Type 1 Diabetes Induces Hearing Loss: Functional and Histological Findings in An Akita Mouse Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Analysis of the Auditory Brainstem Response (ABR)
2.3. Metabolic Studies
2.4. Haematoxylin and Eosin (H&E) and Immunohistochemistry Staining
2.5. Electron Microscopy
2.6. SGN and Spiral Ligament (SL) Counts
2.7. Assessment of SV Thickness
2.8. CD31, Na+/K+-ATPase α1, Cleaved Caspase-3, and Intermediate Cell Intensity Quantitation
2.9. Statistical Analysis
2.10. Antibodies
3. Results
3.1. Akita Mice with Type 1 Diabetes Exhibit Hearing Loss
3.2. Type 1 Diabetes Induces Damage in the SGNs but Not the Organ of Corti (OC)
3.3. Type 1 Diabetes Induces Degeneration of the SV
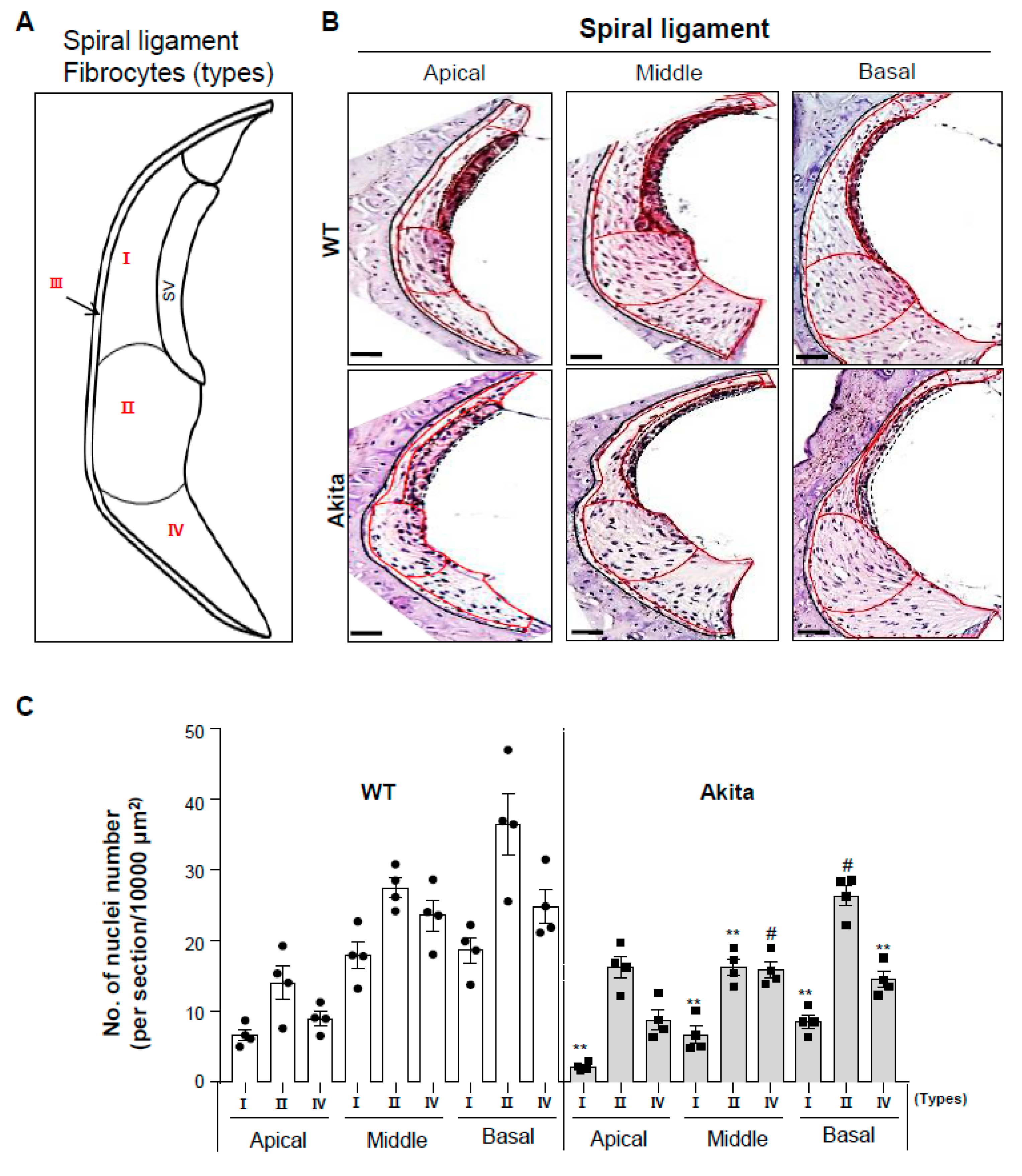
3.4. Type 1 Diabetes Induces a Decrease in Type I, II, and IV Fibrocytes in the Spiral Ligament
3.5. Type 1 Diabetes Induces a Decrease in Na+/K+-Atpase A1 Expression in the SL and SGNs
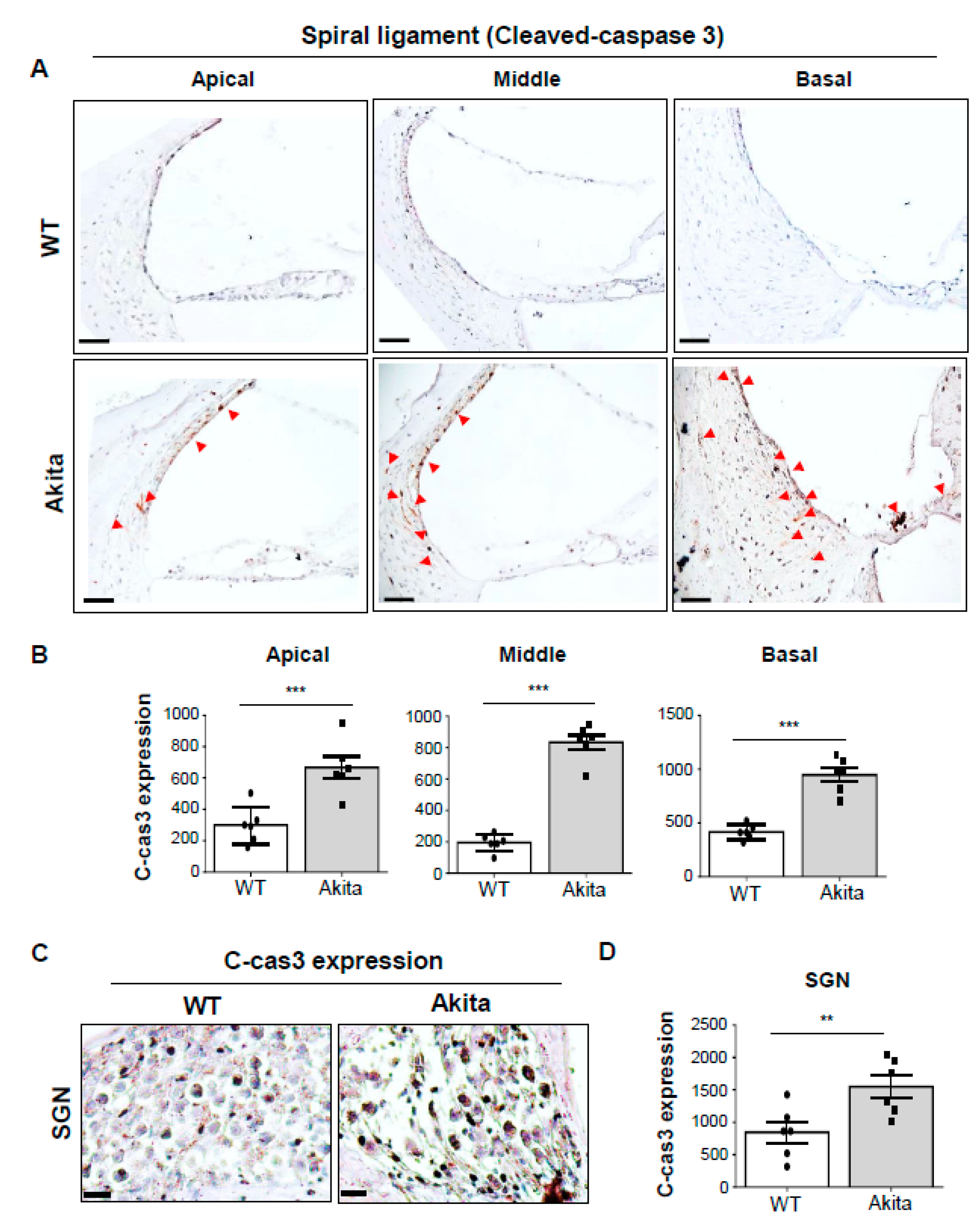
3.6. Activation of Caspase-3 is Associated with Degeneration of the SV and SL in the Type 1 Diabetes Cochlea
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABR | Auditory brainstem responses |
| SNHL | Sensorineural hearing loss |
| OC | Organ of Corti |
| SGN | Spiral ganglion neuron |
| LW | Lateral wall |
| SV | Stria vascularis |
| IHC | Inner hair cell |
| OHC | Outer hair cell |
| SC | Supporting cell; |
| SL | Spiral ligaments |
| Na+/K+-ATPase α1 | α1 subunit of Na+, K+-ATPase |
References
- Bagai, A.; Thavendiranathan, P.; Detsky, A.S. Does this patient have hearing impairment? JAMA 2006, 295, 416–428. [Google Scholar] [CrossRef] [PubMed]
- Stachler, R.J.; Chandrasekhar, S.S.; Archer, S.M.; Rosenfeld, R.M.; Schwartz, S.R.; Barrs, D.M.; Brown, S.R.; Fife, T.D.; Ford, P.; Ganiats, T.G.; et al. Clinical practice guideline: Sudden hearing loss. Otolaryngol. Head Neck Surg. 2012, 146, S1–S35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eizirik, D.L.; Pasquali, L.; Cnop, M. Pancreatic beta-cells in type 1 and type 2 diabetes mellitus: Different pathways to failure. Nat. Rev. Endocrinol. 2020, 16, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Barrett, E.J.; Liu, Z.; Khamaisi, M.; King, G.L.; Klein, R.; Klein, B.E.K.; Hughes, T.M.; Craft, S.; Freedman, B.I.; Bowden, D.W.; et al. Diabetic Microvascular Disease: An Endocrine Society Scientific Statement. J. Clin. Endocrinol. Metab. 2017, 102, 4343–4410. [Google Scholar] [CrossRef]
- Chung, W.S.; Lin, C.L.; Kao, C.H. Diabetes increases the risk of deep-vein thrombosis and pulmonary embolism. A population-based cohort study. Thromb. Haemost. 2015, 114, 812–818. [Google Scholar] [CrossRef]
- Bainbridge, K.E.; Hoffman, H.J.; Cowie, C.C. Diabetes and hearing impairment in the United States: Audiometric evidence from the National Health and Nutrition Examination Survey, 1999 to 2004. Ann. Intern. Med. 2008, 149, 1–10. [Google Scholar] [CrossRef]
- Kim, M.B.; Zhang, Y.; Chang, Y.; Ryu, S.; Choi, Y.; Kwon, M.J.; Moon, I.J.; Deal, J.A.; Lin, F.R.; Guallar, E.; et al. Diabetes mellitus and the incidence of hearing loss: A cohort study. Int. J. Epidemiol. 2017, 46, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Ashkezari, S.J.; Namiranian, N.; Rahmanian, M.; Atighechi, S.; Mohajeri-Tehrani, M.R.; Gholami, S. Is hearing impairment in diabetic patients correlated to other complications? J. Diabetes. Metab. Disord. 2018, 17, 173–179. [Google Scholar] [CrossRef]
- Fukushima, H.; Cureoglu, S.; Schachern, P.A.; Kusunoki, T.; Oktay, M.F.; Fukushima, N.; Paparella, M.M.; Harada, T. Cochlear changes in patients with type 1 diabetes mellitus. Otolaryngol. Head Neck Surg. 2005, 133, 100–106. [Google Scholar] [CrossRef]
- Gupta, S.; Eavey, R.D.; Wang, M.; Curhan, S.G.; Curhan, G.C. Type 2 diabetes and the risk of incident hearing loss. Diabetologia 2019, 62, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.L.; Raynor, E.; Prazma, J.; Buenting, J.E.; Pillsbury, H.C. Insulin-dependent diabetic microangiopathy in the inner ear. Laryngoscope 1995, 105, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, H.; Cureoglu, S.; Schachern, P.A.; Paparella, M.M.; Harada, T.; Oktay, M.F. Effects of type 2 diabetes mellitus on cochlear structure in humans. Arch. Otolaryngol. Head Neck Surg. 2006, 132, 934–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kariya, S.; Cureoglu, S.; Fukushima, H.; Morita, N.; Baylan, M.Y.; Maeda, Y.; Nishizaki, K.; Paparella, M.M. Comparing the cochlear spiral modiolar artery in type-1 and type-2 diabetes mellitus:a human temporal bone study. Acta Med. Okayama 2010, 64, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Wackym, P.A.; Linthicum, F.H., Jr. Diabetes mellitus and hearing loss: Clinical and histopathologic relationships. Am. J. Otol. 1986, 7, 176–182. [Google Scholar]
- Ishikawa, T.; Naito, Y.; Taniguchi, K. Hearing impairment in WBN/Kob rats with spontaneous diabetes mellitus. Diabetologia 1995, 38, 649–655. [Google Scholar] [CrossRef]
- Nakae, S.; Tachibana, M. The cochlea of the spontaneously diabetic mouse. II. Electron microscopic observations of non-obese diabetic mice. Arch. Otorhinolaryngol. 1986, 243, 313–316. [Google Scholar] [CrossRef]
- Tachibana, M.; Nakae, S. The cochlea of the spontaneously diabetic mouse. I. Electron microscopic observation of KK mice. Arch. Otorhinolaryngol. 1986, 243, 238–241. [Google Scholar] [CrossRef]
- Rust, K.R.; Prazma, J.; Triana, R.J.; Michaelis, O.E.t.; Pillsbury, H.C. Inner ear damage secondary to diabetes mellitus. II. Changes in aging SHR/N-cp rats. Arch. Otolaryngol. Head Neck Surg. 1992, 118, 397–400. [Google Scholar] [CrossRef]
- Raynor, E.M.; Carrasco, V.N.; Prazma, J.; Pillsbury, H.C. An assessment of cochlear hair-cell loss in insulin-dependent diabetes mellitus diabetic and noise-exposed rats. Arch. Otolaryngol. Head Neck Surg. 1995, 121, 452–456. [Google Scholar] [CrossRef]
- Geering, K. Functional roles of Na,K-ATPase subunits. Curr. Opin. Nephrol. Hypertens. 2008, 17, 526–532. [Google Scholar] [CrossRef]
- Skou, J.C. The identification of the sodium pump. Biosci. Rep. 2004, 24, 436–451. [Google Scholar] [CrossRef]
- Kerr, T.P.; Ross, M.D.; Ernst, S.A. Cellular localization of Na+,K+-ATPase in the mammalian cochlear duct: Significance for cochlear fluid balance. Am. J. Otolaryngol. 1982, 3, 332–338. [Google Scholar] [CrossRef]
- Gratton, M.A.; Smyth, B.J.; Schulte, B.A.; Vincent, D.A., Jr. Na,K-ATPase activity decreases in the cochlear lateral wall of quiet-aged gerbils. Hear. Res. 1995, 83, 43–50. [Google Scholar] [CrossRef]
- Choung, Y.H.; Kim, S.W.; Tian, C.; Min, J.Y.; Lee, H.K.; Park, S.N.; Lee, J.B.; Park, K. Korean red ginseng prevents gentamicin-induced hearing loss in rats. Laryngoscope 2011, 121, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, M.; Kayo, T.; Ikeda, T.; Koizumi, A. A novel locus, Mody4, distal to D7Mit189 on chromosome 7 determines early-onset NIDDM in nonobese C57BL/6 (Akita) mutant mice. Diabetes 1997, 46, 887–894. [Google Scholar] [CrossRef]
- Fujita, T.; Yamashita, D.; Katsunuma, S.; Hasegawa, S.; Tanimoto, H.; Nibu, K. Increased inner ear susceptibility to noise injury in mice with streptozotocin-induced diabetes. Diabetes 2012, 61, 2980–2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spicer, S.S.; Schulte, B.A. Differentiation of inner ear fibrocytes according to their ion transport related activity. Hear. Res. 1991, 56, 53–64. [Google Scholar] [CrossRef]
- Spicer, S.S.; Schulte, B.A. The fine structure of spiral ligament cells relates to ion return to the stria and varies with place-frequency. Hear. Res. 1996, 100, 80–100. [Google Scholar] [CrossRef]
- Yakovlev, A.G.; Faden, A.I. Caspase-dependent apoptotic pathways in CNS injury. Mol. Neurobiol. 2001, 24, 131–144. [Google Scholar] [CrossRef]
- Nicotera, T.M.; Hu, B.H.; Henderson, D. The caspase pathway in noise-induced apoptosis of the chinchilla cochlea. J. Assoc. Res. Otolaryngol. 2003, 4, 466–477. [Google Scholar] [CrossRef] [Green Version]
- Tadros, S.F.; D’Souza, M.; Zhu, X.; Frisina, R.D. Apoptosis-related genes change their expression with age and hearing loss in the mouse cochlea. Apoptosis 2008, 13, 1303–1321. [Google Scholar] [CrossRef] [Green Version]
- Haryuna, T.S.; Purba, A.H.; Farhat, F.; Alviandi, W. The Antiapoptotic Effect of Curcumin in the Fibroblast of the Cochlea in an Ototoxic Rat Model. Iran. J. Otorhinolaryngol. 2018, 30, 247–253. [Google Scholar] [PubMed]
- Makishima, K.; Tanaka, K. Pathological changes of the inner ear and central auditory pathway in diabetics. Ann. Otol. Rhinol. Laryngol. 1971, 80, 218–228. [Google Scholar] [CrossRef]
- Kovar, M. The inner ear in diabetes mellitus. ORL J. Otorhinolaryngol. Relat. Spec. 1973, 35, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, J.P.; Biurrun, O.; Lorente, J.; Conget, J.I.; de Espana, R.; Esmatjes, E.; Gomis, R. Auditory function in young patients with type 1 diabetes mellitus. Diabetes Res. Clin. Pract. 1991, 11, 17–22. [Google Scholar] [CrossRef]
- Oliveira, C.A.; Kruse, C.; Juhn, S.K. Cochlear vascular changes in streptozotocin diabetes in chinchillas. Trans. Sect. Otolaryngol. Am. Acad. Ophthalmol. Otolaryngol. 1977, 84, 443–451. [Google Scholar]
- Rubini, R.; Biasiolo, F.; Fogarolo, F.; Magnavita, V.; Martini, A.; Fiori, M.G. Brainstem auditory evoked potentials in rats with streptozotocin-induced diabetes. Diabetes Res. Clin. Pract. 1992, 16, 19–25. [Google Scholar] [CrossRef]
- Vasilyeva, O.N.; Frisina, S.T.; Zhu, X.; Walton, J.P.; Frisina, R.D. Interactions of hearing loss and diabetes mellitus in the middle age CBA/CaJ mouse model of presbycusis. Hear. Res. 2009, 249, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Teng, Z.P.; Tian, R.; Xing, F.L.; Tang, H.; Xu, J.J.; Zhang, B.W.; Qi, J.W. An association of type 1 diabetes mellitus with auditory dysfunction: A systematic review and meta-analysis. Laryngoscope 2017, 127, 1689–1697. [Google Scholar] [CrossRef]
- Robertson, D. Effects of acoustic trauma on stereocilia structure and spiral ganglion cell tuning properties in the guinea pig cochlea. Hear. Res. 1982, 7, 55–74. [Google Scholar] [CrossRef]
- Liberman, M.C.; Dodds, L.W. Single-neuron labeling and chronic cochlear pathology. III. Stereocilia damage and alterations of threshold tuning curves. Hear. Res. 1984, 16, 55–74. [Google Scholar] [CrossRef]
- Puel, J.L.; Ruel, J.; Gervais d’Aldin, C.; Pujol, R. Excitotoxicity and repair of cochlear synapses after noise-trauma induced hearing loss. Neuroreport 1998, 9, 2109–2114. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Kim, K.R.; Chung, W.H.; Cho, Y.S.; Hong, S.H. Early sensorineural hearing loss in ob/ob mouse, an animal model of type 2 diabetes. Clin. Exp. Otorhinolaryngol. 2008, 1, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keithley, E.M.; Feldman, M.L. Spiral ganglion cell counts in an age-graded series of rat cochleas. J. Comp. Neurol. 1979, 188, 429–442. [Google Scholar] [CrossRef]
- White, J.A.; Burgess, B.J.; Hall, R.D.; Nadol, J.B. Pattern of degeneration of the spiral ganglion cell and its processes in the C57BL/6J mouse. Hear. Res. 2000, 141, 12–18. [Google Scholar] [CrossRef]
- Kujawa, S.G.; Liberman, M.C. Adding insult to injury: Cochlear nerve degeneration after “temporary” noise-induced hearing loss. J. Neurosci. 2009, 29, 14077–14085. [Google Scholar] [CrossRef] [Green Version]
- Raynor, E.; Robison, W.G.; Garrett, C.G.; McGuirt, W.T.; Pillsbury, H.C.; Prazma, J. Consumption of a high-galactose diet induces diabetic-like changes in the inner ear. Otolaryngol. Head Neck Surg. 1995, 113, 748–754. [Google Scholar] [CrossRef]
- Gao, Y.; Yechikov, S.; Vazquez, A.E.; Chen, D.; Nie, L. Impaired surface expression and conductance of the KCNQ4 channel lead to sensorineural hearing loss. J. Cell Mol. Med. 2013, 17, 889–900. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, H.B. The role of an inwardly rectifying K(+) channel (Kir4.1) in the inner ear and hearing loss. Neuroscience 2014, 265, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Wangemann, P. Supporting sensory transduction: Cochlear fluid homeostasis and the endocochlear potential. J. Physiol. 2006, 576, 11–21. [Google Scholar] [CrossRef]
- Kikuchi, T.; Kimura, R.S.; Paul, D.L.; Takasaka, T.; Adams, J.C. Gap junction systems in the mammalian cochlea. Brain Res. Brain Res. Rev. 2000, 32, 163–166. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.Y.; Kim, Y.J.; Gil, E.S.; Kim, H.; Jang, J.H.; Choung, Y.-H. Type 1 Diabetes Induces Hearing Loss: Functional and Histological Findings in An Akita Mouse Model. Biomedicines 2020, 8, 343. https://doi.org/10.3390/biomedicines8090343
Lee YY, Kim YJ, Gil ES, Kim H, Jang JH, Choung Y-H. Type 1 Diabetes Induces Hearing Loss: Functional and Histological Findings in An Akita Mouse Model. Biomedicines. 2020; 8(9):343. https://doi.org/10.3390/biomedicines8090343
Chicago/Turabian StyleLee, Yun Yeong, Yeon Ju Kim, Eun Sol Gil, Hantai Kim, Jeong Hun Jang, and Yun-Hoon Choung. 2020. "Type 1 Diabetes Induces Hearing Loss: Functional and Histological Findings in An Akita Mouse Model" Biomedicines 8, no. 9: 343. https://doi.org/10.3390/biomedicines8090343