1. Introduction
The military are particularly exposed to infectious diseases as a consequence of their lifestyle with close inter-individual contacts and operational activity [
1]. Even if schedules vary among the armed forces of different countries, vaccination against many diseases using simultaneous inoculations of multiple antigens has been a common practice for many decades. Studies on military personnel have markedly contributed to vaccine development [
2] and to global public health [
3]. However, relatively few studies have faced the possible reciprocal negative or positive interference of simultaneous vaccines in adults [
4]. Simultaneous inoculation of multiple antigens, including combined vaccines, is nowadays a routine practice in children, but the development of combined vaccines is generally preceded by long and accurate studies demonstrating safety and efficacy and aimed at preventing the reduction in immunogenicity as a consequence of antigens’ interference [
5,
6]. In fact, studies in children receiving multiple vaccines demonstrated the possibility of reduced antibody response caused by antigen interference [
7,
8,
9]. A negative interference of tetanus/diphtheria and pneumococcal CRM197-conjugate vaccine was suggested [
10]. On the contrary, a possible positive interference has also been recently reported in the US military [
4], suggesting that the study of adults undergoing multiple, simultaneous vaccinations represents a valuable model to test this issue.
In the 1990s, the epidemiological situation of infectious diseases in the Italian military population was characterized by the net increase in varicella, rubella, and measles, as documented by the comparison of the period 1991–1995 with the period 1976–1980 [
11]. On this basis, in 1998, the military health authorities decided to introduce the trivalent live measles/mumps/rubella (MMR) vaccination, later associated to varicella, in the compulsory schedule for all recruits, irrespective of the possible already established protection for natural immunization [
12]. The effectiveness of the trivalent MMR was promptly demonstrated by observing 95% of measles and rubella and 70% of mumps cases’ reduction as early as two years after the introduction of MMR [
13]. Moreover, even the mumps vaccine’s effectiveness would have probably been higher if in the effectiveness calculation, vaccine-induced mumps-like clinical cases caused by the not sufficiently attenuated mumps vaccine strain Urabe Am9, included in the vaccine used at that time, had not been considered [
14]. Although in the pivotal study by Amanna et al. [
15], the very long duration of antibodies induced by viral antigens was clearly calculated, and in addition, pre-licensure studies have indicated the persistence of antibodies induced by the MMR vaccine as lifelong, the matter has been poorly studied [
16]. Even the possible interference by other concomitantly administered vaccines has only rarely been investigated.
A progressive lowering of hepatitis A virus (HAV) circulation was observed in Italy and documented by studies on military population at approximately 10-year intervals [
17]. However, the HAV vaccination was added to the compulsory vaccine schedule for the Italian military in 1998 [
12] considering that HAV is the most frequent vaccine-preventable infection in travelers [
18] and military personnel operate in many international scenarios. Although the vaccine’s immunogenicity is high and the anti-HAV antibody persistence is generally considered long-lasting, the possible negative or positive interference exerted by other viral or bacterial vaccines has not yet been thoroughly investigated.
Up until the end of the last century, the polio vaccine used for Italian infants was the live trivalent Sabin’s vaccine (oral polio vaccine (OPV)), then replaced in 2003 by the Salk’s trivalent inactivated polio vaccine (IPV) [
19]. In the military, a booster of IPV was introduced in 1998 in the compulsory vaccine schedule of permanent staff for possible deployment abroad [
12]. The efficacy of the polio vaccine has been so widely demonstrated that the World Health Organization (WHO) has already certified the global eradication of polioviruses types 2 [
20] and 3 [
21]. The wild type 1 is still present only in Pakistan and Afghanistan, even though the number of cases in 2019 increased fourfold compared to 2018 [
22]. However, to our knowledge, the immunogenicity of an IPV booster administered to adults primed at infancy by OPV and the persistence of the induced antibodies have not yet been explored or calculated, respectively.
Finally, influenza vaccination is offered on a voluntary basis, mainly to Italian military personnel deployed abroad. However, although influenza control is considered a crucial issue by the military [
23] and civil–military collaboration on influenza dates back to at least the Second World War [
24], only a limited number of world countries have included influenza vaccination in the compulsory vaccination program of their military personnel [
25]. Nevertheless, the study of the immune response to this vaccination in the military allows to gain more insights on a vaccination model which is unique. In fact, the immunization is repeated yearly due to the annual variability of viral strains.
The aim of the current study, which is part of a larger survey on the safety [
26] and immunogenicity [
27,
28] of vaccinations in the Italian military, was therefore to analyze immunogenicity and evaluate whether the current vaccination schedule for the military is adequate and the possible reciprocal positive and negative interferences of inactivated and living viral vaccines in two groups of Italian military personnel. The first group was represented by students from military schools working in Italy, and the second group was composed of soldiers who worked abroad (Lebanon) for nine months and received additional vaccinations before departure.
4. Discussion
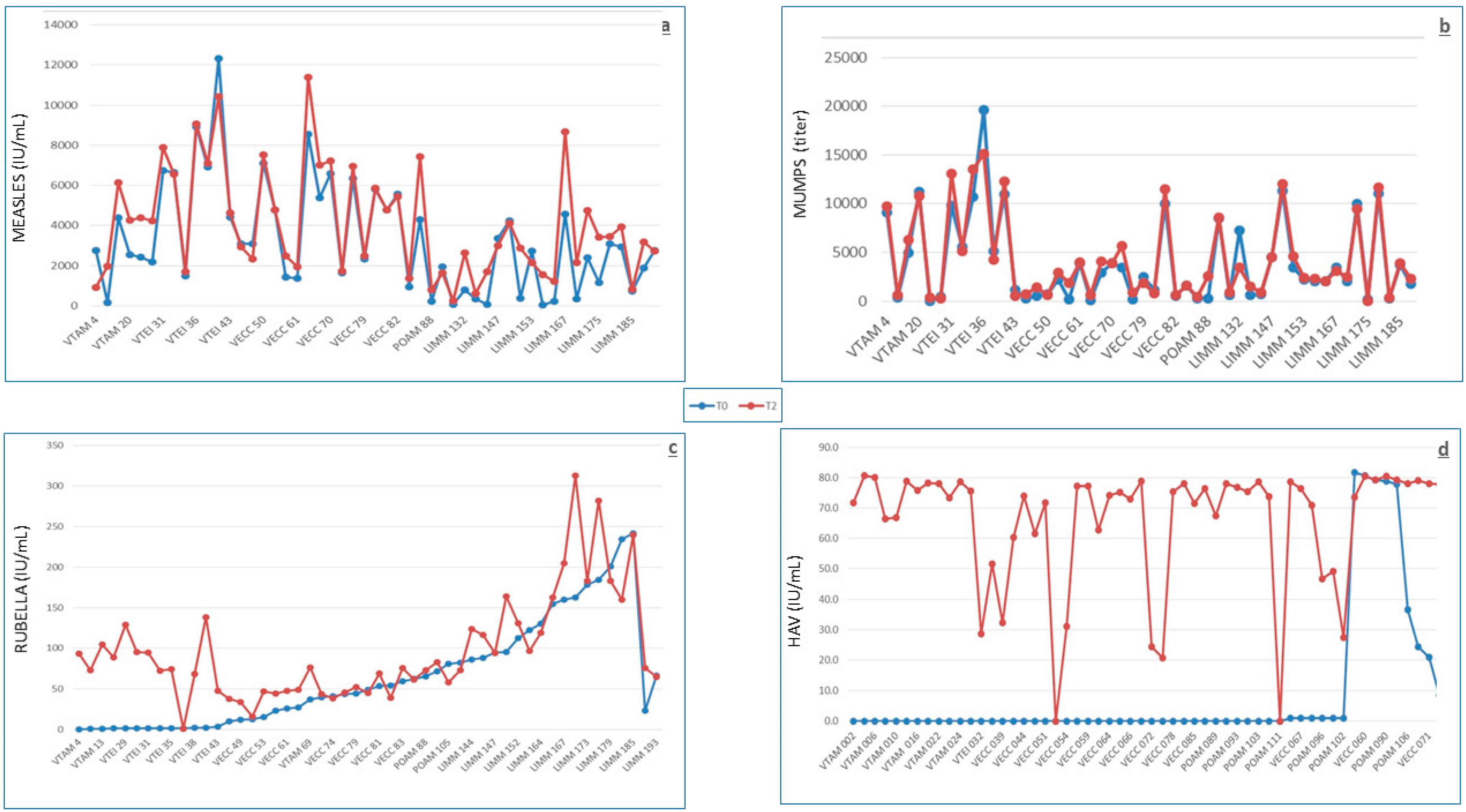
The analysis of antibody levels against MMR in 49 young Italian subjects showed a high rate of subjects with putative protective levels before vaccination. In fact, 82%, 82%, and 73.5% of the subjects already had antibodies above the putative threshold for protection against measles, mumps, and rubella, respectively, and 43% had protective antibody levels for all three viral antigens. Considering that the MMR vaccination was introduced for infants in Italy in 1999 [
40] and that the subjects described here had been vaccinated in 2013 at a mean age of over 21 years old, it can be argued that the large majority had antibody levels as a consequence of natural immunization. In fact, the circulation of measles and rubella viruses in Italy at the beginning of this century was high, and the relative infections were still very frequent, with an observed shift towards higher ages, albeit not always clinically evident [
41]. The long antibody persistence that we estimated on the basis of the calculated half-lives of 62, 50, and 58 years for measles, mumps, and rubella, respectively, is in line with the observations of Amanna et al. [
15]. Even if our estimates are markedly lower than those previously reported [
15], the expected lifelong persistence of these antibodies strongly indicates that booster doses might not be required for this vaccination, at least in healthy people. This suggestion is not in line with some reports showing a drop in circulating antibodies [
16,
42,
43]. Interestingly, in this line, there was an observation that a progressive decrease in antibody titers is not associated to a significant reduction in neutralizing antibodies when neutralizing antibodies were measured in parallel with specific antibodies by ELISA [
44]. The uselessness of booster doses is also suggested by the observation that the effect of a third booster is transient and is not required to maintain specific antibodies above the threshold of putative protection for a longer time [
44]. Long-lasting specific antibodies may also be involved in the rareness of post-vaccine adverse events. In addition to the general safety of the MMR vaccine, preexisting antibodies at putative protective titers observed in many subjects probably act as live vaccine-inactivating agents, reducing their capacity to interact with target cells. However, some doubts arise when considering the inverse relationship between baseline antibody levels and immune response and, even more so, the significant reduction in antibody levels in one third and one fourth of the subjects with putative protective antibody levels at baseline following measles and rubella vaccinations, respectively. Such doubts might derive from a vaccination strategy indiscriminately applied and not based on previous antibody level screening, which is a procedure that has also been considered for cost effectiveness reasons [
45]. This study was designed without control groups receiving single vaccinations, which would have enabled the analysis of vaccine reciprocal interference; however, the excellent immune response observed allows to infer a lack of negative interference by simultaneously administered vaccines, as already reported in the literature [
46].
The HAV baseline seropositivity, generally induced by natural immunization as the vaccination is not included in the Italian infant vaccination schedule, showed a net increase when compared with the military population analyzed over 10 years before [
17], being almost threefold higher (14% vs 5.3%). It is difficult to identify the possible reason(s) for such an increase considering that in Italy, a progressively decreasing epidemiological trend for HAV has been observed in recent decades and following the introduction, in 2012, of the HAV vaccine. In the Italian general population, the rate of HAV infection in 2012 was 0.8 per 100,000 [
47]. According to the Italian surveillance system for hepatitis, the main risk factors for HAV infection have been traditionally represented by consumption of raw shellfish, travel to endemic areas, and male homosexuality [
48]. Since 2013–2014, a new risk factor, represented by the consumption of contaminated mixed frozen berries, appeared and was responsible for a large outbreak in different Italian regions and European countries [
49]. The first cause of increased occurrence may be represented by the high prevalence of military personnel (nearly 50% of those vaccinated for HAV) coming from Campania and Puglia, the two Italian regions where the habit of raw seafood consumption is still traditionally present [
50]. However, the distribution of these individuals is not significantly different between the two groups, one with positive and the other with negative baseline antibody levels. As a consequence, this cause was ruled out, as was the consumption of contaminated frozen berries, considering that these subjects were studied one year before the outbreak. Other foods which may be contaminated and transmit HAV infection are poorly washed fresh vegetables, grilled meat, and fresh cheese [
51], as recently thoroughly discussed in relation to the description of a family cluster of HAV infections, not associated with raw shellfish or berries [
52]. Nonetheless, in the Italian military, detailed information on these issues as well as on the prevalence of male homosexuality is lacking. The vaccine’s immunogenicity has been confirmed to be very high, as already described [
53]. No post-vaccine case of HAV infection in the Italian military has been described, despite the military representing professional travelers and HAV being recognized as the biggest threat for travelers [
19], thus allowing to confirm a very high vaccine-induced protection. A significantly better response was observed in subjects with low rather than high baseline antibody levels, as already observed for quadrivalent meningococcal polysaccharide [
27] and for tetanus/diphtheria [
28] vaccines. The calculated half-life of antibodies is in line with the direct measurement of antibody levels at 22 years post-vaccination [
54], which has shown a very high percentage of still-protected individuals, thus suggesting that a booster would be unnecessary, considering the protection induced by immune memory [
55]. Lack of interference by concurrent vaccinations may be inferred from the results of the logistic regression.
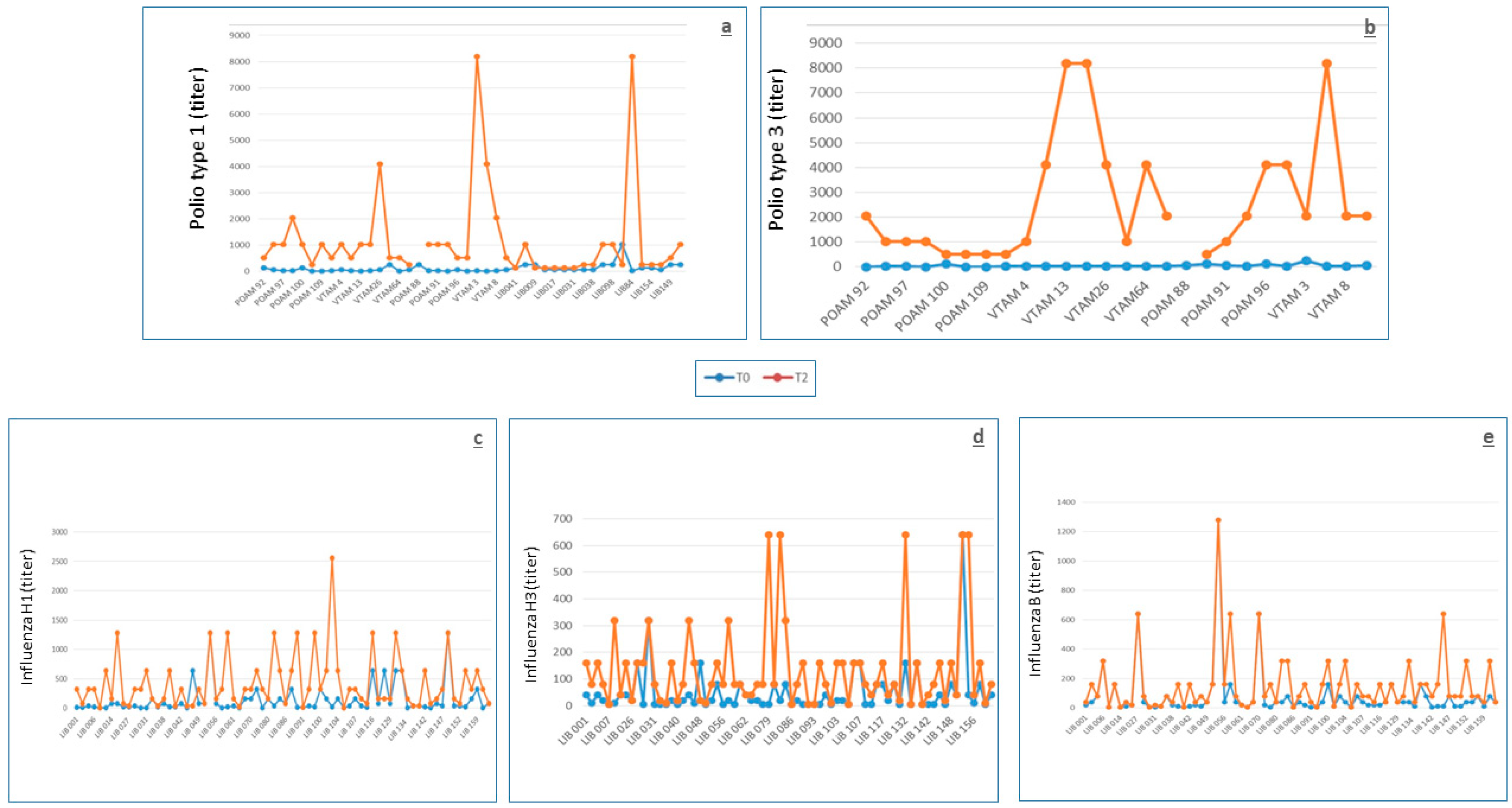
In this study, volunteers were vaccinated with the trivalent inactivated polio vaccine, but only the anti-type 1 and 3 antibodies were measured because type 2 has been officially considered eradicated by the WHO, which recommended the global withdrawal of the type 2 component of OPV in April 2016 [
20]. We could measure anti-type 3 antibodies, last isolated in 2012, since type 3 was declared eradicated by the WHO in October 2019 [
21]. Anti-polio antibodies types 1 and 3 were present in all tested individuals, except from only four individuals of group 1 who lacked protection against poliovirus type 3. These individuals showed specific antibodies at a titer of 1:4, which is considered just as protective [
31]. Considering that in Italy, the compulsory polio vaccination for infants was modified from OPV to a transient regime of OPV/IPV and then IPV only at the end of the last century [
19], we may argue that the baseline high level of protection could be a consequence of priming immunization at infancy with four doses of Sabin’s OPV, which was present in the Italian infant schedule until 1998. The level of baseline protection observed here is markedly higher than that found in an Italian population whose blood was collected in 2009 [
19], approximately in the same time period of the current study. In fact, by comparing the 26 subjects of group 1 of the current study, for whom the vaccination history is comparable to that of the 99 subjects, age range 15–43 years, of the cited study (only four doses of OPV at infancy), the prevalence of putatively protected individuals (neutralizing antibody titer ≥ 1:8) was 100% vs 78.8% for type 1 and 84.6% vs 38.4% for type 3, respectively (
p = 0.02292 for type 1;
p = 0.00006931 for type 3). The reasons for such discrepancy are not clear; however, lack of response and/or poor duration of antibodies to OPV at protective levels have already been described [
56] and confirmed in the Italian population [
57]. Moreover, several different factors, some of which are still unknown, may probably influence the immune response to the polio vaccine and the duration of anti-polio antibodies. This was shown by the relatively high rate of seronegative US military recruits observed in the 1970s [
58] as compared with a sample of Italian subjects analyzed in the same period [
59], who presented markedly lower rates of seronegativity, even though they were ranging in age from 6 months to 88 years. The response to the IPV booster was excellent and inversely associated with the baseline antibody levels (
Table 3), as generally observed with other vaccines and with the OPV [
56], but never, to the best of our knowledge, in the context of the OPV/IPV prime/booster. Despite the fact that the sequence IPV/OPV has been more extensively used, mainly for preventing vaccine-associated paralytic polio [
60], the model of prime-boosting OPV-IPV has also largely been used and has been studied at infancy [
61,
62,
63,
64] with either a simultaneous OPV-IPV administration or administration of OPV-IPV delivered separately in a short time interval. However, to the best of our knowledge, no study has been conducted on the serum immunogenicity of the IPV booster administered to adults primed at infancy by OPV. In the current experience, this model has proven able to provide good results, considering that all group 1 subjects seroconverted (at least fourfold titer increase), at 1 month, to both serotypes, and at 9 months, to type 3, and all subjects but one seroconverted to type 1. Therefore, the IPV booster introduced by the Italian military health authorities in the compulsory vaccination schedule for the military in 1998 has shown to be capable of efficiently boosting the immune response towards the two types of polioviruses tested, without adverse effects. In fact, despite the continuous decline of wild polioviruses in circulation globally, maintaining a high level of immunity towards the three types of polioviruses is pivotal for the military. This can be considered as such not only in order to avoid possible reintroduction of the wild viruses in Italy as a consequence of infections taken during international missions abroad but also to prevent possible vulnerabilities, which may be exploited in case of malevolent use of polioviruses as biological weapons [
57]. The lack of interference by the other concomitant vaccines may be inferred by the flat serum antibody titers observed in the seven subjects who had not received a recent polio booster. To the best of our knowledge, the post-vaccine serum antibody half-life has never been calculated; nevertheless, the long, even lifelong, persistence of anti-polio antibodies is well known [
65], and thus, the relatively low half-lives calculated here are probably a consequence of the low number of explored subjects.
The model of influenza vaccination is particularly intriguing, considering that the immunization is repeated yearly due to the annual drifts; thus, the immune response and effectiveness are markedly influenced by the immune history of subjects [
66]. In the current study, the immune response to the vaccine was good, considering that nearly 70% of subjects reached an antibody titer ≥ 1:40 towards all three H1, H3, and B antigens, which is generally considered putatively protective, even though the association between antibody titer and protection should only be considered as a guide [
67] and not as absolute. In fact, it may depend on many causes, including the history of previous immunizations, which may negatively interfere with antibody production in cases of tight similarity between the viral strains circulating in two subsequent influenza seasons [
66]. However, to underline the complexity of the influenza vaccination model, in cases of tight similarity between the viral strains circulating in subsequent influenza seasons, even an increase in baseline antibody levels has been described for a sort of cohort effect [
68]. Additionally, even a mismatch between vaccines and circulating viral strains may be responsible for reduced immune response [
66]. Conversely, the presence of the MF59 adjuvant provided a substantial contribution to the good antibody response [
69]. Nevertheless, no case of ILI has been described in vaccinated individuals, despite the circulation of influenza in Lebanon in 2014 (mainly the H3N2 strain) [
70]. It must be underlined that the influenza vaccine already showed good effectiveness in the military in 1944, inducing a 70% ILI reduction [
25]. Moreover, no mismatch between vaccines and circulating viral strains was noticed in 2014. An inverse association between baseline antibody values and the level of immune response was confirmed, as already observed in patients with rheumatoid arthritis [
68]. No interference by concomitant vaccines was observed, as documented by the results of the logistic regression.
The lack of association between HLA antigens and the immune response to the analyzed viral vaccines may perhaps be linked to the relatively low number of subjects explored, which prevented the possibility to study homogeneous subgroups consisting of enough subjects.

 ,
, 


{kind=link}
{kind=link}