Predictors of HbA1c among Adipocytokine Biomarkers in African-American Men with Varied Glucose Tolerance

 , and
, and
Abstract
:1. Introduction
2. Experimental Section
2.1. Design and Participants
2.2. Biometrics and Glycemic Measures
2.3. Adipocytokine Biomarker Analysis
2.4. Statistical Analysis
3. Results
3.1. Study Population Characteristics and Glycemic Indices
3.2. Adipocytokines and Diabetes Status
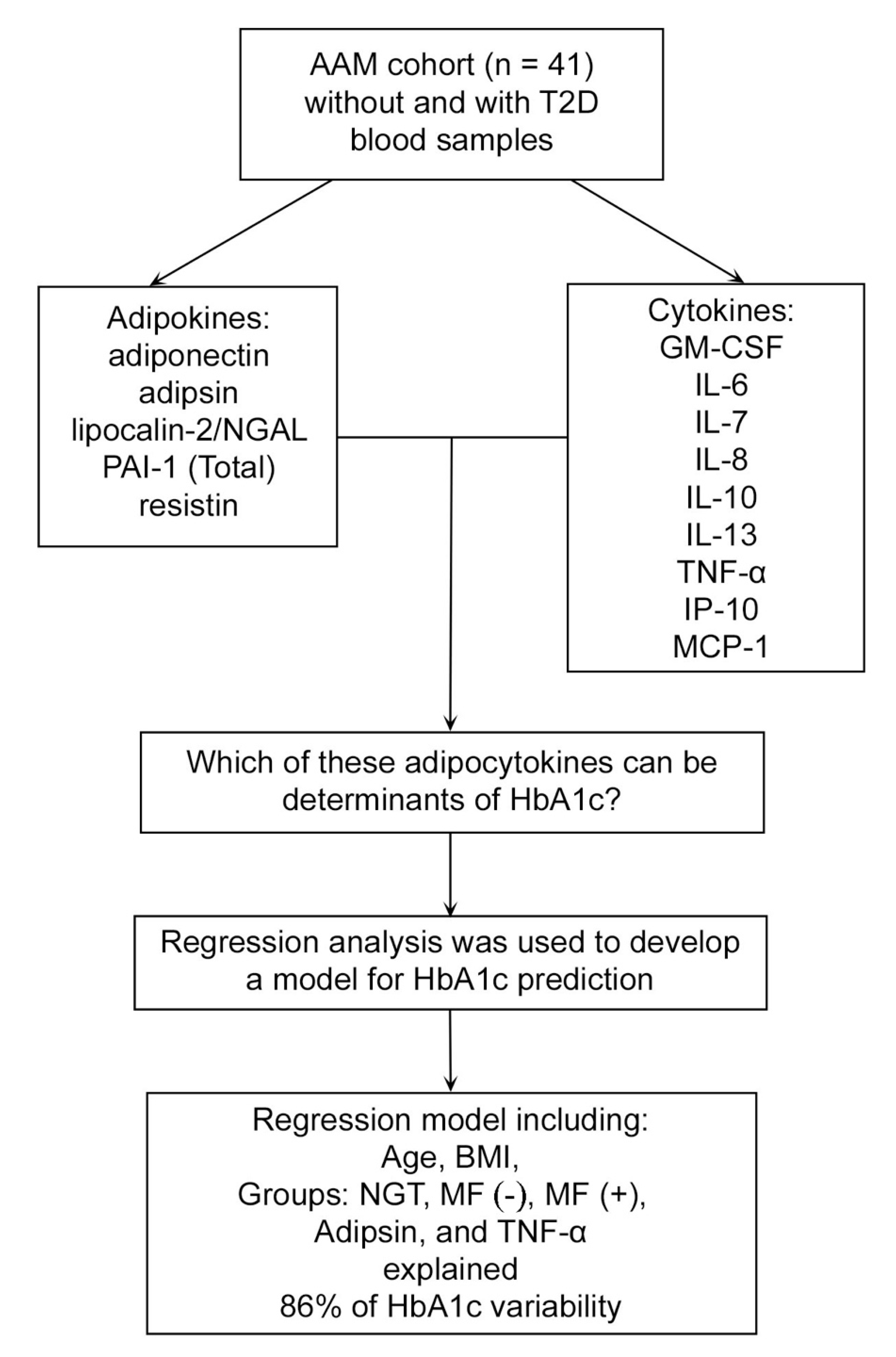
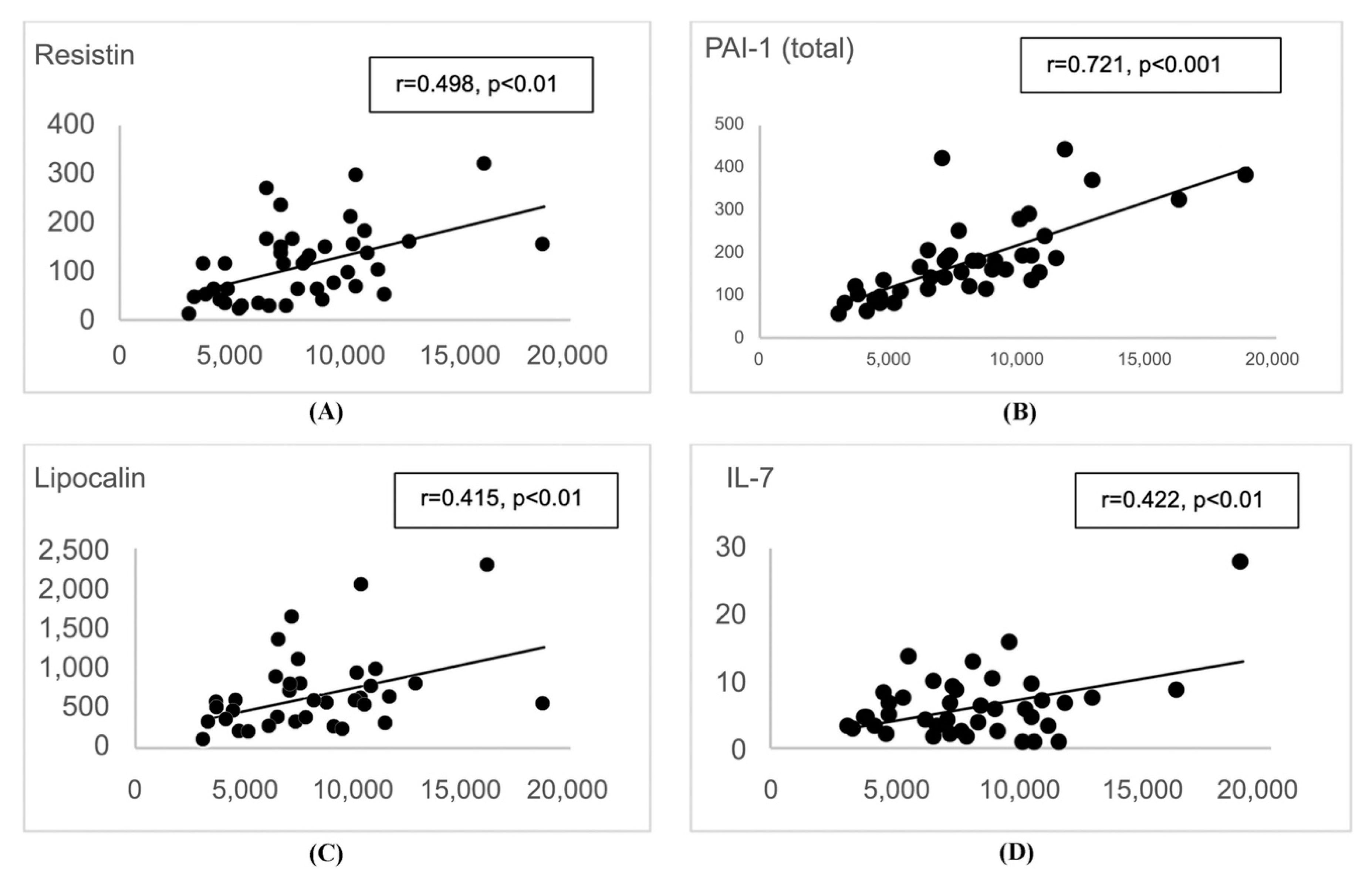
3.3. Adipocytokine Predictors of HbA1c
4. Discussion
4.1. Adipokine Adipsin as a Predictor of HbA1c
4.2. Cytokine TNF-α as a Predictor of HbA1c
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. National Diabetes Statistics Report 2017; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2017.
- WHO Diabetes; WHO: Geneva, Switzerland, 2016.
- American Diabetes Association. 5. Lifestyle Management: Standards of Medical Care in Diabetes-2019. Diabetes Care 2019, 42 (Suppl. 1), S46–S60. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. 3. Prevention or Delay of Type 2 Diabetes: Standards of Medical Care in Diabetes-2019. Diabetes Care 2019, 42 (Suppl. 1), S29–S33. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. 9. Pharmacologic Approaches to Glycemic Treatment: Standards of Medical Care in Diabetes-2019. Diabetes Care 2019, 42 (Suppl. 1), S90–S102. [Google Scholar] [CrossRef] [Green Version]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Primers 2015, 1, 15019. [Google Scholar] [CrossRef]
- Liu, C.; Feng, X.; Li, Q.; Wang, Y.; Li, Q.; Hua, M. Adiponectin, TNF-alpha and inflammatory cytokines and risk of type 2 diabetes: A systematic review and meta-analysis. Cytokine 2016, 86, 100–109. [Google Scholar] [CrossRef]
- Hang, H.; Yuan, S.; Yang, Q.; Yuan, D.; Liu, Q. Multiplex bead array assay of plasma cytokines in type 2 diabetes mellitus with diabetic retinopathy. Mol. Vis. 2014, 20, 1137–1145. [Google Scholar]
- Lucas, R.; Parikh, S.J.; Sridhar, S.; Guo, D.H.; Bhagatwala, J.; Dong, Y.; Caldwell, R.; Mellor, A.; Caldwell, W.; Zhu, H.; et al. Cytokine profiling of young overweight and obese female African American adults with prediabetes. Cytokine 2013, 64, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Cox, A.J.; Zhang, P.; Bowden, D.W.; Devereaux, B.; Davoren, P.M.; Cripps, A.W.; West, N.P. Enteroendocrine and adipokine associations with type 2 diabetes: Phenotypic risk scoring approaches. J. Gastroenterol. Hepatol. 2018, 33, 1357–1364. [Google Scholar] [CrossRef] [Green Version]
- Adela, R.; Reddy, P.N.C.; Ghosh, T.S.; Aggarwal, S.; Yadav, A.K.; Das, B.; Banerjee, S.K. Serum protein signature of coronary artery disease in type 2 diabetes mellitus. J. Transl. Med. 2019, 17, 17. [Google Scholar] [CrossRef]
- Daniele, G.; Guardado Mendoza, R.; Winnier, D.; Fiorentino, T.V.; Pengou, Z.; Cornell, J.; Andreozzi, F.; Jenkinson, C.; Cersosimo, E.; Federici, M.; et al. The inflammatory status score including IL-6, TNF-alpha, osteopontin, fractalkine, MCP-1 and adiponectin underlies whole-body insulin resistance and hyperglycemia in type 2 diabetes mellitus. Acta Diabetol. 2014, 51, 123–131. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Amato, M.C.; Pizzolanti, G.; Torregrossa, V.; Misiano, G.; Milano, S.; Giordano, C. Visceral adiposity index (VAI) is predictive of an altered adipokine profile in patients with type 2 diabetes. PLoS ONE 2014, 9, e91969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, P.; Spiegelman, B.M. Cell biology of fat storage. Mol. Biol. Cell. 2016, 27, 2523–2527. [Google Scholar] [CrossRef] [PubMed]
- Elekofehinti, O.O.; Ejelonu, O.C.; Kamdem, J.P.; Akinlosotu, O.B.; Adanlawo, I.G. Saponins as adipokines modulator: A possible therapeutic intervention for type 2 diabetes. World J. Diabetes 2017, 8, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Leroith, D.; Accili, D. Mechanisms of disease: Using genetically altered mice to study concepts of type 2 diabetes. Nat. Clin. Pract. Endocrinol. Metab. 2008, 4, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Treuter, E.; Fan, R.; Huang, Z.; Jakobsson, T.; Venteclef, N. Transcriptional repression in macrophages-basic mechanisms and alterations in metabolic inflammatory diseases. FEBS Lett. 2017, 591, 2959–2977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasilenko, M.A.; Kirienkova, E.V.; Skuratovskaia, D.A.; Zatolokin, P.A.; Mironyuk, N.I.; Litvinova, L.S. The role of production of adipsin and leptin in the development of insulin resistance in patients with abdominal obesity. Dokl. Biochem. Biophys. 2017, 475, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Domingueti, C.P.; Dusse, L.M.; Carvalho, M.; de Sousa, L.P.; Gomes, K.B.; Fernandes, A.P. Diabetes mellitus: The linkage between oxidative stress, inflammation, hypercoagulability and vascular complications. J. Diabetes Complicat. 2016, 30, 738–745. [Google Scholar] [CrossRef]
- Hua, F.; Yu, J.J.; Hu, Z.W. Diabetes and cancer, common threads and missing links. Cancer Lett. 2016, 374, 54–61. [Google Scholar] [CrossRef]
- Wintrob, Z.A.; Hammel, J.P.; Khoury, T.; Nimako, G.K.; Fu, H.W.; Fayazi, Z.S.; Gaile, D.P.; Forrest, A.; Ceacareanu, A.C. Insulin use, adipokine profiles and breast cancer prognosis. Cytokine 2017, 89, 45–61. [Google Scholar] [CrossRef]
- Pavan Kumar, N.; Nair, D.; Banurekha, V.V.; Dolla, C.; Kumaran, P.; Sridhar, R.; Babu, S. Type 2 diabetes mellitus coincident with pulmonary or latent tuberculosis results in modulation of adipocytokines. Cytokine 2016, 79, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Lu, Y.; Ma, D.L.; Du, T.T.; Shao, S.Y.; Yu, X.F. A meta-analysis of salicylates for type 2 diabetes mellitus. J. Huazhong Univ. Sci. Technol. Med. Sci. 2013, 33, 1–14. [Google Scholar] [CrossRef] [PubMed]
- McKie, E.A.; Reid, J.L.; Mistry, P.C.; DeWall, S.L.; Abberley, L.; Ambery, P.D.; Gil-Extremera, B. A Study to Investigate the Efficacy and Safety of an Anti-Interleukin-18 Monoclonal Antibody in the Treatment of Type 2 Diabetes Mellitus. PLoS ONE 2016, 11, e0150018. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Mukheem, A.; Kamarul, T. The prevention and treatment of hypoadiponectinemia-associated human diseases by up-regulation of plasma adiponectin. Life Sci. 2015, 135, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Fain, J.N. Release of interleukins and other inflammatory cytokines by human adipose tissue is enhanced in obesity and primarily due to the nonfat cells. Vitam. Horm. 2006, 74, 443–477. [Google Scholar]
- Hamada, M.; Abe, M.; Miyake, T.; Kawasaki, K.; Tada, F.; Furukawa, S.; Matsuura, B.; Hiasa, Y.; Onji, M. B cell-activating factor controls the production of adipokines and induces insulin resistance. Obesity (Silver Spring) 2011, 19, 1915–1922. [Google Scholar] [CrossRef]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Binder, B.R.; Christ, G.; Gruber, F.; Grubic, N.; Hufnagl, P.; Krebs, M.; Mihaly, J.; Prager, G.W. Plasminogen activator inhibitor 1: Physiological and pathophysiological roles. News Physiol. Sci. 2002, 17, 56–61. [Google Scholar] [CrossRef]
- Zhao, R.; Le, K.; Moghadasian, M.H.; Shen, G.X. Reduced monocyte adhesion to aortae of diabetic plasminogen activator inhibitor-1 knockout mice. Inflamm. Res. 2017, 66, 783–792. [Google Scholar] [CrossRef]
- Anderson, W.D.; Makadia, H.K.; Greenhalgh, A.D.; Schwaber, J.S.; David, S.; Vadigepalli, R. Computational modeling of cytokine signaling in microglia. Mol. Biosyst. 2015, 11, 3332–3346. [Google Scholar] [CrossRef] [Green Version]
- Sinha, N.; Sharma, S.; Tripathi, P.; Negi, S.K.; Tikoo, K.; Kumar, D.; Rao, K.V.; Chatterjee, S. Molecular signatures for obesity and associated disorders identified through partial least square regression models. BMC Syst. Biol. 2014, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef] [Green Version]
- Breen, E.J.; Tan, W.; Khan, A. The Statistical Value of Raw Fluorescence Signal in Luminex xMAP Based Multiplex Immunoassays. Sci. Rep. 2016, 6, 26996. [Google Scholar] [CrossRef]
- Ozanne, S.E.; Rahmoune, H.; Guest, P.C. Multiplex Biomarker Approaches in Type 2 Diabetes Mellitus Research. Methods Mol. Biol. 2017, 1546, 37–55. [Google Scholar]
- Nowak, C.; Carlsson, A.C.; Ostgren, C.J.; Nystrom, F.H.; Alam, M.; Feldreich, T.; Sundstrom, J.; Carrero, J.J.; Leppert, J.; Hedberg, P.; et al. Multiplex proteomics for prediction of major cardiovascular events in type 2 diabetes. Diabetologia 2018, 61, 1748–1757. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.E.; Lyass, A.; Courchesne, P.; Chen, G.; Liu, C.; Yin, X.; Hwang, S.J.; Massaro, J.M.; Larson, M.G.; Levy, D. Protein Biomarkers of Cardiovascular Disease and Mortality in the Community. J. Am. Heart Assoc. 2018, 7, e008108. [Google Scholar] [CrossRef] [Green Version]
- Barengolts, E.; Eisenberg, Y.; Ciubotaru, I.; Akbar, A. Circulating IP-10 and C3 Declined, and Pancreatic Polypeptide, Peptide YY, VEGF and Adiponectin Increased in Response to Oral Glucose Tolerance Test While 25(OH)D Strongly Correlated with Post- Glucose-Load Adipsin and IL-13 in African American Men with Prediabetes and Hypovitaminosis D. In Proceedings of the Endocrine Society Annual Meeting, San Diego, CA, USA, 5–8 March 2015. [Google Scholar]
- Zhou, Q.; Ge, Q.; Ding, Y.; Qu, H.; Wei, H.; Wu, R.; Yao, L.; Wei, Q.; Feng, Z.; Long, J.; et al. Relationship between serum adipsin and the first phase of glucose-stimulated insulin secretion in individuals with different glucose tolerance. J. Diabetes Investig. 2018, 9, 1128–1134. [Google Scholar] [CrossRef] [Green Version]
- Mirza, S.; Hossain, M.; Mathews, C.; Martinez, P.; Pino, P.; Gay, J.L.; Rentfro, A.; McCormick, J.B.; Fisher-Hoch, S.P. Type 2-diabetes is associated with elevated levels of TNF-alpha, IL-6 and adiponectin and low levels of leptin in a population of Mexican Americans: A cross-sectional study. Cytokine 2012, 57, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Barengolts, E.; Green, S.J.; Eisenberg, Y.; Akbar, A.; Reddivari, B.; Layden, B.T.; Dugas, L.; Chlipala, G. Gut microbiota varies by opioid use, circulating leptin and oxytocin in African American men with diabetes and high burden of chronic disease. PLoS ONE 2018, 13, e0194171. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, Y.; Dugas, L.R.; Akbar, A.; Reddivari, B.; Layden, B.T.; Barengolts, E. Oxytocin is lower in African American men with diabetes and associates with psycho-social and metabolic health factors. PLoS ONE 2018, 13, e0190301. [Google Scholar] [CrossRef] [Green Version]
- Barengolts, E.; Manickam, B.; Eisenberg, Y.; Akbar, A.; Kukreja, S.; Ciubotaru, I. Effect of High-Dose Vitamin D Repletion on Glycemic Control in African-American Males with Prediabetes and Hypovitaminosis D. Endocr Pract 2015, 21, 604–612. [Google Scholar] [CrossRef] [Green Version]
- Seder, C.W.; Kubasiak, J.C.; Pithadia, R.; Basu, S.; Fhied, C.; Tarhoni, I.; Davila, E.; Alnajjar, H.; Chmielewski, G.W.; Warren, W.H.; et al. Angiogenesis Biomarkers May Be Useful in the Management of Patients With Indeterminate Pulmonary Nodules. Ann. Thorac. Surg. 2015, 100, 429–436. [Google Scholar] [CrossRef]
- Fidler, M.J.; Frankenberger, C.; Seto, R.; Lobato, G.C.; Fhied, C.L.; Sayidine, S.; Basu, S.; Pool, M.; Karmali, R.; Batus, M.; et al. Differential expression of circulating biomarkers of tumor phenotype and outcomes in previously treated non-small cell lung cancer patients receiving erlotinib vs. cytotoxic chemotherapy. Oncotarget 2017, 8, 58108–58121. [Google Scholar] [CrossRef]
- Loo, B.M.; Marniemi, J.; Jula, A. Evaluation of multiplex immunoassays, used for determination of adiponectin, resistin, leptin, and ghrelin from human blood samples, in comparison to ELISA assays. Scand. J. Clin. Lab. Investig. 2011, 71, 221–226. [Google Scholar] [CrossRef]
- Schipper, H.S.; de Jager, W.; van Dijk, M.E.; Meerding, J.; Zelissen, P.M.; Adan, R.A.; Prakken, B.J.; Kalkhoven, E. A multiplex immunoassay for human adipokine profiling. Clin. Chem. 2010, 56, 1320–1328. [Google Scholar] [CrossRef]
- Richens, J.L.; Urbanowicz, R.A.; Metcalf, R.; Corne, J.; O’Shea, P.; Fairclough, L. Quantitative validation and comparison of multiplex cytokine kits. J. Biomol. Screen. 2010, 15, 562–568. [Google Scholar] [CrossRef] [Green Version]
- Elshal, M.F.; McCoy, J.P. Multiplex bead array assays: Performance evaluation and comparison of sensitivity to ELISA. Methods 2006, 38, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Cook, K.S.; Min, H.Y.; Johnson, D.; Chaplinsky, R.J.; Flier, J.S.; Hunt, C.R.; Spiegelman, B.M. Adipsin: A circulating serine protease homolog secreted by adipose tissue and sciatic nerve. Science 1987, 237, 402–405. [Google Scholar] [CrossRef]
- Flier, J.S.; Cook, K.S.; Usher, P.; Spiegelman, B.M. Severely impaired adipsin expression in genetic and acquired obesity. Science 1987, 237, 405–408. [Google Scholar] [CrossRef]
- Rosen, B.S.; Cook, K.S.; Yaglom, J.; Groves, D.L.; Volanakis, J.E.; Damm, D.; White, T.; Spiegelman, B.M. Adipsin and complement factor D activity: An immune-related defect in obesity. Science 1989, 244, 1483–1487. [Google Scholar] [CrossRef]
- Lo, J.C.; Ljubicic, S.; Leibiger, B.; Kern, M.; Leibiger, I.B.; Moede, T.; Kelly, M.E.; Chatterjee Bhowmick, D.; Murano, I.; Cohen, P.; et al. Adipsin is an adipokine that improves beta cell function in diabetes. Cell 2014, 158, 41–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legakis, I.; Mantzouridis, T.; Bouboulis, G.; Chrousos, G.P. Reciprocal changes of serum adispin and visfatin levels in patients with type 2 diabetes after an overnight fast. Arch. Endocrinol. Metab. 2016, 60, 76–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.S.; Lee, W.J.; Lee, I.T.; Lin, S.Y.; Lee, W.L.; Liang, K.W.; Sheu, W.H. Association Between Serum Adipsin Levels and Insulin Resistance in Subjects With Various Degrees of Glucose Intolerance. J. Endocr. Soc. 2019, 3, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Whaley, K. Biosynthesis of the complement components and the regulatory proteins of the alternative complement pathway by human peripheral blood monocytes. J. Exp. Med. 1980, 151, 501–516. [Google Scholar] [CrossRef]
- Su, X.; Yan, H.; Huang, Y.; Yun, H.; Zeng, B.; Wang, E.; Liu, Y.; Zhang, Y.; Liu, F.; Che, Y.; et al. Expression of FABP4, adipsin and adiponectin in Paneth cells is modulated by gut Lactobacillus. Sci. Rep. 2015, 5, 18588. [Google Scholar] [CrossRef] [Green Version]
- Volanakis, J.E.; Narayana, S.V. Complement factor D, a novel serine protease. Protein Sci. 1996, 5, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Plante, E.; Menaouar, A.; Danalache, B.A.; Yip, D.; Broderick, T.L.; Chiasson, J.L.; Jankowski, M.; Gutkowska, J. Oxytocin treatment prevents the cardiomyopathy observed in obese diabetic male db/db mice. Endocrinology 2015, 156, 1416–1428. [Google Scholar] [CrossRef]
- Nashef, A.; Abu-Toamih Atamni, H.J.; Buchnik, Y.; Hasturk, H.; Kantarci, A.; Stephens, D.; Wiess, E.I.; Houri-Haddad, Y.; Iraqi, F.A. Collaborative Cross Mouse Population for Studying Alveolar Bone Changes and Impaired Glucose Tolerance Comorbidity After High-Fat Diet Consumption. J. Periodontol. 2017, 88, e150–e158. [Google Scholar] [CrossRef]
- Bhavsar, S.K.; Singh, S.; Giri, S.; Jain, M.R.; Santani, D.D. Effect of saponins from Helicteres isora on lipid and glucose metabolism regulating genes expression. J. Ethnopharmacol. 2009, 124, 426–433. [Google Scholar] [CrossRef]
- Coulthard, L.G.; Woodruff, T.M. Is the complement activation product C3a a proinflammatory molecule? Re-evaluating the evidence and the myth. J. Immunol. 2015, 194, 3542–3548. [Google Scholar] [CrossRef] [Green Version]
- Spranger, J.; Kroke, A.; Mohlig, M.; Hoffmann, K.; Bergmann, M.M.; Ristow, M.; Boeing, H.; Pfeiffer, A.F. Inflammatory cytokines and the risk to develop type 2 diabetes: Results of the prospective population-based European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Study. Diabetes 2003, 52, 812–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ofei, F.; Hurel, S.; Newkirk, J.; Sopwith, M.; Taylor, R. Effects of an engineered human anti-TNF-alpha antibody (CDP571) on insulin sensitivity and glycemic control in patients with NIDDM. Diabetes 1996, 45, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, Y.; Kiyohara, C.; Kashiwado, Y.; Sawabe, T.; Nagano, S.; Kimoto, Y.; Ayano, M.; Mitoma, H.; Akahoshi, M.; Arinobu, Y.; et al. Effects of tumor necrosis factor inhibitors and tocilizumab on the glycosylated hemoglobin levels in patients with rheumatoid arthritis; an observational study. PLoS ONE 2018, 13, e0196368. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Budavari, A.; Murray, D.; Spiegelman, B.M. Reduced tyrosine kinase activity of the insulin receptor in obesity-diabetes. Central role of tumor necrosis factor-alpha. J. Clin. Investig. 1994, 94, 1543–1549. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Peraldi, P.; Budavari, A.; Ellis, R.; White, M.F.; Spiegelman, B.M. IRS-1-mediated inhibition of insulin receptor tyrosine kinase activity in TNF-alpha- and obesity-induced insulin resistance. Science 1996, 271, 665–668. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef]
- Esposito, K.; Nappo, F.; Marfella, R.; Giugliano, G.; Giugliano, F.; Ciotola, M.; Quagliaro, L.; Ceriello, A.; Giugliano, D. Inflammatory cytokine concentrations are acutely increased by hyperglycemia in humans: Role of oxidative stress. Circulation 2002, 106, 2067–2072. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Variable | NGT (n = 20) | T2D (n = 21) | ||
|---|---|---|---|---|
| MF(+) (n = 16) | MF(−) (n = 5) | Combined | ||
| (n = 21) | ||||
| General | ||||
| Age, y | 54.1 ± 5.8 | 60.1 ± 3.1 ** | 56.6 ± 6.7 | 59.2 ± 4.3 ** |
| Body weight, kg | 84.7 ± 11.5 | 105.5 ± 14.2 ** | 106.8 ± 14.7 ** | 105.8 ± 13.9 ** |
| BMI, kg/m2 | 27.4 ± 3.3 | 35.4 ± 3.2 ** | 34.8 ± 2.4 ** | 35.3 ± 3.0 ** |
| Charlson index | 1.7 ± 1.1 | 3.4 ± 1.4 ** | 1.2 ± 0.6 | 2.9 ± 1.5 * |
| Number of all meds | 8.3 ± 5.0 | 11.3 ± 4.9 | 2.2 ± 1.1 | 9.1 ± 5.0 |
| Fasting Glycemic Indices | ||||
| HbA1c, % | 5.3 ± 0.3 | 6.9 ± 0.5 ** | 6.8 ± 0.2 ** | 6.8 ± 0.4 ** |
| Fasting glucose, mg/dL | 93.6 ± 17.4 | 123.6 ± 27.8 | 119.8 ± 24.0 | 122.7 ± 26.4 |
| Fasting Insulin, mIU/L | 8.1 ± 3.9 | 35.9 ± 55.7 | 19.7 ± 5.1 | 32.1 ± 48.8 |
| 120 min Glycemic Indices | ||||
| Glucose 120 min, mg/dL | 109.6 ± 42.0 | 198.1 ± 70.1 | 187.4 ± 100.0 | 195.6 ± 75.5 |
| Insulin 120 min, mIU/L | 43.3 ± 45.1 | 104.2 ± 63.7 | 67.9 ± 25.6 | 95.6 ± 58.5 |
| Variable | NGT (n = 20) | T2D (n = 21) | ||
|---|---|---|---|---|
| MF(+) (n = 16) | MF(−) (n = 5) | Combined | ||
| (n = 21) | ||||
| Fasting Cytokines | ||||
| GM-CSF | 20.3 ± 15.3 | 21.4 ± 27.1 | 26.1 ± 28.9 | 22.5 ± 26.8 |
| IL-10 | 7.3 ± 4.9 | 7.3 ± 6.7 | 6.4 ± 4.4 | 7.1 ± 6.1 |
| IL-13 | 15.3 ± 41.1 | 9.7 ± 12.0 * | 11.0 ± 14.3 | 10.0 ± 12.1 * |
| IL-6 | 6.3 ± 11.5 | 2.5 ± 2.8 ** | 3.7 ± 5.2 * | 2.8 ± 3.4 ** |
| IL-7 | 5.4 ± 3.2 | 6.9 ± 6.3 | 7.4 ± 6.9 | 7.0 ± 6.3 |
| IL-8 | 74.9 ± 91.3 | 13.4 ± 21.8 ** | 9.4 ± 8.0 * | 12.4 ± 19.3 |
| TNF-α | 14.5 ± 14.2 | 4.5 ± 2.7 ** | 5.0 ± 2.8 ** | 4.6 ± 2.7 ** |
| 120 min Cytokines | ||||
| GM-CSF | 19.8 ± 14.8 | 20.8 ± 28.2 | 20.2 ± 21.6 | 20.6 ± 26.3 |
| IL-10 | 7.2 ± 4.7 | 6.9 ± 6.9 | 6.4 ± 3.6 | 6.8 ± 6.3 |
| IL-13 | 15.4 ± 41.5 | 9.8 ± 13.0 ** | 10.9 ±9.9 | 10.0 ± 12.3 ** |
| IL-6 | 4.0 ± 9.3 | 2.7 ± 3.0 * | 2.5 ± 3.4 * | 2.6 ± 3.0 ** |
| IL-7 | 5.1 ± 3.0 | 6.4 ± 6.9 | 5.7 ± 5.7 | 6.2 ± 6.5 |
| IL-8 | 11.6 ± 14.9 | 5.0 ± 4.8 ** | 3.9 ± 1.3 * | 4.7 ± 4.2 ** |
| TNF-α | 9.9 ± 10.3 | 4.3 ± 3.5 ** | 4.4 ± 2.2 ** | 4.3 ± 3.2 ** |
| Variable | NGT (n = 20) | T2D (n = 21) | ||
|---|---|---|---|---|
| MF(+) (n = 16) | MF(−) (n = 5) | Combined | ||
| (n = 21) | ||||
| Fasting Adipokines | ||||
| Adiponectin | 60,785 ± 64,564 | 13,495 ± 15,055 * | 12,227 ± 6467 | 13,193 ± 13,366 * |
| Lipocalin | 897 ± 545 | 407 ± 204 * | 399 ± 225 * | 406 ± 203 * |
| Resistin | 139 ± 89 | 89 ± 53 | 70 ± 56 | 84 ± 53 |
| Adipsin | 8758 ± 3037 | 7524 ± 4097 ** | 7469 ± 1512 | 7511 ± 3612 * |
| PAI-1 (total) | 196 ± 101 | 168 ± 100 ** | 161 ± 53 * | 166 ± 90 ** |
| IP-10 | 409 ± 183 | 441 ± 243 | 286 ± 52 | 404 ± 222 |
| MCP-1 | 434 ± 320 | 335 ± 147 | 438 ± 355 | 359 ± 20 |
| 120 min Adipokines | ||||
| Adiponectin | 58,965 ± 67,994 | 10,190 ± 5774 ** | 9518 ± 1579 ** | 10,030 ± 5827 ** |
| Lipocalin | 568 ± 283 | 282 ± 144 | 273 ± 196 | 280 ± 152 * |
| Resistin | 99 ± 60 | 66 ± 43 | 44 ± 42 * | 61 ± 43 * |
| Adipsin | 7947 ± 2773 | 6746 ± 5154 ** | 5364 ± 1579 ** | 6417 ± 4559 ** |
| PAI-1 (total) | 185 ± 111 | 141 ± 96 ** | 124 ± 84 ** | 137 ± 92 ** |
| IP-10 | 345 ± 136 | 348 ± 207 | 245 ± 41.4 | 324 ± 125 |
| MCP-1 | 384 ± 286 | 291 ± 131 | 396 ± 261 | 316 ± 170 |
| Independent Variable | β-Coefficient (SE) | p-Value |
|---|---|---|
| GM-CSF | −0.14 (0.08) | NS |
| IL-10 | −0.02 (0.07) | NS |
| IL-13 | −0.05 (0.07) | NS |
| IL-6 | −0.08 (0.07) | NS |
| IL-7 | −0.01 (0.11) | NS |
| IL-8 | −0.14 (0.06) | 0.032 |
| TNF-α | −0.42 (0.09) | <0.001 |
| Adiponectin | -0.28 (0.09) | 0.003 |
| Lipocalin | −0.42 (0.14) | 0.005 |
| Resistin | −0.33 (0.10) | 0.003 |
| Adipsin | −0.0001 (0.00002) | <0.001 |
| PAI-1 (total) | −0.67 (0.14) | <0.001 |
| IP-10 | 0.11 (0.21) | NS |
| MCP-1 | −0.02 (0.18) | NS |
| Independent Variables | Beta (SE) | 95% CI | p-Value |
|---|---|---|---|
| TNF-α | −0.16 (0.09) | −0.34, 0.03 | 0.088 |
| Adipsin | −0.43 (0.14) | −0.72, −0.15 | 0.004 |
| Age (years) | 0.03 (0.01) | 0.004, 0.05 | 0.025 |
| BMI (kg/m2) | 0.04 (0.02) | 0.002, 0.08 | 0.036 |
| NGT | Reference | - | - |
| MF(−) | 0.93 (0.26) | 0.40, 1.45 | 0.001 |
| MF(+) | 0.86 (0.26) | 0.32, 1.10 | 0.003 |
| Variable | VIF | 1/VIF |
|---|---|---|
| TNF-α | 1.93 | 0.519311 |
| Adipsin | 1.22 | 0.818808 |
| Age (years) | 1.49 | 0.670785 |
| BMI (kg/m2) | 3.56 | 0.280930 |
| NGT | - | - |
| MF(−) | 2.72 | 0.367760 |
| MF(+) | 6.11 | 0.163683 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barengolts, E.; Akbar, A.; Layden, B.T.; Eisenberg, Y.; Priyadarshini, M.; Borgia, J.A.; Fhied, C.L.; Salim, M.; Dugas, L.R. Predictors of HbA1c among Adipocytokine Biomarkers in African-American Men with Varied Glucose Tolerance. Biomedicines 2020, 8, 520. https://doi.org/10.3390/biomedicines8110520
Barengolts E, Akbar A, Layden BT, Eisenberg Y, Priyadarshini M, Borgia JA, Fhied CL, Salim M, Dugas LR. Predictors of HbA1c among Adipocytokine Biomarkers in African-American Men with Varied Glucose Tolerance. Biomedicines. 2020; 8(11):520. https://doi.org/10.3390/biomedicines8110520
Chicago/Turabian StyleBarengolts, Elena, Arfana Akbar, Brian T. Layden, Yuval Eisenberg, Medha Priyadarshini, Jeffrey A. Borgia, Cristina L. Fhied, Michael Salim, and Lara R. Dugas. 2020. "Predictors of HbA1c among Adipocytokine Biomarkers in African-American Men with Varied Glucose Tolerance" Biomedicines 8, no. 11: 520. https://doi.org/10.3390/biomedicines8110520