Effects of Anchomanes difformis on Inflammation, Apoptosis, and Organ Toxicity in STZ-Induced Diabetic Cardiomyopathy


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Preparation
2.3. Ethical Approval
2.4. Animals
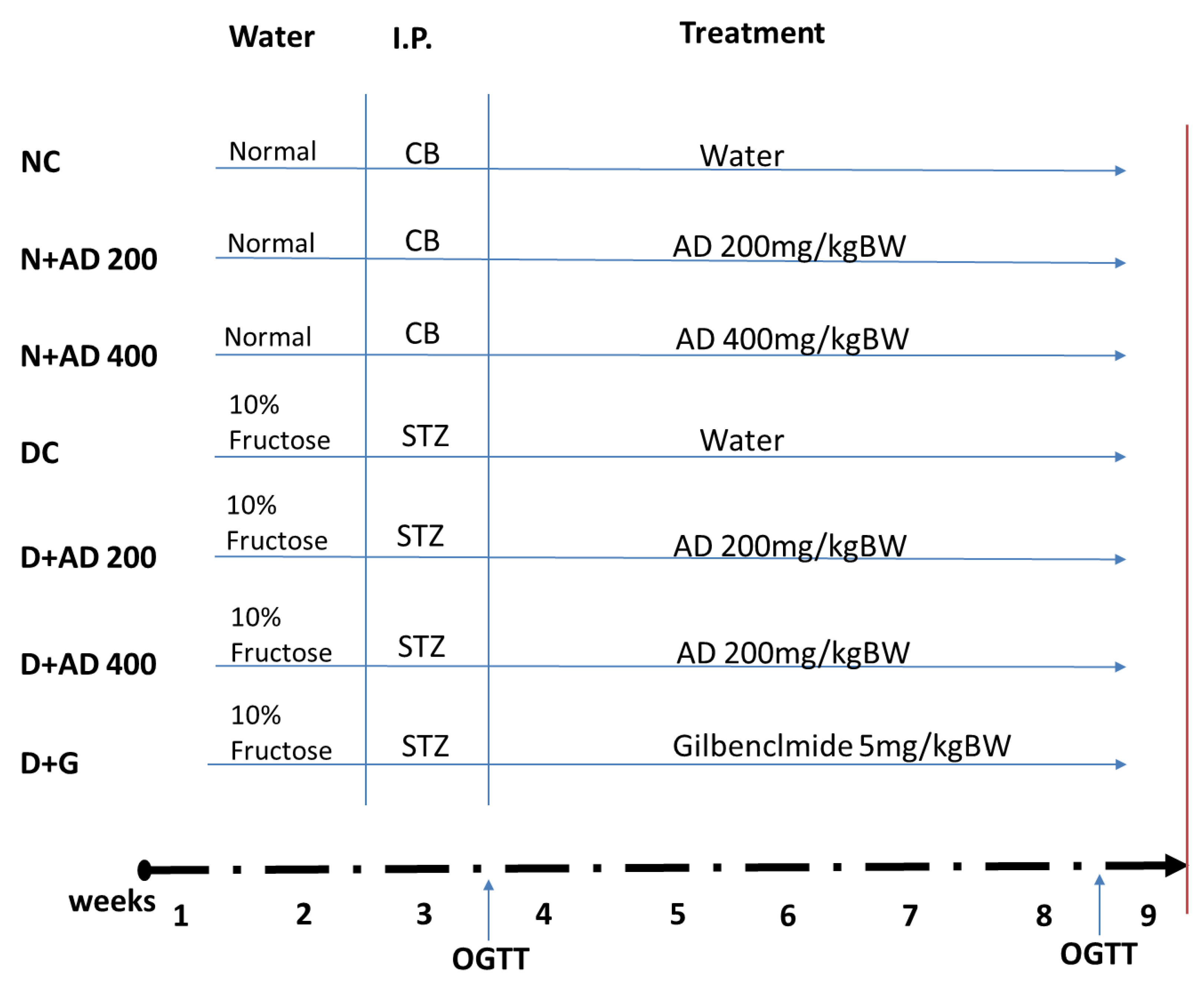
2.5. Experimental Design
2.6. Blood and Tissue Collection
2.7. Tissue Preparation
2.8. Determination of Organ Function and Toxicity Markers
2.9. Analysis of Antioxidant Status and Lipid Peroxidation Indices
2.10. Estimation of Pro-and Anti-Inflammatory Biomarkers
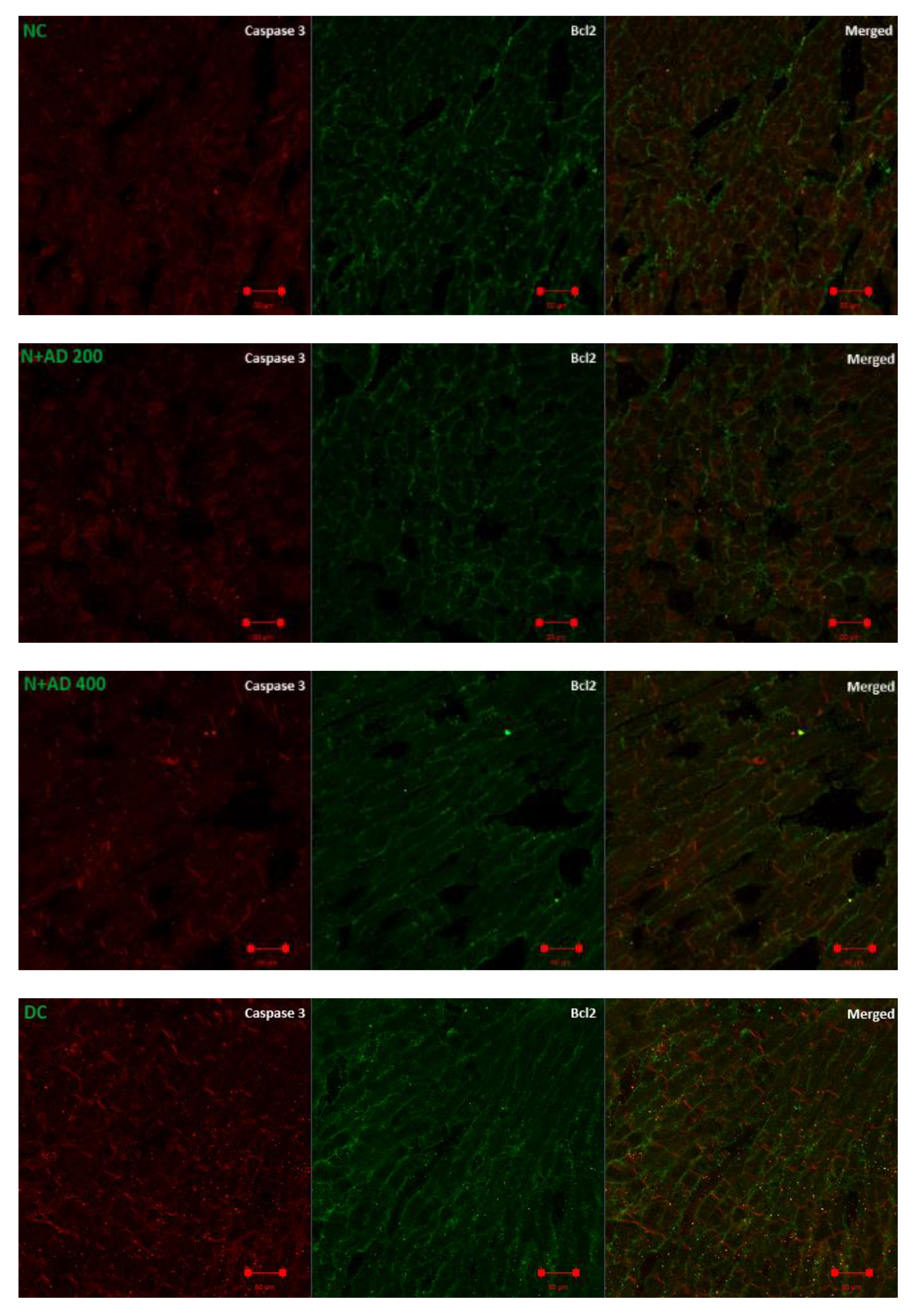
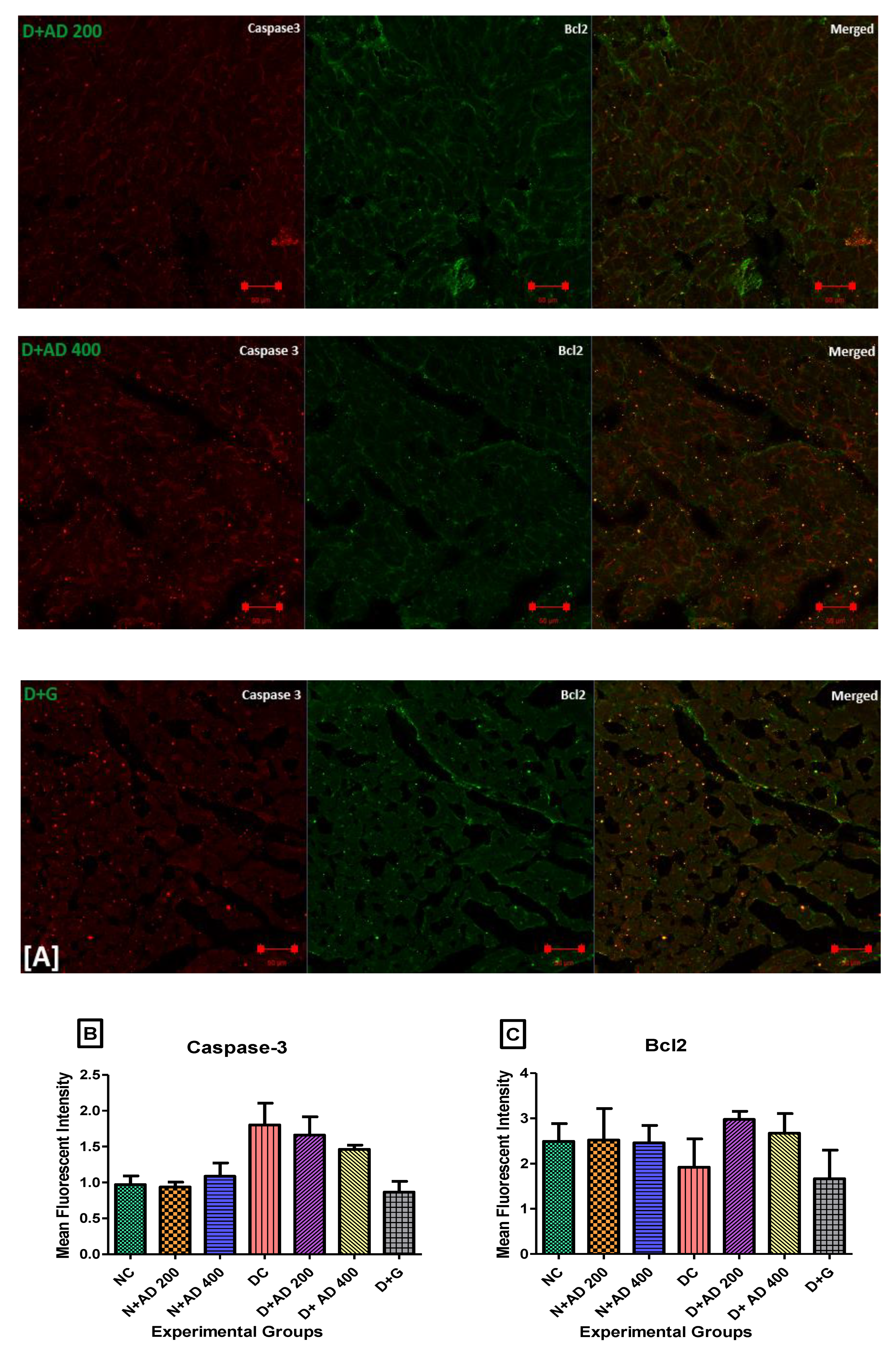
2.11. Evaluation of Apoptotic and Transcriptional Proteins Expression
2.12. Antibodies
2.13. Tissue Preparation and Staining
2.14. Imaging
2.15. Data Analysis
3. Results
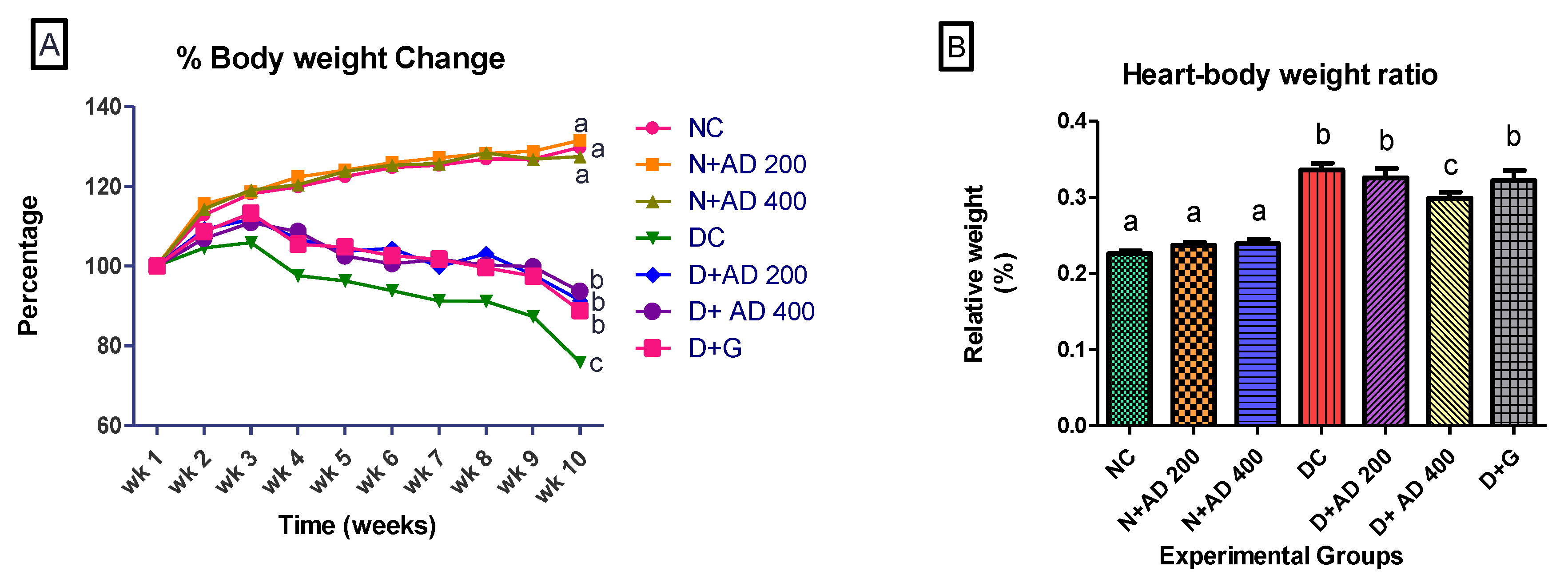
3.1. AD Reduced Weight Loss and Organ Toxicity in STZ-Induced Diabetes
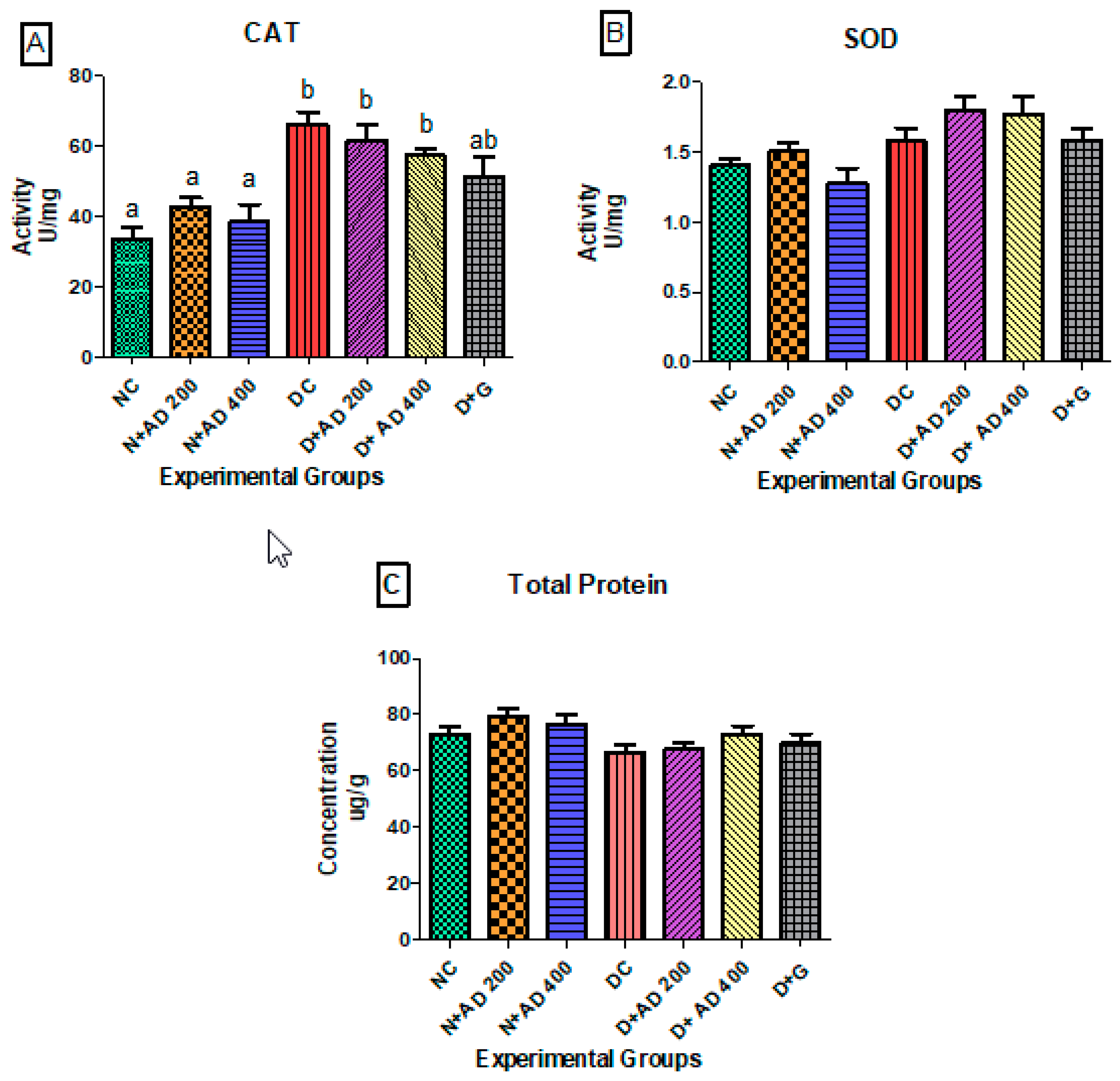
3.2. Effect of AD on Antioxidant Enzymes and Protein Synthesis in STZ Induced Diabetes
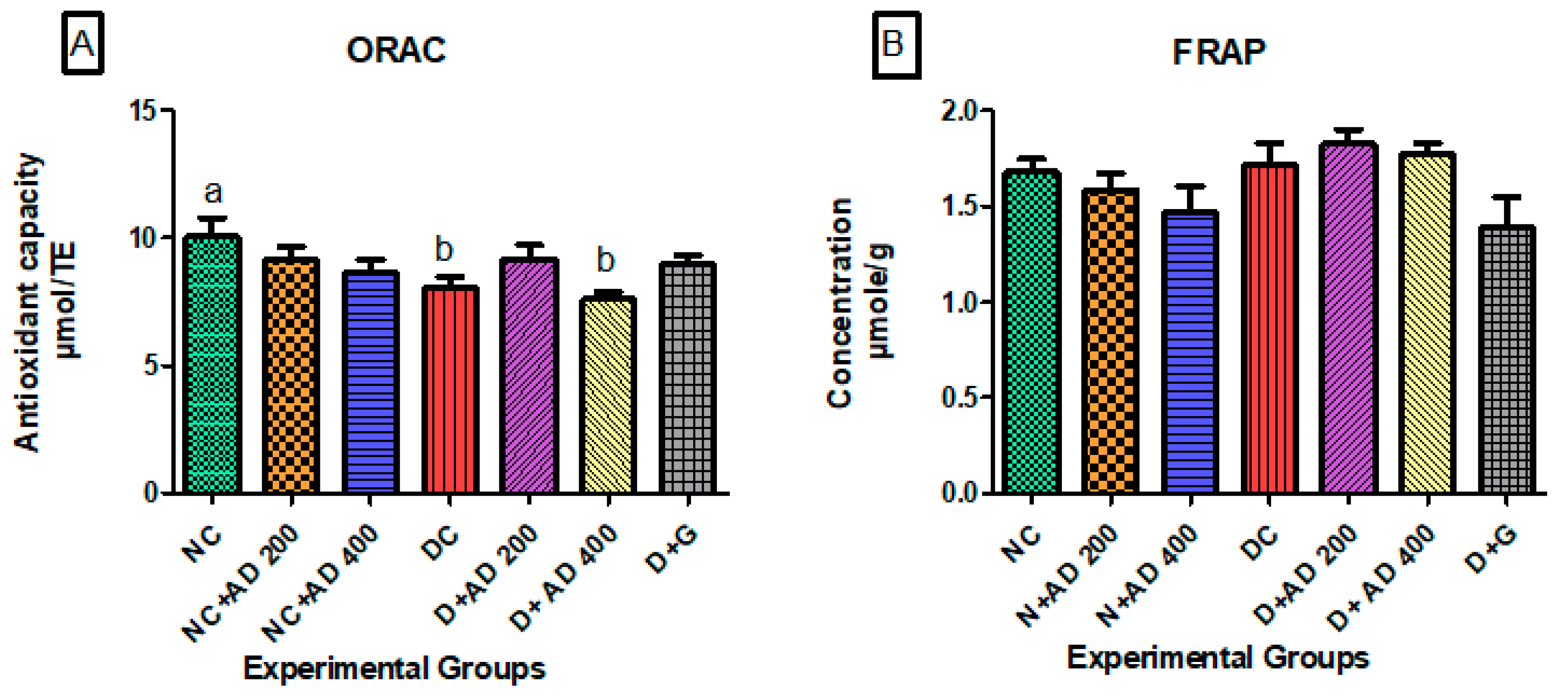
3.3. Effect of Treatment with AD on Antioxidant Indices in the Hearts of Normal and Diabetic Rats
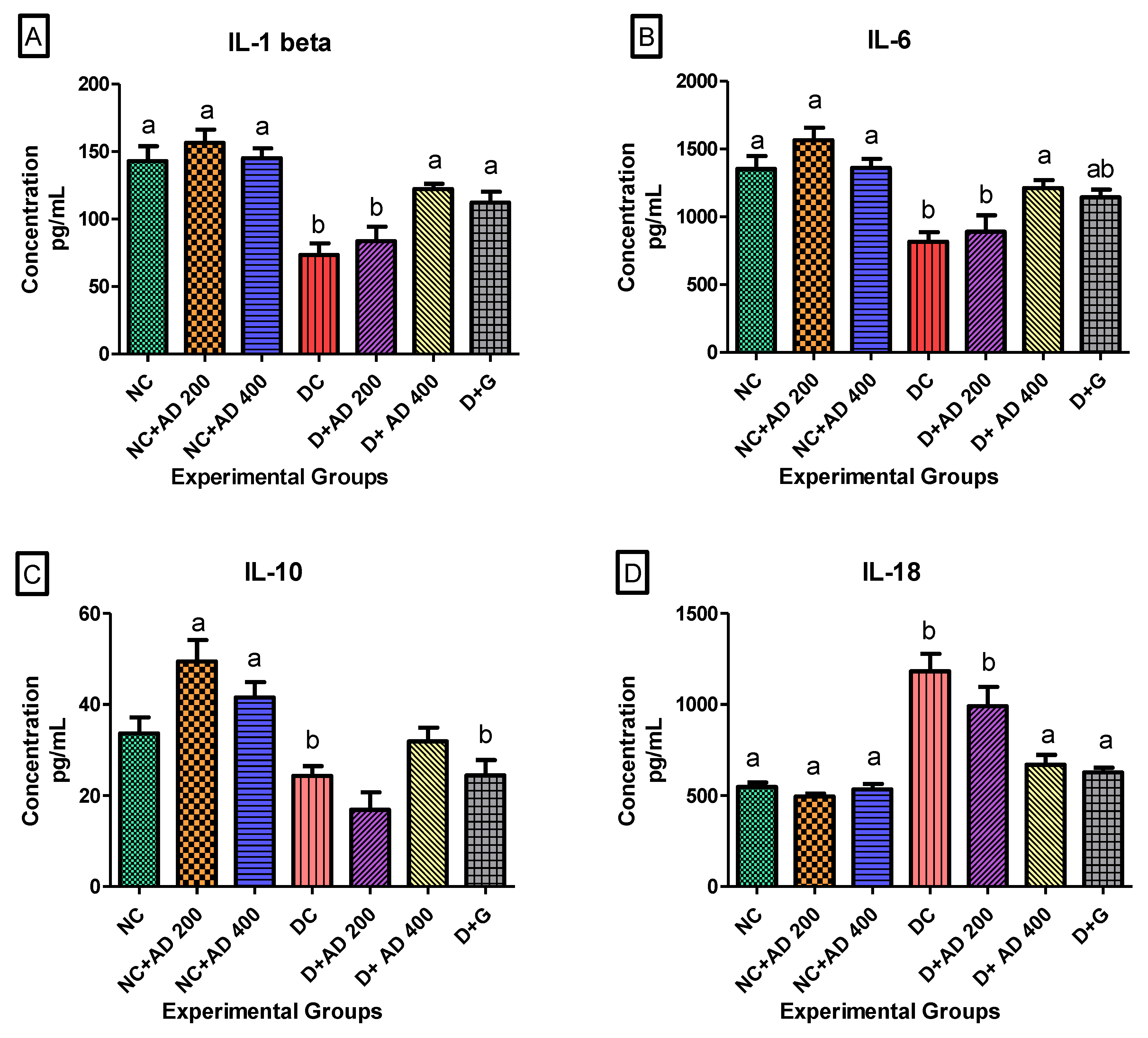
3.4. AD Modulated Hyperglycemia-Induced Immune Response in the Heart in T2D Model
3.5. The Effect of AD on Lipid Peroxidation and Heart Function Markers
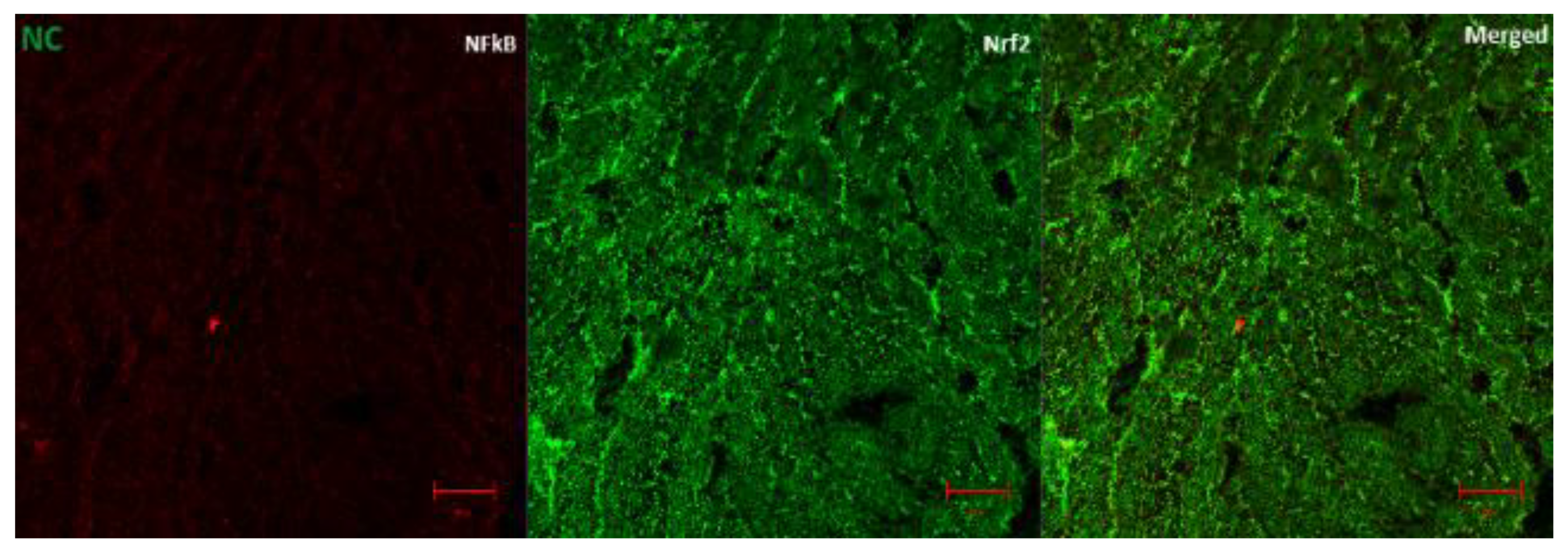
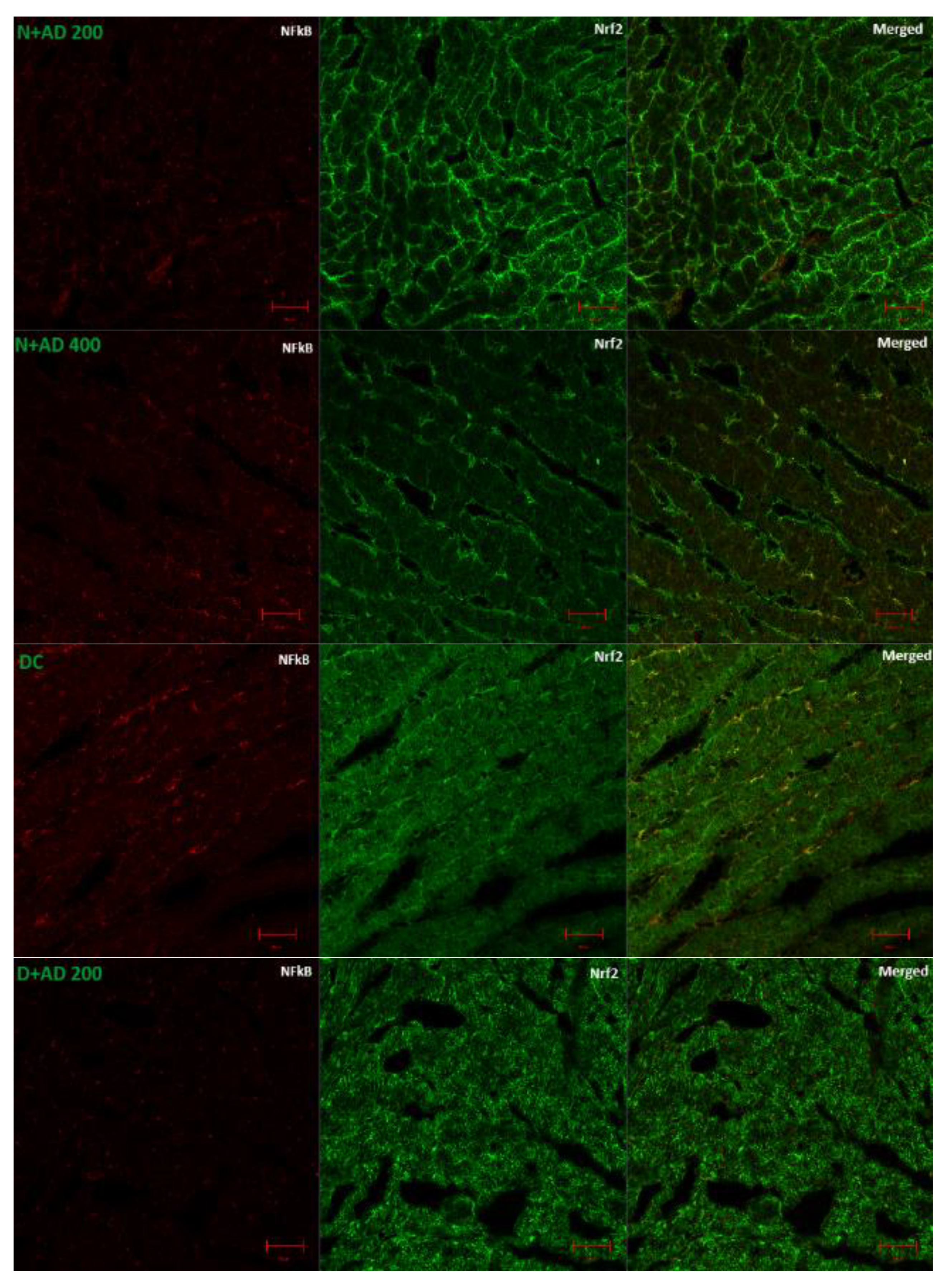
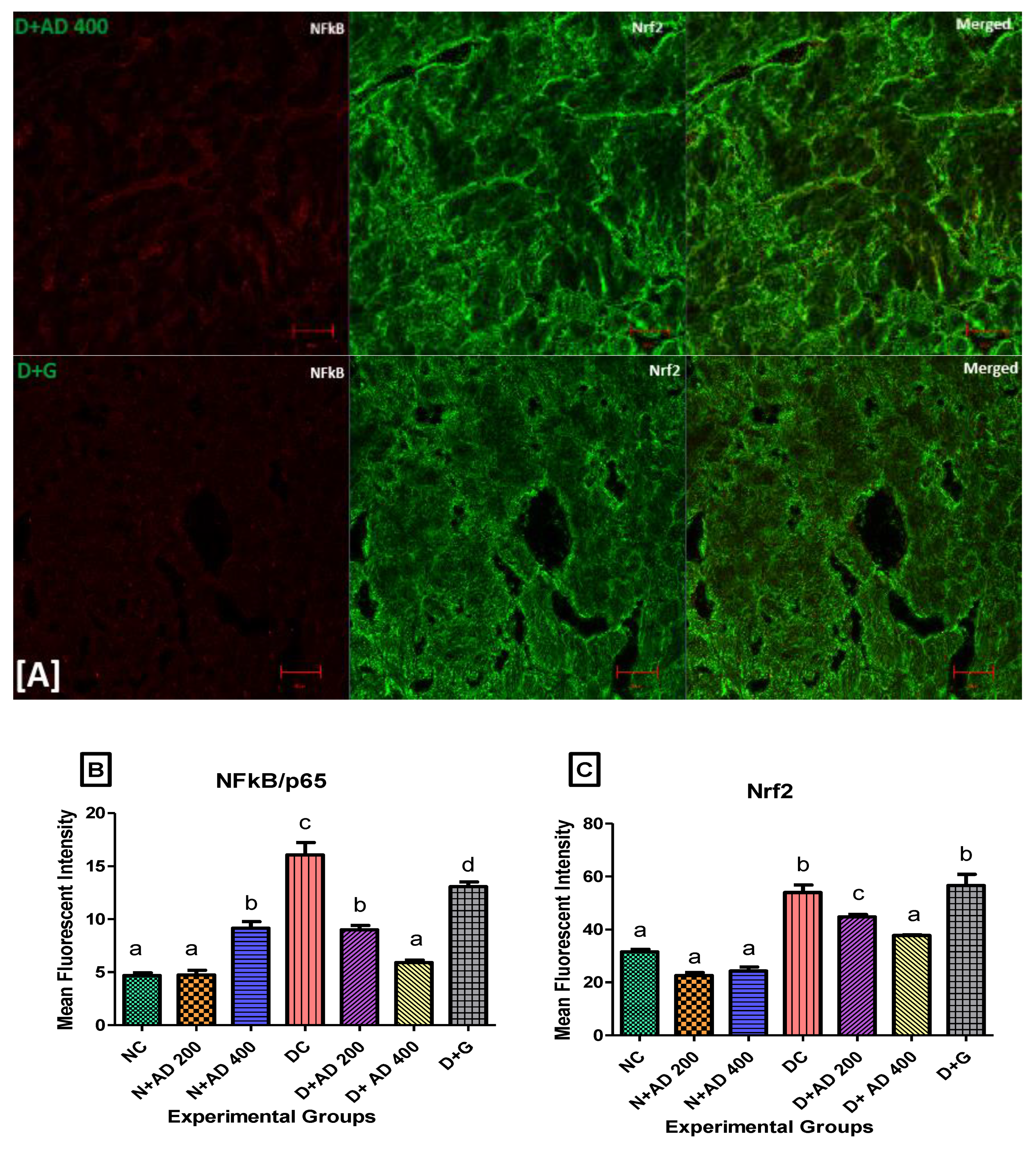
3.6. The Regulation of Transcription Factors by AD in T2D Model
3.7. Anti-Apoptotic Effect of AD on T2D Model
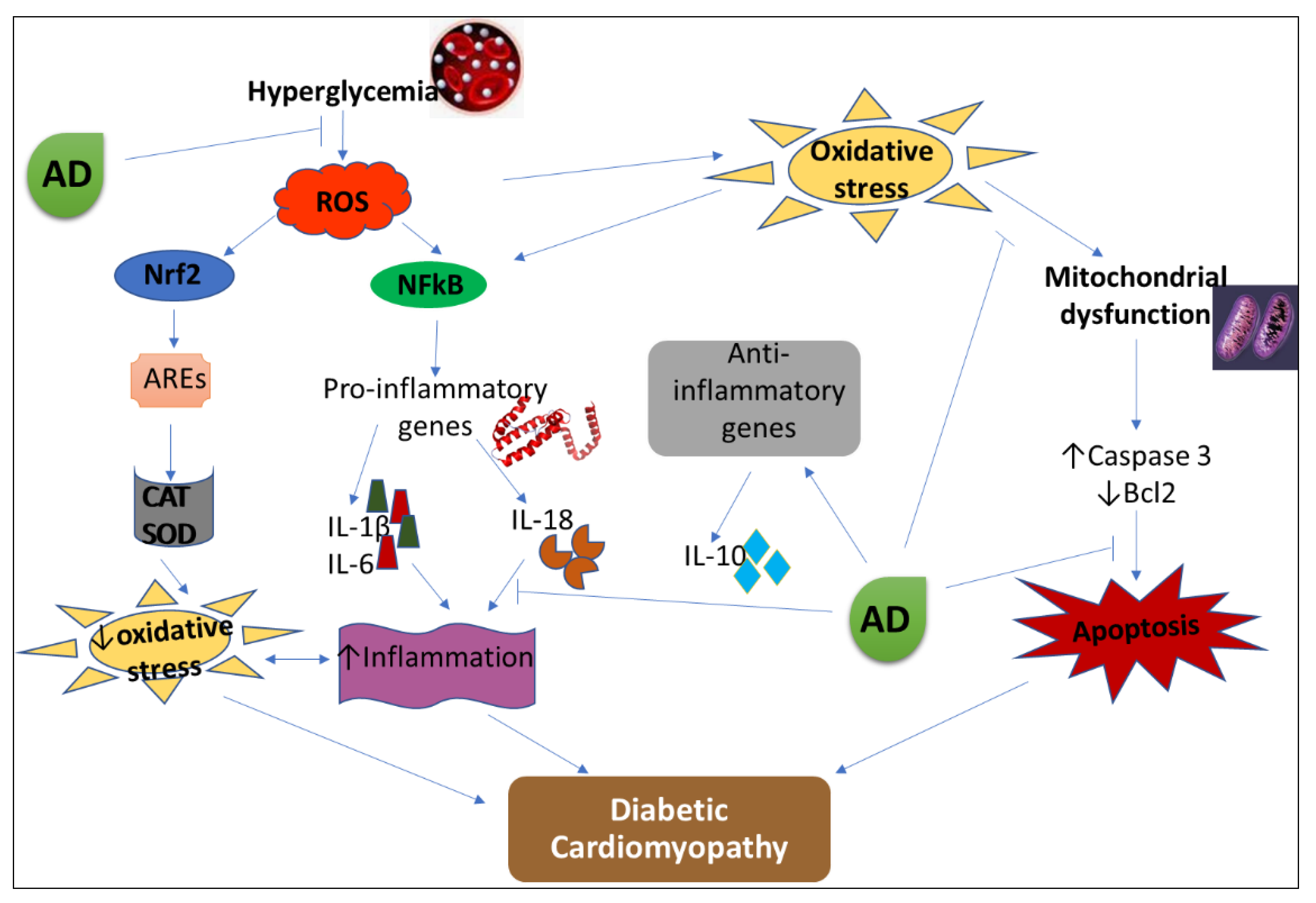
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Anchomanes difformis |
| ARE | Antioxidant response elements |
| Bcl2 | B-cell lymphoma 2 |
| CAT | Catalase |
| FRAP | Ferric reducing antioxidant power |
| H-FABP | Heart fatty acid binding protein |
| IL | Interleukin |
| KEAP-1 | Kelch-like ECH-associated protein 1 |
| MDA | Malondialdehyde |
| MCP-1 | Monocyte chemoattractant protein 1 |
| Nrf2 | Nuclear factor-erythroid 2-related factor 2 |
| NFkB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| ORAC | Oxygen radical absorbance capacity |
| PBS | Phosphate buffer saline |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| STZ | Streptozotocin |
| TBARS | Thiobarbituric acid reactive substances |
| TNF-α | Tumor necrosis factor alpha |
References
- Miranda-Díaz, A.G.; Pazarín-Villaseñor, L.; Yanowsky-Escatell, F.G.; Andrade-Sierra, J. Oxidative Stress in Diabetic Nephropathy with Early Chronic Kidney Disease. J. Diabetes Res. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Althunibat, O.Y.; al Hroob, A.M.; Abukhalil, M.H.; Germoush, M.O.; Bin-Jumah, M.; Mahmoud, A.M. Fisetin ameliorates oxidative stress, inflammation and apoptosis in diabetic cardiomyopathy. Life Sci. 2019, 221, 83–92. [Google Scholar] [CrossRef]
- Li, L.; Luo, W.; Qian, Y.; Zhu, W.; Qian, J.; Li, J.; Jin, Y.; Xu, X.; Liang, G. Luteolin protects against diabetic cardiomyopathy by inhibiting NF-κB-mediated inflammation and activating the Nrf2-mediated antioxidant responses. Phytomedicine 2019, 59, 152774. [Google Scholar] [CrossRef]
- Al Hroob, A.M.; Abukhalil, M.H.; Hussein, O.E.; Mahmoud, A.M. Pathophysiological mechanisms of diabetic cardiomyopathy and the therapeutic potential of epigallocatechin-3-gallate. Biomed. Pharmacother. 2019, 109, 2155–2172. [Google Scholar] [CrossRef]
- Paolillo, S.; Marsico, F.; Prastaro, M.; Renga, F.; Esposito, L.; de Martino, F.; di Napoli, P.; Esposito, I.; Ambrosio, A.; Ianniruberto, M.; et al. Diabetic Cardiomyopathy. Heart Fail. Clin. 2019, 15, 341–347. [Google Scholar] [CrossRef]
- Bajpai, A.; Tilley, D.G. The Role of Leukocytes in Diabetic Cardiomyopathy. Front. Physiol. 2018, 9, 1547. [Google Scholar] [CrossRef]
- Nunes, S.; Rolo, A.P.; Palmeira, C.M.; Reis, F. Diabetic Cardiomyopathy: Focus on Oxidative Stress, Mitochondrial Dysfunction and Inflammation. In Cardiomyopathies—Types Treat; InTech: Rijeka, Croatia, 2017. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Sun, S.-C. NF-κB in inflammation and renal diseases. Cell Biosci. 2015, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation, Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Mano, Y.; Anzai, T.; Kaneko, H.; Nagatomo, Y.; Nagai, T.; Anzai, A.; Maekawa, Y.; Takahashi, T.; Meguro, T.; Yoshikawa, T.; et al. Overexpression of Human C-Reactive Protein Exacerbates Left Ventricular Remodeling in Diabetic Cardiomyopathy. Circ. J. 2011, 75, 1717–1727. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.-Y.; Chen, H.-M.; Liang, J.-L.; Lin, Q.-X.; Tan, H.-H.; Fu, Y.-H.; Liu, X.-Y.; Shan, Z.-X.; Li, X.-H.; Yang, H.-Z.; et al. Hyperglycemic Myocardial Damage Is Mediated by Proinflammatory Cytokine: Macrophage Migration Inhibitory Factor. PLoS ONE 2011, 6, e16239. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Antrás, J.; Ioan, A.M.; Tuñón, J.; Egido, J.; Lorenzo, Ó. Activation of Toll-Like Receptors and Inflammasome Complexes in the Diabetic Cardiomyopathy-Associated Inflammation. Int. J. Endocrinol. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Goldin, A.; Beckman, J.A.; Schmidt, A.M.; Creager, M.A. Advanced Glycation End Products. Circulation 2006, 114, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Otsu, K. Inflammation and metabolic cardiomyopathy. Cardiovasc. Res. 2017, 113, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Kiraz, Y.; Adan, A.; Yandim, M.K.; Baran, Y. Major apoptotic mechanisms and genes involved in apoptosis. Tumor Biol. 2016, 37, 8471–8486. [Google Scholar] [CrossRef] [Green Version]
- Redza-dutordoir, M.; Averill-bates, D.A. Biochimica et Biophysica Acta Activation of apoptosis signalling pathways by reactive oxygen species. BBA Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Nunes, S.; Soares, E.; Pereira, F.; Reis, F. The role of inflammation in diabetic cardiomyopathy. Int. J. Infereron Cytokine Mediat. Res. 2012, 4, 59–73. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Hong, F.; Sekhar, K.R.; Freeman, M.L.; Liebler, D.C. Specific Patterns of Electrophile Adduction Trigger Keap1 Ubiquitination and Nrf2 Activation. J. Biol. Chem. 2005, 280, 31768–31775. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Yamamoto, M. Nrf2–Keap1 regulation of cellular defense mechanisms against electrophiles and reactive oxygen species. Adv. Enzyme Regul. 2006, 46, 113–140. [Google Scholar] [CrossRef]
- Baird, L.; Dinkova-Kostova, A.T. The cytoprotective role of the Keap1–Nrf2 pathway. Arch. Toxicol. 2011, 85, 241–272. [Google Scholar] [CrossRef]
- Merchant, A.A.; Singh, A.; Matsui, W.; Biswal, S. The redox-sensitive transcription factor Nrf2 regulates murine hematopoietic stem cell survival independently of ROS levels. Blood 2011, 118, 6572–6579. [Google Scholar] [CrossRef] [Green Version]
- Hennig, P.; Garstkiewicz, M.; Grossi, S.; di Filippo, M.; French, L.; Beer, H.-D. The Crosstalk between Nrf2 and Inflammasomes. Int. J. Mol. Sci. 2018, 19, 562. [Google Scholar] [CrossRef] [Green Version]
- Eleazu, C.; Eleazu, K.; Chukwuma, S.; Essien, U. Review of the mechanism of cell death resulting from streptozotocin challenge in experimental animals, its practical use and potential risk to humans. J. Diabetes Metab. Disord. 2013, 12, 60. [Google Scholar] [CrossRef] [Green Version]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2015, 70, 5–47. [Google Scholar] [CrossRef]
- Ma, M.; Mu, T. Anti-diabetic effects of soluble and insoluble dietary fibre from deoiled cumin in low-dose streptozotocin and high glucose-fat diet-induced type 2 diabetic rats. J. Funct. Foods 2016, 25, 186–196. [Google Scholar] [CrossRef]
- Barrière, D.A.; Noll, C.; Roussy, G.; Lizotte, F.; Kessai, A.; Kirby, K.; Belleville, K.; Beaudet, N.; Longpré, J.-M.; Carpentier, A.C.; et al. Combination of high-fat/high-fructose diet and low-dose streptozotocin to model long-term type-2 diabetes complications. Sci. Rep. 2018, 8, 424. [Google Scholar] [CrossRef]
- Wilson, R.D.; Islam, M.S. Fructose-fed streptozotocin-injected rat: An alternative model for type 2 diabetes. Pharmacol. Rep. 2012, 64, 129–139. [Google Scholar] [CrossRef]
- Udje, T.D.; Brooks, N.L.; Oguntibeju Oluwafemi, O. Medicinal Activities of Anchomanes difformis and its Potential in the Treatment of Diabetes Mellitus and Other Disease Conditions: A Review. In Bioactive Compounds of Medicinal Plants, 1st ed.; Goyal, M.R., Ayeleso, A.O., Eds.; Apple Academic Press: New York, NY, USA, 2018; pp. 219–235. [Google Scholar]
- Agyare, C.; Boakye, Y.D.; Apenteng, J.A.; Dapaah, S.O.; Appiah, T.; Adow, A. Antimicrobial and Anti-Inflammatory Properties of Anchomanes difformis (Bl.) Engl. and Colocasia esculenta (L.) Schott. Biochem. Pharmacol. Open Access. 2015, 5. [Google Scholar] [CrossRef]
- Adebayo, A.H.; John-Africa, L.B.; Agbafor, A.G.; Omotosho, O.E.; Mosaku, T.O. Anti-nociceptive and anti-inflammatory activities of extract of Anchomanes difformis in rats. Pak. J. Pharm. Sci. 2014, 27, 265–270. [Google Scholar]
- Aderonke, S.O.; Ezinwanne, A.J. Evaluation of the Anti Diabetic Activity of Ethanol Extract of Anchomanes Difformis (Araceae) Leaves in Albino Rats. Int. Res. J. Pharm. 2015, 6, 90–93. [Google Scholar] [CrossRef]
- Adeyemi, O.; Makinwa, T.T.; Uadia, R.N. Ethanol Extracts of Roots of Anchomanes difformis ENGL Roots as an Antihyperglycemic Agent in Diabetic Rats. Chem. J. 2015, 1, 68–73. [Google Scholar]
- Alabi, T.D.; Brooks, N.L.; Oguntibeju, O.O. Antioxidant Capacity, Phytochemical Analysis and Identification of Active Compounds in Anchomanes difformis. Nat. Prod. J. 2019, 9. [Google Scholar] [CrossRef]
- Najafian, M.; Jahromi, M.Z.; Nowroznejhad, M.J.; Khajeaian, P.; Kargar, M.M.; Sadeghi, M.; Arasteh, A. Phloridzin reduces blood glucose levels and improves lipids metabolism in streptozotocin-induced diabetic rats. Mol. Biol. Rep. 2012, 39, 5299–5306. [Google Scholar] [CrossRef]
- Josef, R.; Giribabu, N.; Karim, K.; Salleh, N. Quercetin ameliorates oxidative stress, inflammation and apoptosis in the heart of streptozotocin-nicotinamide-induced adult male diabetic rats. Biomed. Pharmacother. 2017, 86, 570–582. [Google Scholar]
- Ellerby, L.M.; Bredesen, D.E. Measurement of cellular oxidation, reactive oxygen species, and antioxidant enzymes during apoptosis. Methods Enzymol. 2000, 322, 413–421. [Google Scholar]
- Matsunami, T.; Sato, Y.; Sato, T.; Yukawa, M. Antioxidant status and lipid peroxidation in diabetic rats under hyperbaric oxygen exposure. Physiol. Res. 2010, 59, 97–104. [Google Scholar]
- Wasowicz, W.; Neve, J.; Peretz, A. Optimized steps in fluorometric determination of thiobarbituric acid-reactive substances in serum: Importance of extraction pH and influence of sample preservation and storage. Clin. Chem. 1993, 39, 2522–2526. [Google Scholar] [CrossRef]
- Prior, R.L.; Hoang, H.A.; Gu, L.; Wu, X.; Bacchiocca, M.; Howard, L.; Hampsch-Woodill, M.; Huang, D.; Ou, B.; Jacob, R. Assays for hydrophilic and lipophilic antioxidant capacity (oxygen radical absorbance capacity (ORACFL)) of plasma and other biological and food samples. J. Agric. Food Chem. 2003, 51, 3273–3279. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; pp. 15–27. [Google Scholar]
- Akhigbe, R.E. Discordant Results in Plant Toxicity Studies in Africa: Attempt of Standardization. In Toxicological Survey of African Medicinal Plants; Elsevier: Amsterdam, The Netherlands, 2014; pp. 53–61. [Google Scholar] [CrossRef]
- Delahanty, L.M. Weight loss in the prevention and treatment of diabetes. Prev. Med. 2017, 104, 120–123. [Google Scholar] [CrossRef]
- Hispard, F.; de Vaufleury, A.; Martin, H.; Devaux, S.; Cosson, R.P.; Scheifler, R.; Richert, L.; Berthelot, A.; Badot, P.-M. Effects of subchronic digestive exposure to organic or inorganic cadmium on biomarkers in rat tissues. Ecotoxicol. Environ. Saf. 2008, 70, 490–498. [Google Scholar] [CrossRef]
- Ige, S.F.; Akhigbe, R.E.; Edeogho, O.; Ajao, F.O.; Owolabi, O.Q.; Oyekunle, O.S.; Ajayi, A.F. Hepatoprotective activities of Allium cepa in cadmium-treated rats. Int. J. Pharm. Pharm. Sci. 2011, 3, 60–63. [Google Scholar]
- Khanra, R.; Dewanjee, S.; Dua, T.K.; Sahu, R.; Gangopadhyay, M.; DeFeo, V.; Zia-Ul-Haq, M. Abroma augusta L. (Malvaceae) leaf extract attenuates diabetes induced nephropathy and cardiomyopathy via inhibition of oxidative stress and inflammatory response. J. Transl. Med. 2015, 13, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mátyás, C.; Németh, B.T.; Oláh, A.; Török, M.; Ruppert, M.; Kellermayer, D.; Barta, B.A.; Szabó, G.; Kökény, G.; Horváth, E.M.; et al. Prevention of the development of heart failure with preserved ejection fraction by the phosphodiesterase-5A inhibitor vardenafil in rats with type 2 diabetes. Eur. J. Heart Fail. 2017, 19, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Addepalli, V.; Suryavanshi, S.V. Catechin attenuates diabetic autonomic neuropathy in streptozotocin induced diabetic rats. Biomed. Pharmacother. 2018, 108, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Al-Malki, A.L.; el Rabey, H.A. The Antidiabetic Effect of Low Doses of Moringa oleifera Lam. Seeds on Streptozotocin Induced Diabetes and Diabetic Nephropathy in Male Rats. Biomed. Res. Int. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, K.; Bernardo, B.C.; McMullen, J.R.; Ritchie, R.H. Diabetic cardiomyopathy: Mechanisms and new treatment strategies targeting antioxidant signaling pathways. Pharmacol. Ther. 2014, 142, 375–415. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Ichikawa, T.; Li, J.; Si, Q.; Yang, H.; Chen, X.; Goldblatt, C.S.; Meyer, C.J.; Li, X.; Cai, L.; et al. Diabetic downregulation of Nrf2 activity via ERK contributes to oxidative stress-induced insulin resistance in cardiac cells in vitro and in vivo. Diabetes 2011, 60, 625–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, T.; Sun, X.; Li, Y.; Chai, Q.; Wang, X.L.; Lee, H.C. Role of Nrf2 signaling in the regulation of vascular BK channel β1 subunit expression and BK channel function in high-fat diet–induced diabetic mice. Diabetes 2017, 66, 2681–2690. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Liu, S.; Miao, L.; Cai, L. Prevention of Diabetic Complications by Activation of Nrf2: Diabetic Cardiomyopathy and Nephropathy. Exp. Diabetes Res. 2012, 2012, 1–7. [Google Scholar] [CrossRef]
- Kobayashi, A.; Kang, M.-I.; Watai, Y.; Tong, K.I.; Shibata, T.; Uchida, K.; Yamamoto, M. Oxidative and Electrophilic Stresses Activate Nrf2 through Inhibition of Ubiquitination Activity of Keap1. Mol. Cell. Biol. 2006, 26, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Benipal, S.S.; Liu, T.; Knowlton, A. Repetitive ROS Inhibits Nrf2 Antioxidant Defense in Ischemic Heart Failure. FASEB J. 2017, 31, 1080-19. [Google Scholar]
- Dodson, M.; Castro-Portuguez, R.; Zhang, D.D. NRF2 plays a critical role in mitigating lipid peroxidation and ferroptosis. Redox Biol. 2019, 23, 101107. [Google Scholar] [CrossRef] [PubMed]
- Hasan, H.R.; Abdulsattar, A. Influence of diabetes disease on concentration of total protein, albumin and globulins in saliva and serum: A comparative study. Iraqi Natl. J. Chem. 2015, 15, 1–11. [Google Scholar]
- Mathy-Hartert, M.; Hogge, L.; Sanchez, C.; Deby-Dupont, G.; Crielaard, J.M.; Henrotin, Y. Interleukin-1β and interleukin-6 disturb the antioxidant enzyme system in bovine chondrocytes: A possible explanation for oxidative stress generation. Osteoarthr. Cartil. 2008, 16, 756–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez-Ruiz, M.C.; Quiroz, L.E.G.; Hernandez, E.; Bucio, L.; Souza, V.; Llorente, L.; Kershenobich, D. Cytokine response and oxidative stress produced by ethanol, acetaldehyde and endotoxin treatment in HepG2 cells. Isr. Med. Assoc. J. 2001, 3, 131–136. [Google Scholar]
- Del Giudice, M.; Gangestad, S.W. Rethinking IL-6 and CRP: Why they are more than inflammatory biomarkers, and why it matters. Brain. Behav. Immun. 2018, 70, 61–75. [Google Scholar] [CrossRef]
- Schett, G. Physiological effects of modulating the interleukin-6 axis. Rheumatology 2018, 57, ii43–ii50. [Google Scholar] [CrossRef] [Green Version]
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef]
- Nakamura, H.; Matoba, S.; Iwai-Kanai, E.; Kimata, M.; Hoshino, A.; Nakaoka, M.; Katamura, M.; Okawa, Y.; Ariyoshi, M.; Mita, Y.; et al. p53 Promotes Cardiac Dysfunction in Diabetic Mellitus Caused by Excessive Mitochondrial Respiration-Mediated Reactive Oxygen Species Generation and Lipid Accumulation. Circ. Heart Fail. 2012, 5, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Boudina, S.; Abel, E.D. Diabetic Cardiomyopathy Revisited. Circulation 2007, 115, 3213–3223. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Marker | Antibody | Host | Source | Dilution |
|---|---|---|---|---|---|
| 1. | Nrf2 | Anti-Nrf2 | Mouse | ABCAM, UK | 1:200 |
| 2. | NFkB/p65 | Anti-NFkB/p65 | Rabbit | ABCAM, UK | 1:250 |
| 3. | Bcl2 | Anti-Bcl2 | Mouse | BioLegend, SA | 1:125 |
| 4. | Caspase-3 | Anti-Caspase-3 | Rabbit | ABCAM, UK | 1:150 |
| 5. | Alexa Fluor 488 | Anti-mouse IgG H&L | Goat | ABCAM, UK | 1:400 |
| 6. | Alexa Fluor 594 | Anti-rabbit IgG H&L | Goat | ABCAM, UK | 1:400 |
| Experimental Groups | TNFα (pg/mL) | MCP-1 (pg/mL) |
|---|---|---|
| NC | 56.87 ± 3.68 | 288.8 ± 19.34 |
| N + AD 200 | 61.39 ± 2.42 | 272.1 ± 23.34 |
| N + AD 400 | 60.05 ± 3.38 | 287.4 ± 15.24 |
| DC | 50.40 ± 3.06 | 306.3 ± 15.03 |
| D + AD 200 | 48.50 ± 3.68 | 280.7 ± 12.25 |
| D + AD 400 | 49.72 ± 2.28 | 301.1 ± 23.00 |
| D + G | 51.52 ± 3.06 | 313.1 ± 27.05 |
| Experimental Groups | H-FABP (ng/mL) | TBARS (pmole/g) |
|---|---|---|
| NC | 0.216 ± 0.011 a | 2.226 ± 0.15 |
| N + AD 200 | 0.2784 ± 0.004 | 2.080 ± 0.09 |
| N + AD 400 | 0.2896 ± 0.003 | 2.032 ± 0.16 |
| DC | 0.2955 ± 0.002 b | 1.809 ± 0.10 |
| D + AD 200 | 0.2926 ± 0.006 b | 2.068 ± 0.11 |
| D + AD 400 | 0.2918 ± 0.005 b | 2.371 ± 0.10 |
| D + G | 0.3121 ± 0.003 b | 1.867 ± 0.12 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alabi, T.D.; Chegou, N.N.; Brooks, N.L.; Oguntibeju, O.O. Effects of Anchomanes difformis on Inflammation, Apoptosis, and Organ Toxicity in STZ-Induced Diabetic Cardiomyopathy. Biomedicines 2020, 8, 29. https://doi.org/10.3390/biomedicines8020029
Alabi TD, Chegou NN, Brooks NL, Oguntibeju OO. Effects of Anchomanes difformis on Inflammation, Apoptosis, and Organ Toxicity in STZ-Induced Diabetic Cardiomyopathy. Biomedicines. 2020; 8(2):29. https://doi.org/10.3390/biomedicines8020029
Chicago/Turabian StyleAlabi, Toyin D., Novel N. Chegou, Nicole L. Brooks, and Oluwafemi O. Oguntibeju. 2020. "Effects of Anchomanes difformis on Inflammation, Apoptosis, and Organ Toxicity in STZ-Induced Diabetic Cardiomyopathy" Biomedicines 8, no. 2: 29. https://doi.org/10.3390/biomedicines8020029