, Wang S-J. Asiatic Acid Prevents Cognitive Deficits by Inhibiting Calpain Activation and Preserving Synaptic and Mitochondrial Function in Rats with Kainic Acid-Induced Seizure. Biomedicines. 2021; 9(3):284. https://doi.org/10.3390/biomedicines9030284
Chicago/Turabian StyleLu, Cheng-Wei, Tzu-Yu Lin, Tai-Long Pan, Pei-Wen Wang, Kuan-Ming Chiu, Ming-Yi Lee, and Su-Jane Wang. 2021. "Asiatic Acid Prevents Cognitive Deficits by Inhibiting Calpain Activation and Preserving Synaptic and Mitochondrial Function in Rats with Kainic Acid-Induced Seizure" Biomedicines 9, no. 3: 284. https://doi.org/10.3390/biomedicines9030284