
Regenerative Therapy for Corneal Scarring Disorders
 , ,
, ,
Abstract
:1. Introduction
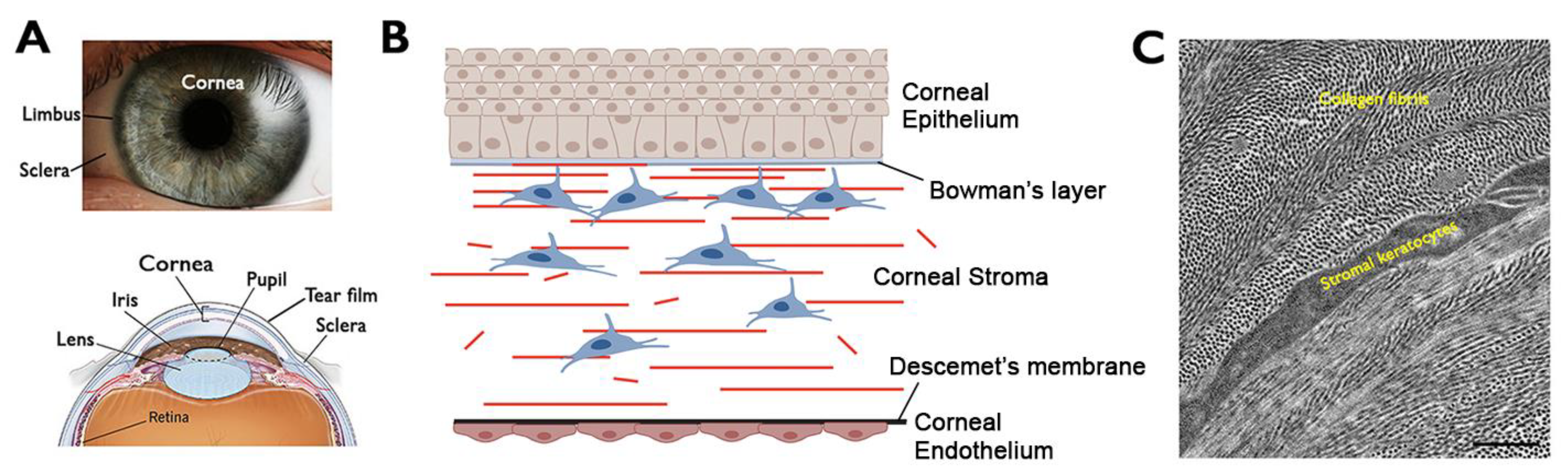
2. Cornea—Structure and Functions
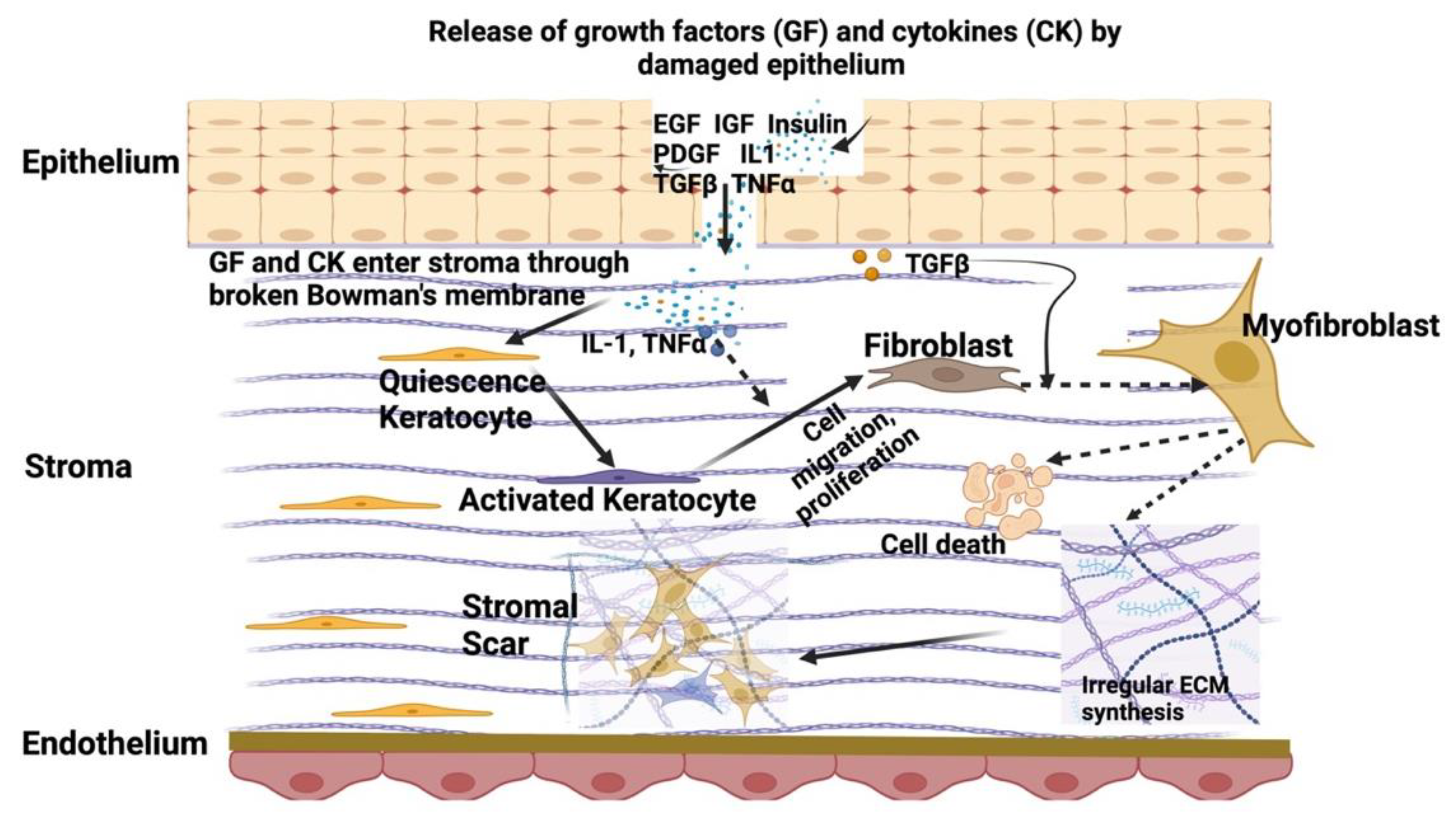
3. Corneal Stromal Homeostasis and Wound Healing
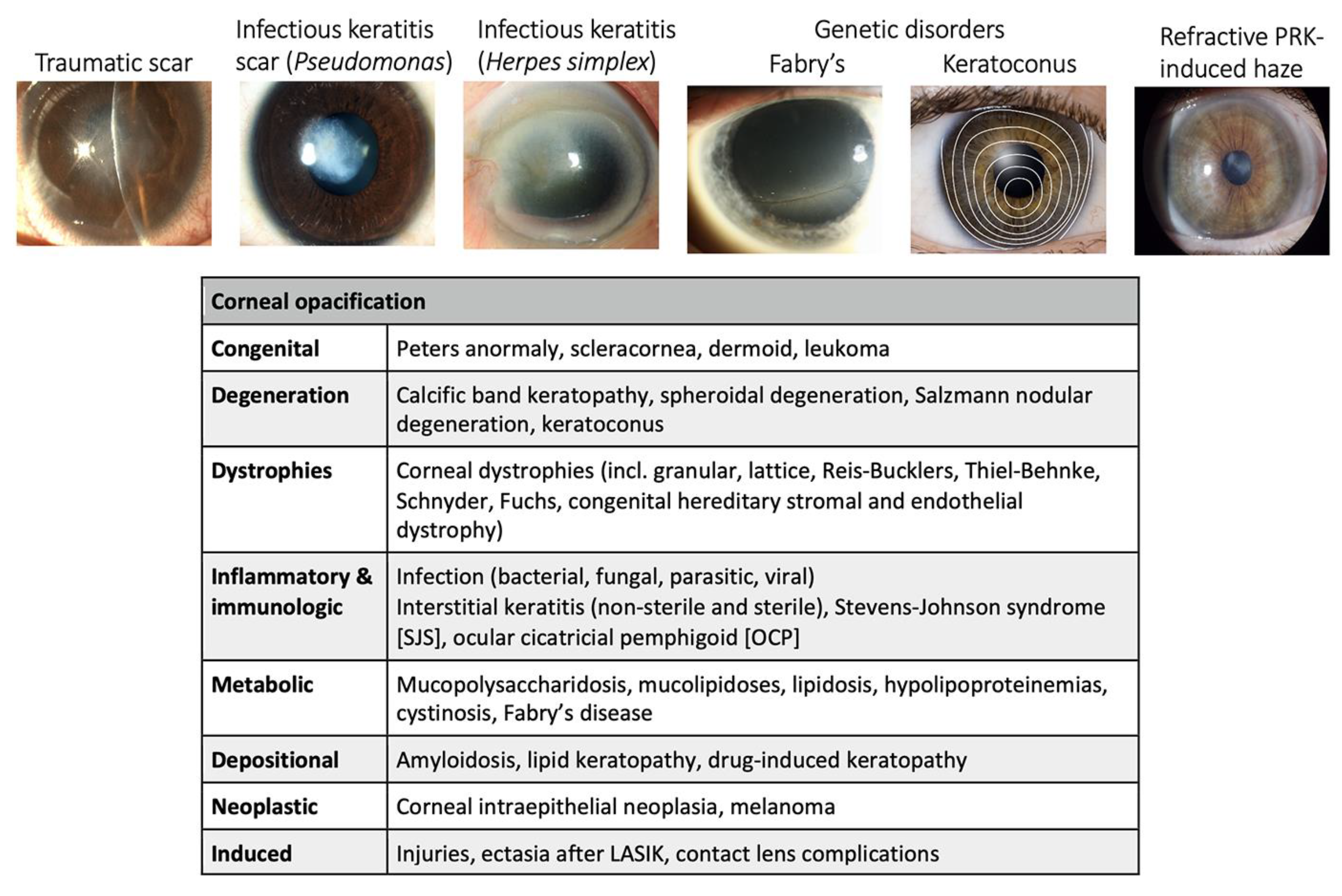
4. Etiology of Corneal Scarring Disorders
4.1. Ocular Trauma
4.2. Corneal Infection
4.3. Chemical Injury
4.4. Ocular Refractive Surgeries
4.5. Acquired and Inherited Corneal Disorders
5. Current Clinical Management of Corneal Scarring
5.1. Topical Antibiotics
5.2. Topical Corticosteroids
5.3. Mitomycin C (MMC)
5.4. Amniotic Membrane (AM) Grafting
5.5. Collagen-Based Hydrogel
6. Emerging Therapeutic Strategies for Stromal Regeneration and Scar Inhibition
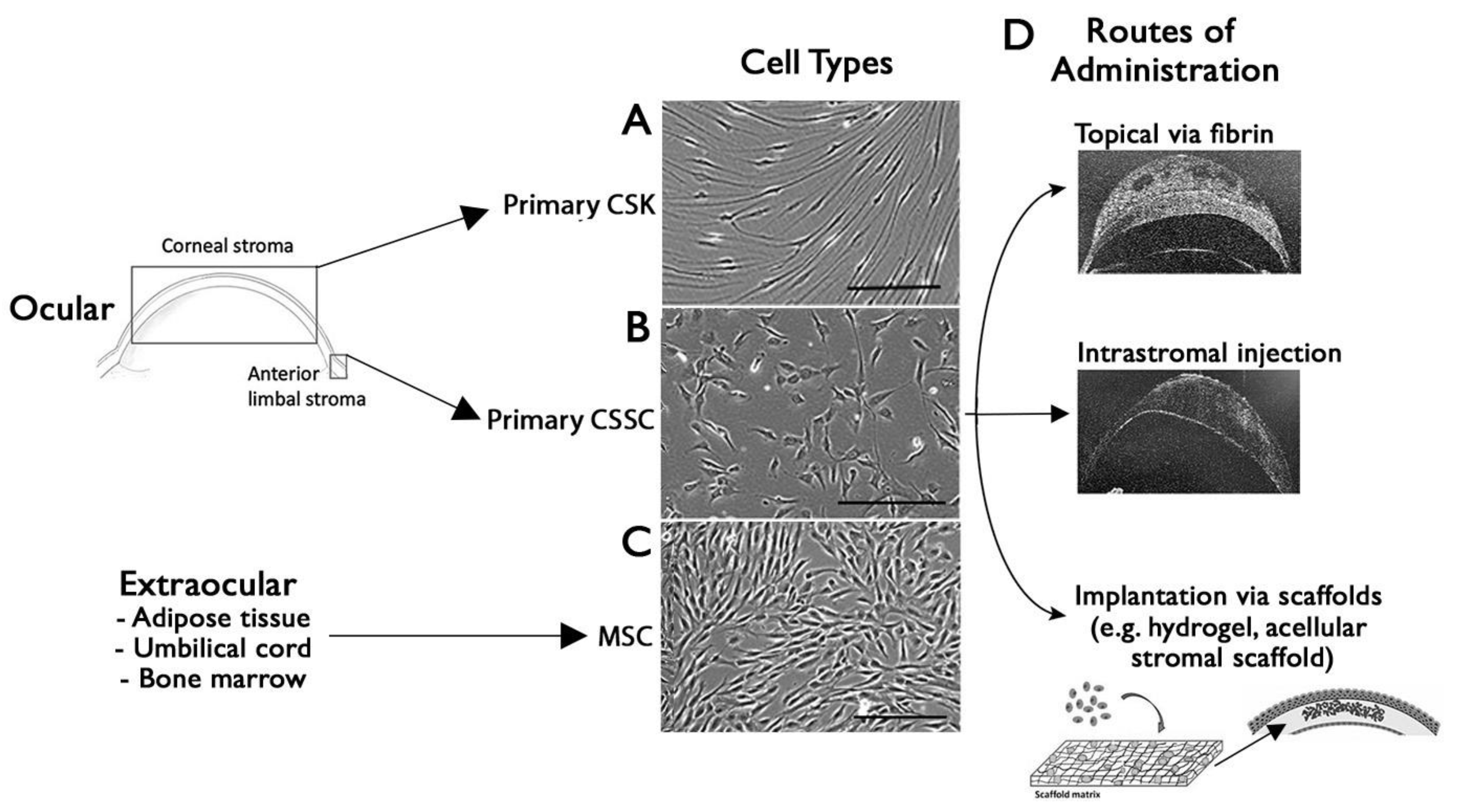
7. Cell-Based Approach for Corneal Wound Healing and Scar Management
7.1. Stromal Keratocytes as a Novel Therapeutic Tool for Scar Inhibition
7.2. Stem Cell Therapy for a Scarless Corneal Stromal Regeneration
8. Cell-Free Approach for Stromal Wound Healing and Regeneration
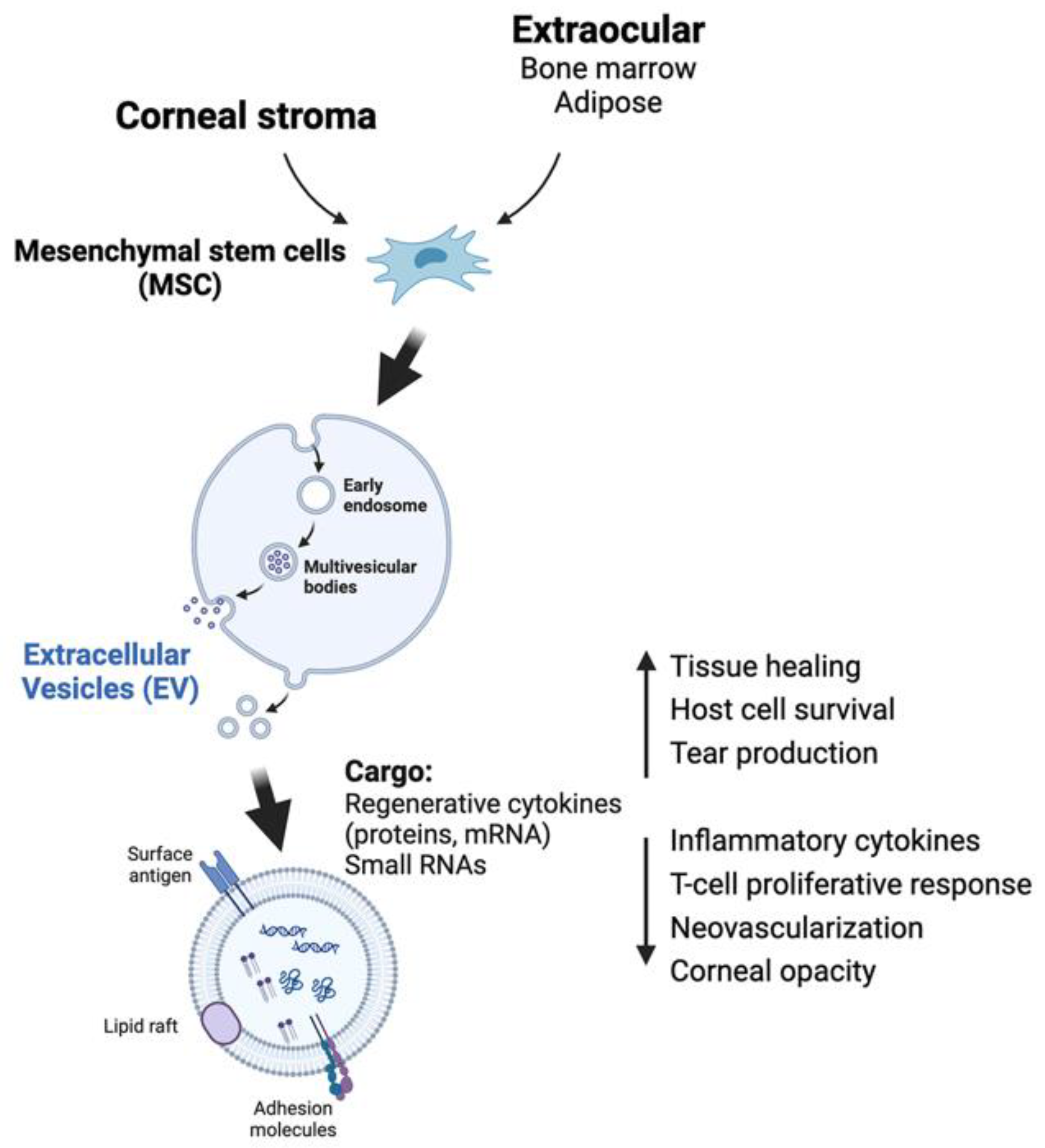
8.1. Extracellular Vesicles (EVs)
8.2. The Extracellular Matrix (ECM)
9. Molecular Approach in Corneal Wound Healing
9.1. Hevin
9.2. Krüppel-like Factor 4 (KLF4)
9.3. Inhibitor of Differentiation 3 (Id3)
9.4. SMAD7
9.5. Bone Morphogenic Protein 7 (BMP7)
9.6. Decorin
9.7. Regenerative Biomolecules and Immunomodulators to Route Scar-Forming Healing to Scar-Free Healing
9.7.1. HDAC Inhibitor
9.7.2. Members of TGFβ Family
9.7.3. Losartan
9.7.4. Hepatocyte Growth Factor (HGF)
9.7.5. Lumikine
10. Targeted Gene Silencing to Prevent Corneal Scarring
11. Tissue Engineering Approach for Corneal Regeneration
11.1. Stromal Lenticule Engineering
11.2. Synthetic Non-Collagen-Based Scaffolds
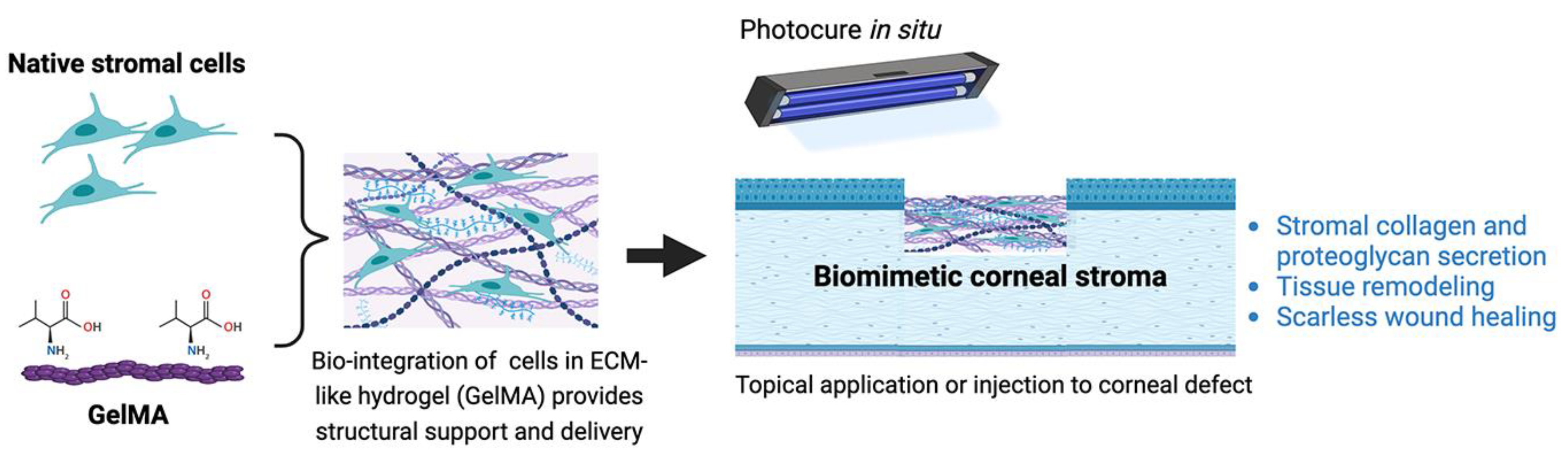
11.2.1. Gelatin-Based Hydrogels
11.2.2. Silk Fibroin
11.2.3. Chitosan
12. Summary and Future Perspectives

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Approach | Types | Mechanisms of Action | Risks/Potential Side Effects | Limitations |
|---|---|---|---|---|
| Cell-based | Corneal stromal keratocytes | Produce and deposit native stromal collagen and proteoglycans to restore ECM composition | Transit to fibroblasts and MyoF under wound conditions, need to apply after pro-inflammatory and fibrotic cytokines are suppressed [9] | Low cell yield due to slow ex vivo expansion [15] |
| Corneal stromal stem cells | Anti-inflammatory with TSG-6 expression; anti-fibrosis with TGFβ3 expression; differentiation to keratocytes [98,137] | Cell fate and phenotypic variation in response to pH changes and inflammatory response in corneal wound | Donor to donor variation in cell characteristics and functions [99] | |
| Mesenchymal stem cells from adipose, bone marrow | Anti-inflammatory; immuno-modulatory; keratocyte differentiation [76,78,89] | Uncertainty in ECM production specific to corneal stroma;risk of angiogenesis [88] | Donor to donor variation in cell features | |
| Cell-free | Extracellular vesicles from CSSCs, MSCs | Anti-fibrosis microRNAs (miR19a, 29a, 381) to prevent M1 macrophage activation, suppress JNK fibrotic and TGFβ pathways [110,111,112,190] | Easy application with minimal immunogenic effects. However, uncharacterized EV content results in unwanted effects. | Large-scale cell culture to prepare EVs; clearance or binding of EVs to ECM restricts cellular uptake [191] |
| Extracellular matrix | ECM microparticles reduced inflammatory and fibrotic gene expression; prevented MyoF generation [114] | Wide range of applications in different physical forms—sheets, suspension; easy to modify and functionalize | Material heterogeneity; need to develop isolation methods with high yield and purity [192] |
| ClinicalTrial.gov ID Year Initiated | Title | Target Disease, Treatment | Sponsor | Status | Publications |
|---|---|---|---|---|---|
| NCT01562002 2012 | Safety Study of Stem Cell Transplant to Treat Limbus Insufficiency Syndrome Phase I/II, double masked | LSCD | Institute of Applied Ophthalmobio-logy, Spain | Completed recruitment; N = 17 patients; No adverse effects. Improved CEpi healing | [193] |
| Allogenic bone marrow MSCs Stem cells with amniotic membrane transplant | |||||
| NCT02291770 2015 | Mesenchymal stromal cells treatment attenuates dry eye in patients with chronic graft-versus-host disease | GVHD-DED | Guangdong Provincial People’s Hospital, China | No adverse effects. In total, 12 out of 22 patients had improved dry eye score, ocular surface disease index scores, and Schirmer test results | [194,195] |
| Allogenic bone marrow MSCs Intravenous injection | |||||
| Phase III, multi-center, randomized, Open-label | |||||
| NCT02592330 2015 | Limbal Stem Cell Deficiency (LSCD) Treatment With Cultivated Stem Cell (CALEC) Graft | LSCD | Massachusetts Eye and Ear Infirmary, USA | Completed recruitment | |
| Cultivated autologous limbal epithelial cell graft | |||||
| Phase I/II, open-label | CALEC Transplant | ||||
| NCT03687632 2018 | ST266 Eye Drops for the Treatment of Persistent Corneal Epithelial Defects Phase II, multi-center, open-label | PED | Noveome Biotherapeutics | No adverse effects. A total of 10 out of 12 eyes had reduced PED area | [196] |
| Multi-cytokine biologic solution from Amnion-derived Multipotent Progenitor culture Eye drops | |||||
| NCT03878628 2019 | Treatment With Allogeneic Adipose-derived MSC in Patients With Aqueous Deficient Dry Eye Disease (MESADDE) | DED Kerato-Conjunctivitis Sicca Aqueous Tear Deficiency | Rigshospitalet, Denmark | No adverse effects. Decreased mean OSDI score, tear osmolarity; increased TBUT, Schirmer’s I test | [197] |
| Allogeneic adipose-derived MSC | |||||
| Early Phase I, open-label | Transconjunctival injection | ||||
| NCT04213248 2019 | Effect of UMSCs Derived Exosomes on Dry Eye in Patients With cGVHD Phase I/II, open-label | Dry Eye | Zhongshan Ophthalmic Center, Sun Yat-sen University, China | Recruiting No adverse effects; reduced fluorescein scores, longer tear-film breakup time; increased tear secretion; and lower OSDI scores | [198] |
| Umbilical MSC-derived exosomes Eye drops | |||||
| NCT04932629 2021 | To Evaluate the Clinical Safety and Efficacy of Limbal Stem Cell for Treatment of Superficial Corneal Pathologies | Corneal scar and opacities | L.V. Prasad Eye Institute, India | ||
| Ex vivo cultivated allogeneic limbal stromal stem cells | |||||
| Early phase I open-label | Topical with fibrin glue | ||||
| NCT05279157 2022 | Autologous Adipose-Derived Adult Stem Cell Implantation for Corneal Diseases (ADASCs-CT-CD) Phase II | Corneal dystrophy, keratoconus | Vissum, Instituto Oftalmológico de Alicante, Spain | Completed No adverse effects; improved stromal cell density, modulated scarring, visual improvement (~2 lines gain) | [81,90,199] |
| Autologous adipose MSCs Corneal implantation | |||||
| NCT06257355 2024 | Study to Evaluate the Safety and Efficacy of CSB-001 Ophthalmic Solution 0.1% in Subjects With Corneal Scars | Corneal scar | Claris Biotherapeutics, Inc. | Recruiting | |
| Human recombinant dHGF (hepatocyte growth factor) Eye drops | |||||
| Phase I Open-label |
| Genes | Mechanisms of Action | Risks/Potential Side Effects | Limitations of Approach |
|---|---|---|---|
| Hevin | Suppressed early fibrosis; reduced myoF [118,200] | Not studied | Overexpression or misexpression of genes can induce phenotypic variations and extra stress of cells. Overloading of translational and protein biosynthesis machinery leading to folding, localization, degradation, and post-translational problems. Abnormal complex formation; cellular toxicity [201,202] |
| KLF4 | Suppressed EMT and fibroblast activation; reduced SMAD2/3 phosphorylation | Negatively regulates cellular anti-viral immune response; complex effects on tumor inhibition; promotes pre-cancerous lesions [203,204] | |
| Id3 | Suppressed MyoF generation | Positively suppressed TGFβ-induced IOP elevation; relates to oncogenesis but with exceptions [205,206] | |
| SMAD7 | Reduced SMAD2/3 phosphorylation and inhibited EMT; restrained MyoF generation | Targets TGFβ receptor for proteasomal degradation; activates EGFR-signaling in carcinogenesis [207,208] | |
| BMP7 | Suppressed pro-fibrotic TGF-β/SMAD signaling and pro-inflammatory cytokine production | Risk of cancer metastasis [209,210] | |
| Decorin | Sequestered TGFβ from receptor binding and suppressed fibrosis [130,211] | Altered proteoglycan content may modulate growth factor activity [130,211] | |
| HDAC inhibitor | Inhibited histone H3 and H4 deacetylation to modulate cell growth and differentiation, suppressing fibroblast and MyoF generation. | Multiple HDACs induce opposite effects on a single event, indicating the pan-inhibitory action of HDAC inhibitor could result in unwanted effects [212] | |
| Losartan | Blocked TGFβ signaling to suppress MyoF generation and fibrosis | A well-tolerated medication with few side effects [213] | |
| HGF | Activated Smad7 to inhibit TGFβ/Smad pro-fibrotic signaling and reduced myofibroblast generation; anti-inflammatory | Pro-angiogenic activity could lead to neovascularization; HGF/c-Met signaling to trigger tumorigenesis [214] |
| Genes | Mechanism of Action | Risks/Potential Side Effects | Limitations of Approach |
|---|---|---|---|
| SEMA3A | siRNA-mediated downregulation of fibroblast/TGFβ-fibrotic pathways | Neuron polarization defects; corneal sensory alterations; risk of VEGF-mediated corneal neovascularization [215,216] | Variable knockdown efficiency by siRNAs and instability and degradation of siRNAs inside target cells. Lack of reliable delivery methods—transfection approach is poor for primary cells and electroporation induces cell death. Altered target gene expression induces phenotypic variations and altered cellular signaling [217]. |
| USP-10 | siRNA-mediated downregulation of immune cell infiltration and fibrosis gene expression | Altered de-ubiquitination modulates multiple cellular issues, e.g., protein stability [202] | |
| KCa3.1 | Using TRAM 34, an ion channel block to modulate Ca++-activated K+ signaling in fibroblast and MyoF activation; suppressed macrophages polarization towards M1 phenotype [165,218,219] | Affects cell growth and survival; triggers cell death [165,218,219] |
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Flaxman, S.R.; Bourne, R.R.A.; Resnikoff, S.; Ackland, P.; Braithwaite, T.; Cicinelli, M.V.; Das, A.; Jonas, J.B.; Keeffe, J.; Kempen, J.H.; et al. Global causes of blindness and distance vision impairment 1990–2020: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e1221–e1234. [Google Scholar] [CrossRef] [PubMed]
- Jeng, B.H.; Ahmad, S. In Pursuit of the Elimination of Corneal Blindness: Is Establishing Eye Banks and Training Surgeons Enough? Ophthalmology 2021, 128, 813–815. [Google Scholar] [CrossRef] [PubMed]
- Burton, M.J. Corneal blindness: Prevention, treatment and rehabilitation. Community Eye Health 2009, 22, 33–35. [Google Scholar] [PubMed]
- Sridhar, M.S. Anatomy of cornea and ocular surface. Indian J. Ophthalmol. 2018, 66, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Yam, G.H.F.; Riau, A.K.; Funderburgh, M.L.; Mehta, J.S.; Jhanji, V. Keratocyte biology. Exp. Eye Res. 2020, 196, 108062. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Hertsenberg, A.J.; Funderburgh, M.L.; Burrow, M.K.; Mann, M.M.; Du, Y.; Lathrop, K.L.; Syed-Picard, F.N.; Adams, S.M.; Birk, D.E.; et al. Human limbal biopsy-derived stromal stem cells prevent corneal scarring. Sci. Transl. Med. 2014, 6, 266ra172. [Google Scholar] [CrossRef]
- Khandaker, I.; Funderburgh, J.L.; Geary, M.L.; Funderburgh, M.L.; Jhanji, V.; Du, Y.; Yam, G.H. A novel transgenic mouse model for corneal scar visualization. Exp. Eye Res. 2020, 200, 108270. [Google Scholar] [CrossRef]
- Jhanji, V.; Santra, M.; Riau, A.K.; Geary, M.L.; Yang, T.; Rubin, E.; Yusoff, N.; Dhaliwal, D.K.; Mehta, J.S.; Yam, G.H. Combined Therapy Using Human Corneal Stromal Stem Cells and Quiescent Keratocytes to Prevent Corneal Scarring after Injury. Int. J. Mol. Sci. 2022, 23, 6980. [Google Scholar] [CrossRef]
- Tuft, S.J.; Coster, D.J. The corneal endothelium. Eye 1990, 4 Pt 3, 389–424. [Google Scholar] [CrossRef]
- Chen, S.; Mienaltowski, M.J.; Birk, D.E. Regulation of corneal stroma extracellular matrix assembly. Exp. Eye Res. 2015, 133, 69–80. [Google Scholar] [CrossRef]
- Patel, S.; McLaren, J.; Hodge, D.; Bourne, W. Normal human keratocyte density and corneal thickness measurement by using confocal microscopy in vivo. Investig. Ophthalmol. Vis. Sci. 2001, 42, 333–339. [Google Scholar]
- Scott, S.G.; Jun, A.S.; Chakravarti, S. Sphere formation from corneal keratocytes and phenotype specific markers. Exp. Eye Res. 2011, 93, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.W.; Gouveia, R.M.; Connon, C.J. Low-glucose enhances keratocyte-characteristic phenotype from corneal stromal cells in serum-free conditions. Sci. Rep. 2015, 5, 10839. [Google Scholar] [CrossRef] [PubMed]
- Yam, G.H.; Yusoff, N.Z.; Kadaba, A.; Tian, D.; Myint, H.H.; Beuerman, R.W.; Zhou, L.; Mehta, J.S. Ex vivo propagation of human corneal stromal “activated keratocytes” for tissue engineering. Cell Transplant. 2015, 24, 1845–1861. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Perez, J.; Ahearne, M. Influence of Biochemical Cues in Human Corneal Stromal Cell Phenotype. Curr. Eye Res. 2019, 44, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Jester, J.V.; Moller-Pedersen, T.; Huang, J.; Sax, C.M.; Kays, W.T.; Cavangh, H.D.; Petroll, W.M.; Piatigorsky, J. The cellular basis of corneal transparency: Evidence for ‘corneal crystallins’. J. Cell Sci. 1999, 112 Pt 5, 613–622. [Google Scholar] [CrossRef] [PubMed]
- West-Mays, J.A.; Dwivedi, D.J. The keratocyte: Corneal stromal cell with variable repair phenotypes. Int. J. Biochem. Cell Biol. 2006, 38, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; Torricelli, A.A.M.; Marino, G.K. Corneal epithelial basement membrane: Structure, function and regeneration. Exp. Eye Res. 2020, 194, 108002. [Google Scholar] [CrossRef] [PubMed]
- Lam, F.W.; Phillips, J.; Landry, P.; Magadi, S.; Smith, C.W.; Rumbaut, R.E.; Burns, A.R. Platelet recruitment promotes keratocyte repopulation following corneal epithelial abrasion in the mouse. PLoS ONE 2015, 10, e0118950. [Google Scholar] [CrossRef]
- Ljubimov, A.V.; Saghizadeh, M. Progress in corneal wound healing. Prog. Retin. Eye Res. 2015, 49, 17–45. [Google Scholar] [CrossRef] [PubMed]
- Maltseva, O.; Folger, P.; Zekaria, D.; Petridou, S.; Masur, S.K. Fibroblast growth factor reversal of the corneal myofibroblast phenotype. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2490–2495. [Google Scholar]
- Karamichos, D.; Hutcheon, A.E.; Zieske, J.D. Reversal of fibrosis by TGF-beta3 in a 3D in vitro model. Exp. Eye Res. 2014, 124, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Holland, G.; Pandit, A.; Sanchez-Abella, L.; Haiek, A.; Loinaz, I.; Dupin, D.; Gonzalez, M.; Larra, E.; Bidaguren, A.; Lagali, N.; et al. Artificial Cornea: Past, Current, and Future Directions. Front. Med. 2021, 8, 770780. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R.R.; Kempuraj, D.; D’Souza, S.; Ghosh, A. Corneal stromal repair and regeneration. Prog. Retin. Eye Res. 2022, 91, 101090. [Google Scholar] [CrossRef] [PubMed]
- Whitcher, J.P.; Srinivasan, M.; Upadhyay, M.P. Corneal blindness: A global perspective. Bull. World Health Organ. 2001, 79, 214–221. [Google Scholar] [PubMed]
- Wilson, S.L.; El Haj, A.J.; Yang, Y. Control of scar tissue formation in the cornea: Strategies in clinical and corneal tissue engineering. J. Funct. Biomater. 2012, 3, 642–687. [Google Scholar] [CrossRef]
- O’Brien, K.S.; Byanju, R.; Kandel, R.P.; Poudyal, B.; Gonzales, J.A.; Porco, T.C.; Whitcher, J.P.; Srinivasan, M.; Upadhyay, M.; Lietman, T.M.; et al. Village-integrated eye workers for prevention of corneal ulcers in Nepal (VIEW study): A cluster-randomised controlled trial. Lancet Glob. Health 2022, 10, e501–e509. [Google Scholar] [CrossRef]
- de Jong, B.; van der Meulen, I.J.E.; van Vliet, J.M.J.; Lapid-Gortzak, R.; Nieuwendaal, C.P.; van den Berg, T. Effects of Corneal Scars and Their Treatment with Rigid Contact Lenses on Quality of Vision. Eye Contact Lens. 2018, 44 (Suppl. S1), S216–S220. [Google Scholar] [CrossRef]
- Tidke, S.C.; Tidake, P. A Review of Corneal Blindness: Causes and Management. Cureus 2022, 14, e30097. [Google Scholar] [CrossRef]
- Menda, S.A.; Das, M.; Panigrahi, A.; Prajna, N.V.; Acharya, N.R.; Lietman, T.M.; McLeod, S.D.; Keenan, J.D. Association of Postfungal Keratitis Corneal Scar Features with Visual Acuity. JAMA Ophthalmol. 2020, 138, 113–118. [Google Scholar] [CrossRef]
- Bachmann, B.; Taylor, R.S.; Cursiefen, C. Corneal neovascularization as a risk factor for graft failure and rejection after keratoplasty: An evidence-based meta-analysis. Ophthalmology 2010, 117, 1300–1305.e7. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, W.; Wu, J.; Zhang, H.; Zhou, H. Peripheral Ulcerative Keratitis Associated with Autoimmune Disease: Pathogenesis and Treatment. J. Ophthalmol. 2017, 2017, 7298026. [Google Scholar] [CrossRef]
- Ladas, J.G.; Mondino, B.J. Systemic disorders associated with peripheral corneal ulceration. Curr. Opin. Ophthalmol. 2000, 11, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Dua, H.S.; Ting, D.S.J.; Al Saadi, A.; Said, D.G. Chemical eye injury: Pathophysiology, assessment and management. Eye 2020, 34, 2001–2019. [Google Scholar] [CrossRef]
- Shin, Y.J.; Hyon, J.Y.; Choi, W.S.; Yi, K.; Chung, E.S.; Chung, T.Y.; Wee, W.R. Chemical injury-induced corneal opacity and neovascularization reduced by rapamycin via TGF-beta1/ERK pathways regulation. Investig. Ophthalmol. Vis. Sci. 2013, 54, 4452–4458. [Google Scholar] [CrossRef] [PubMed]
- Meller, D.; Pires, R.T.; Mack, R.J.; Figueiredo, F.; Heiligenhaus, A.; Park, W.C.; Prabhasawat, P.; John, T.; McLeod, S.D.; Steuhl, K.P.; et al. Amniotic membrane transplantation for acute chemical or thermal burns. Ophthalmology 2000, 107, 980–989, discussion 990. [Google Scholar] [CrossRef]
- Westekemper, H.; Figueiredo, F.C.; Siah, W.F.; Wagner, N.; Steuhl, K.P.; Meller, D. Clinical outcomes of amniotic membrane transplantation in the management of acute ocular chemical injury. Br. J. Ophthalmol. 2017, 101, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Culla, B.; Kolovou, P.E.; Arzeno, L.; Martinez, S.; Lopez, M.A. Boston Keratoprosthesis Type 1 in Chemical Burns. Cornea 2016, 35, 911–916. [Google Scholar] [CrossRef]
- Ang, M.; Gatinel, D.; Reinstein, D.Z.; Mertens, E.; Alio Del Barrio, J.L.; Alio, J.L. Refractive surgery beyond 2020. Eye 2021, 35, 362–382. [Google Scholar] [CrossRef]
- Vesaluoma, M.; Perez-Santonja, J.; Petroll, W.M.; Linna, T.; Alio, J.; Tervo, T. Corneal stromal changes induced by myopic LASIK. Investig. Ophthalmol. Vis. Sci. 2000, 41, 369–376. [Google Scholar]
- Netto, M.V.; Mohan, R.R.; Sinha, S.; Sharma, A.; Dupps, W.; Wilson, S.E. Stromal haze, myofibroblasts, and surface irregularity after PRK. Exp. Eye Res. 2006, 82, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, Y.S. Keratoconus. Surv. Ophthalmol. 1998, 42, 297–319. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.E.; Hayes, S.; Hardcastle, A.J.; Tuft, S.J. The pathogenesis of keratoconus. Eye 2014, 28, 189–195. [Google Scholar] [CrossRef]
- Gokhale, N.S. Epidemiology of keratoconus. Indian J. Ophthalmol. 2013, 61, 382–383. [Google Scholar] [CrossRef] [PubMed]
- Nishtala, K.; Pahuja, N.; Shetty, R.; Nuijts, R.M.; Ghosh, A. Tear biomarkers for keratoconus. Eye Vis. 2016, 3, 19. [Google Scholar] [CrossRef] [PubMed]
- Pahuja, N.; Kumar, N.R.; Shroff, R.; Shetty, R.; Nuijts, R.M.; Ghosh, A.; Sinha-Roy, A.; Chaurasia, S.S.; Mohan, R.R.; Ghosh, A. Differential Molecular Expression of Extracellular Matrix and Inflammatory Genes at the Corneal Cone Apex Drives Focal Weakening in Keratoconus. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5372–5382. [Google Scholar] [CrossRef]
- Yam, G.H.; Fuest, M.; Zhou, L.; Liu, Y.C.; Deng, L.; Chan, A.S.; Ong, H.S.; Khor, W.B.; Ang, M.; Mehta, J.S. Differential epithelial and stromal protein profiles in cone and non-cone regions of keratoconus corneas. Sci. Rep. 2019, 9, 2965. [Google Scholar] [CrossRef]
- Lerch, M.; Mainetti, C.; Terziroli Beretta-Piccoli, B.; Harr, T. Current Perspectives on Stevens-Johnson Syndrome and Toxic Epidermal Necrolysis. Clin. Rev. Allergy Immunol. 2018, 54, 147–176. [Google Scholar] [CrossRef]
- Gueudry, J.; Roujeau, J.C.; Binaghi, M.; Soubrane, G.; Muraine, M. Risk factors for the development of ocular complications of Stevens-Johnson syndrome and toxic epidermal necrolysis. Arch. Dermatol. 2009, 145, 157–162. [Google Scholar] [CrossRef]
- Klintworth, G.K. Corneal dystrophies. Orphanet J. Rare Dis. 2009, 4, 7. [Google Scholar] [CrossRef]
- NaPier, E.; Camacho, M.; McDevitt, T.F.; Sweeney, A.R. Neurotrophic keratopathy: Current challenges and future prospects. Ann. Med. 2022, 54, 666–673. [Google Scholar] [CrossRef]
- Priyadarsini, S.; Whelchel, A.; Nicholas, S.; Sharif, R.; Riaz, K.; Karamichos, D. Diabetic keratopathy: Insights and challenges. Surv. Ophthalmol. 2020, 65, 513–529. [Google Scholar] [CrossRef]
- Algarni, A.M.; Guyatt, G.H.; Turner, A.; Alamri, S. Antibiotic prophylaxis for corneal abrasion. Cochrane Database Syst. Rev. 2022, 5, CD014617. [Google Scholar] [CrossRef]
- Dang, D.H.; Riaz, K.M.; Karamichos, D. Treatment of Non-Infectious Corneal Injury: Review of Diagnostic Agents, Therapeutic Medications, and Future Targets. Drugs 2022, 82, 145–167. [Google Scholar] [CrossRef]
- Wen, F.Q.; Kohyama, T.; Skold, C.M.; Zhu, Y.K.; Liu, X.; Romberger, D.J.; Stoner, J.; Rennard, S.I. Glucocorticoids modulate TGF-beta production. Inflammation 2002, 26, 279–290. [Google Scholar] [CrossRef]
- Nien, C.J.; Flynn, K.J.; Chang, M.; Brown, D.; Jester, J.V. Reducing peak corneal haze after photorefractive keratectomy in rabbits: Prednisolone acetate 1.00% versus cyclosporine A 0.05%. J. Cataract Refract. Surg. 2011, 37, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Flach, A.J. Corneal melts associated with topically applied nonsteroidal anti-inflammatory drugs. Trans. Am. Ophthalmol. Soc. 2001, 99, 205–210. [Google Scholar] [PubMed]
- Iwamoto, S.; Koga, T.; Ohba, M.; Okuno, T.; Koike, M.; Murakami, A.; Matsuda, A.; Yokomizo, T. Non-steroidal anti-inflammatory drug delays corneal wound healing by reducing production of 12-hydroxyheptadecatrienoic acid, a ligand for leukotriene B4 receptor 2. Sci. Rep. 2017, 7, 13267. [Google Scholar] [CrossRef] [PubMed]
- McKenna, E.; Traganos, F.; Zhao, H.; Darzynkiewicz, Z. Persistent DNA damage caused by low levels of mitomycin C induces irreversible cell senescence. Cell Cycle 2012, 11, 3132–3140. [Google Scholar] [CrossRef] [PubMed]
- Kwok, S.S.; Shih, K.C.; Bu, Y.; Lo, A.C.; Chan, T.C.; Lai, J.S.; Jhanji, V.; Tong, L. Systematic Review on Therapeutic Strategies to Minimize Corneal Stromal Scarring After Injury. Eye Contact Lens. 2019, 45, 347–355. [Google Scholar] [CrossRef]
- de Oliveira, R.C.; Wilson, S.E. Fibrocytes, Wound Healing, and Corneal Fibrosis. Investig. Ophthalmol. Vis. Sci. 2020, 61, 28. [Google Scholar] [CrossRef] [PubMed]
- Kam, K.W.; Belin, M.W.; Young, A.L. Monitoring corneal densities following primary pterygium excision with adjuvant topical mitomycin-C application—An observational study of corneal scar changes. Cornea 2015, 34, 530–534. [Google Scholar] [CrossRef]
- Arranz-Marquez, E.; Katsanos, A.; Kozobolis, V.P.; Konstas, A.G.P.; Teus, M.A. A Critical Overview of the Biological Effects of Mitomycin C Application on the Cornea Following Refractive Surgery. Adv. Ther. 2019, 36, 786–797. [Google Scholar] [CrossRef]
- Shanbhag, S.S.; Singh, S.; Koshy, P.G.; Donthineni, P.R.; Basu, S. A beginner’s guide to mucous membrane grafting for lid margin keratinization: Review of indications, surgical technique and clinical outcomes. Indian J. Ophthalmol. 2021, 69, 794–805. [Google Scholar] [CrossRef] [PubMed]
- Letko, E.; Stechschulte, S.U.; Kenyon, K.R.; Sadeq, N.; Romero, T.R.; Samson, C.M.; Nguyen, Q.D.; Harper, S.L.; Primack, J.D.; Azar, D.T.; et al. Amniotic membrane inlay and overlay grafting for corneal epithelial defects and stromal ulcers. Arch. Ophthalmol. 2001, 119, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Pi, Y.L. Effect of amnion membrane transplantation on corneal neovascularization in 10 patients with alkali burn. Int. J. Ophthalmol. 2011, 4, 110–111. [Google Scholar] [CrossRef]
- Rana, D.; Desai, N.; Salave, S.; Karunakaran, B.; Giri, J.; Benival, D.; Gorantla, S.; Kommineni, N. Collagen-Based Hydrogels for the Eye: A Comprehensive Review. Gels 2023, 9, 643. [Google Scholar] [CrossRef]
- Lee, C.H.; Singla, A.; Lee, Y. Biomedical applications of collagen. Int. J. Pharm. 2001, 221, 1–22. [Google Scholar] [CrossRef]
- Robin, J.B.; Keys, C.L.; Kaminski, L.A.; Viana, M.A. The effect of collagen shields on rabbit corneal reepithelialization after chemical debridement. Investig. Ophthalmol. Vis. Sci. 1990, 31, 1294–1300. [Google Scholar]
- Jester, J.V.; Budge, A.; Fisher, S.; Huang, J. Corneal keratocytes: Phenotypic and species differences in abundant protein expression and in vitro light-scattering. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2369–2378. [Google Scholar] [CrossRef] [PubMed]
- Fenner, B.J.; Yusoff, N.; Fuest, M.; Zhou, L.; Bandeira, F.; Cajucom-Uy, H.Y.; Tan, H.K.; Mehta, J.S.; Yam, G.H.F. A cellular and proteomic approach to assess proteins extracted from cryopreserved human amnion in the cultivation of corneal stromal keratocytes for stromal cell therapy. Eye Vis. 2019, 6, 30. [Google Scholar] [CrossRef] [PubMed]
- Yam, G.H.; Fuest, M.; Yusoff, N.; Goh, T.W.; Bandeira, F.; Setiawan, M.; Seah, X.Y.; Lwin, N.C.; Stanzel, T.P.; Ong, H.S.; et al. Safety and Feasibility of Intrastromal Injection of Cultivated Human Corneal Stromal Keratocytes as Cell-Based Therapy for Corneal Opacities. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3340–3354. [Google Scholar] [CrossRef] [PubMed]
- Arnalich-Montiel, F.; Pastor, S.; Blazquez-Martinez, A.; Fernandez-Delgado, J.; Nistal, M.; Alio, J.L.; De Miguel, M.P. Adipose-derived stem cells are a source for cell therapy of the corneal stroma. Stem Cells 2008, 26, 570–579. [Google Scholar] [CrossRef]
- Du, Y.; Carlson, E.C.; Funderburgh, M.L.; Birk, D.E.; Pearlman, E.; Guo, N.; Kao, W.W.; Funderburgh, J.L. Stem cell therapy restores transparency to defective murine corneas. Stem Cells 2009, 27, 1635–1642. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, J.; Liu, C.Y.; Wang, I.J.; Sieber, M.; Chang, J.; Jester, J.V.; Kao, W.W. Cell therapy of congenital corneal diseases with umbilical mesenchymal stem cells: Lumican null mice. PLoS ONE 2010, 5, e10707. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, J.; Liu, C.Y.; Hayashi, Y.; Kao, W.W. Bone marrow mesenchymal stem cells can differentiate and assume corneal keratocyte phenotype. J. Cell. Mol. Med. 2012, 16, 1114–1124. [Google Scholar] [CrossRef] [PubMed]
- Coulson-Thomas, V.J.; Caterson, B.; Kao, W.W. Transplantation of human umbilical mesenchymal stem cells cures the corneal defects of mucopolysaccharidosis VII mice. Stem Cells 2013, 31, 2116–2126. [Google Scholar] [CrossRef]
- Zhang, S.; Espandar, L.; Imhof, K.M.; Bunnell, B.A. Differentiation of Human Adipose-derived Stem Cells along the Keratocyte Lineage In vitro. J. Clin. Exp. Ophthalmol. 2013, 4, 11435. [Google Scholar] [CrossRef]
- Alio del Barrio, J.L.; Chiesa, M.; Garagorri, N.; Garcia-Urquia, N.; Fernandez-Delgado, J.; Bataille, L.; Rodriguez, A.; Arnalich-Montiel, F.; Zarnowski, T.; Alvarez de Toledo, J.P.; et al. Acellular human corneal matrix sheets seeded with human adipose-derived mesenchymal stem cells integrate functionally in an experimental animal model. Exp. Eye Res. 2015, 132, 91–100. [Google Scholar] [CrossRef]
- Alio Del Barrio, J.L.; El Zarif, M.; Azaar, A.; Makdissy, N.; Khalil, C.; Harb, W.; El Achkar, I.; Jawad, Z.A.; de Miguel, M.P.; Alio, J.L. Corneal Stroma Enhancement with Decellularized Stromal Laminas with or Without Stem Cell Recellularization for Advanced Keratoconus. Am. J. Ophthalmol. 2018, 186, 47–58. [Google Scholar] [CrossRef]
- Hass, R.; Kasper, C.; Bohm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Branch, M.J.; Hashmani, K.; Dhillon, P.; Jones, D.R.; Dua, H.S.; Hopkinson, A. Mesenchymal stem cells in the human corneal limbal stroma. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5109–5116. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, K.W.; Chun, Y.S.; Kim, J.C. Human mesenchymal stem cells differentiate into keratocyte-like cells in keratocyte-conditioned medium. Exp. Eye Res. 2012, 101, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.; Balayan, A.; Funderburgh, M.L.; Ngo, J.; Funderburgh, J.L.; Deng, S.X. Differentiation Capacity of Human Mesenchymal Stem Cells into Keratocyte Lineage. Investig. Ophthalmol. Vis. Sci. 2019, 60, 3013–3023. [Google Scholar] [CrossRef] [PubMed]
- Kao, W.W.; Coulson-Thomas, V.J. Cell Therapy of Corneal Diseases. Cornea 2016, 35 (Suppl. S1), S9–S19. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Liu, G.; Meng, F.; Wang, W.; Hao, P.; Xiang, Y.; Wang, Y.; Han, R.; Li, F.; Wang, L.; et al. Paracrine effects of mesenchymal stem cells on the activation of keratocytes. Br. J. Ophthalmol. 2017, 101, 1583–1590. [Google Scholar] [CrossRef]
- Fuentes-Julian, S.; Arnalich-Montiel, F.; Jaumandreu, L.; Leal, M.; Casado, A.; Garcia-Tunon, I.; Hernandez-Jimenez, E.; Lopez-Collazo, E.; De Miguel, M.P. Adipose-derived mesenchymal stem cell administration does not improve corneal graft survival outcome. PLoS ONE 2015, 10, e0117945. [Google Scholar] [CrossRef]
- Alio Del Barrio, J.L.; El Zarif, M.; de Miguel, M.P.; Azaar, A.; Makdissy, N.; Harb, W.; El Achkar, I.; Arnalich-Montiel, F.; Alio, J.L. Cellular Therapy with Human Autologous Adipose-Derived Adult Stem Cells for Advanced Keratoconus. Cornea 2017, 36, 952–960. [Google Scholar] [CrossRef]
- Alio, J.L.; Alio Del Barrio, J.L.; El Zarif, M.; Azaar, A.; Makdissy, N.; Khalil, C.; Harb, W.; El Achkar, I.; Jawad, Z.A.; de Miguel, M.P. Regenerative Surgery of the Corneal Stroma for Advanced Keratoconus: One Year Outcomes. Am. J. Ophthalmol. 2019, 203, 53–68. [Google Scholar] [CrossRef]
- El Zarif, M.; Alio Del Barrio, J.L.; Mingo, D.; Jawad, K.A.; Alio, J.L. Corneal Stromal Densitometry Evolution in a Clinical Model of Cellular Therapy for Advanced Keratoconus. Cornea 2023, 42, 332–343. [Google Scholar] [CrossRef]
- Du, Y.; Funderburgh, M.L.; Mann, M.M.; SundarRaj, N.; Funderburgh, J.L. Multipotent stem cells in human corneal stroma. Stem Cells 2005, 23, 1266–1275. [Google Scholar] [CrossRef]
- Funderburgh, J.L.; Funderburgh, M.L.; Du, Y. Stem Cells in the Limbal Stroma. Ocul. Surf. 2016, 14, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.H.; Tian, Y.; Funderburgh, J.; Pellegrini, G.; Zhang, K.; Goldberg, J.L.; Ali, R.R.; Young, M.; Xie, Y.; Temple, S. Regenerating Eye Tissues to Preserve and Restore Vision. Cell Stem Cell 2018, 23, 453. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Yun, H.; Funderburgh, M.L.; Du, Y. Regenerative therapy for the Cornea. Prog. Retin. Eye Res. 2022, 87, 101011. [Google Scholar] [CrossRef] [PubMed]
- Yam, G.H.; Pi, S.; Du, Y.; Mehta, J.S. Posterior corneoscleral limbus: Architecture, stem cells, and clinical implications. Prog. Retin. Eye Res. 2023, 96, 101192. [Google Scholar] [CrossRef] [PubMed]
- Ghoubay, D.; Borderie, M.; Grieve, K.; Martos, R.; Bocheux, R.; Nguyen, T.M.; Callard, P.; Chedotal, A.; Borderie, V.M. Corneal stromal stem cells restore transparency after N2 injury in mice. Stem Cells Transl. Med. 2020, 9, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Hertsenberg, A.J.; Shojaati, G.; Funderburgh, M.L.; Mann, M.M.; Du, Y.; Funderburgh, J.L. Corneal stromal stem cells reduce corneal scarring by mediating neutrophil infiltration after wounding. PLoS ONE 2017, 12, e0171712. [Google Scholar] [CrossRef] [PubMed]
- Santra, M.; Geary, M.L.; Rubin, E.; Hsu, M.Y.S.; Funderburgh, M.L.; Chandran, C.; Du, Y.; Dhaliwal, D.K.; Jhanji, V.; Yam, G.H. Good manufacturing practice production of human corneal limbus-derived stromal stem cells and in vitro quality screening for therapeutic inhibition of corneal scarring. Stem Cell Res. Ther. 2024, 15, 11. [Google Scholar] [CrossRef]
- Zhang, Y.; Bi, J.; Huang, J.; Tang, Y.; Du, S.; Li, P. Exosome: A Review of Its Classification, Isolation Techniques, Storage, Diagnostic and Targeted Therapy Applications. Int. J. Nanomed. 2020, 15, 6917–6934. [Google Scholar] [CrossRef]
- Bruno, S.; Grange, C.; Deregibus, M.C.; Calogero, R.A.; Saviozzi, S.; Collino, F.; Morando, L.; Busca, A.; Falda, M.; Bussolati, B.; et al. Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J. Am. Soc. Nephrol. 2009, 20, 1053–1067. [Google Scholar] [CrossRef]
- Lai, R.C.; Arslan, F.; Lee, M.M.; Sze, N.S.; Choo, A.; Chen, T.S.; Salto-Tellez, M.; Timmers, L.; Lee, C.N.; El Oakley, R.M.; et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010, 4, 214–222. [Google Scholar] [CrossRef]
- Doeppner, T.R.; Herz, J.; Gorgens, A.; Schlechter, J.; Ludwig, A.K.; Radtke, S.; de Miroschedji, K.; Horn, P.A.; Giebel, B.; Hermann, D.M. Extracellular Vesicles Improve Post-Stroke Neuroregeneration and Prevent Postischemic Immunosuppression. Stem Cells Transl. Med. 2015, 4, 1131–1143. [Google Scholar] [CrossRef]
- Nikfarjam, S.; Rezaie, J.; Zolbanin, N.M.; Jafari, R. Mesenchymal stem cell derived-exosomes: A modern approach in translational medicine. J. Transl. Med. 2020, 18, 449. [Google Scholar] [CrossRef]
- Tang, Q.; Lu, B.; He, J.; Chen, X.; Fu, Q.; Han, H.; Luo, C.; Yin, H.; Qin, Z.; Lyu, D.; et al. Exosomes-loaded thermosensitive hydrogels for corneal epithelium and stroma regeneration. Biomaterials 2022, 280, 121320. [Google Scholar] [CrossRef]
- Bai, L.; Shao, H.; Wang, H.; Zhang, Z.; Su, C.; Dong, L.; Yu, B.; Chen, X.; Li, X.; Zhang, X. Effects of Mesenchymal Stem Cell-Derived Exosomes on Experimental Autoimmune Uveitis. Sci. Rep. 2017, 7, 4323. [Google Scholar] [CrossRef]
- Nuzzi, R.; Caselgrandi, P.; Vercelli, A. Effect of Mesenchymal Stem Cell-Derived Exosomes on Retinal Injury: A Review of Current Findings. Stem Cells Int. 2020, 2020, 8883616. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Zheng, Q.Q.; Shen, J.; Li, Q.S.; Song, X.H.; Luo, H.B.; Hong, C.Y.; Yao, K. Effects of Adipose-derived Mesenchymal Stem Cell Exosomes on Corneal Stromal Fibroblast Viability and Extracellular Matrix Synthesis. Chin. Med. J. 2018, 131, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Baradaran-Rafii, A.; Eslani, M.; Haq, Z.; Shirzadeh, E.; Huvard, M.J.; Djalilian, A.R. Current and Upcoming Therapies for Ocular Surface Chemical Injuries. Ocul. Surf. 2017, 15, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Ong, H.S.; Riau, A.K.; Yam, G.H.; Yusoff, N.; Han, E.J.Y.; Goh, T.W.; Lai, R.C.; Lim, S.K.; Mehta, J.S. Mesenchymal Stem Cell Exosomes as Immunomodulatory Therapy for Corneal Scarring. Int. J. Mol. Sci. 2023, 24, 7456. [Google Scholar] [CrossRef]
- Shojaati, G.; Khandaker, I.; Funderburgh, M.L.; Mann, M.M.; Basu, R.; Stolz, D.B.; Geary, M.L.; Dos Santos, A.; Deng, S.X.; Funderburgh, J.L. Mesenchymal Stem Cells Reduce Corneal Fibrosis and Inflammation via Extracellular Vesicle-Mediated Delivery of miRNA. Stem Cells Transl. Med. 2019, 8, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Yam, G.H.; Yang, T.; Geary, M.L.; Santra, M.; Funderburgh, M.; Rubin, E.; Du, Y.; Sahel, J.A.; Jhanji, V.; Funderburgh, J.L. Human corneal stromal stem cells express anti-fibrotic microRNA-29a and 381-5p—A robust cell selection tool for stem cell therapy of corneal scarring. J. Adv. Res. 2023, 45, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Kular, J.K.; Basu, S.; Sharma, R.I. The extracellular matrix: Structure, composition, age-related differences, tools for analysis and applications for tissue engineering. J. Tissue Eng. 2014, 5, 2041731414557112. [Google Scholar] [CrossRef]
- Yin, H.; Lu, Q.; Wang, X.; Majumdar, S.; Jun, A.S.; Stark, W.J.; Grant, M.P.; Elisseeff, J.H. Tissue-derived microparticles reduce inflammation and fibrosis in cornea wounds. Acta Biomater. 2019, 85, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Rothe, R.; Voigt, D.; Sayed, A.; Huang, C.; Hauser, S.; Lee, P.W.; Cui, M.; Saenz, J.P.; Boccaccini, A.R.; et al. A self-assembled dynamic extracellular matrix-like hydrogel system with multi-scale structures for cell bioengineering applications. Acta Biomater. 2023, 162, 211–225. [Google Scholar] [CrossRef]
- Luo, Y.; Shen, M.; Feng, P.; Qiu, H.; Wu, X.; Yang, L.; Zhu, Y. Various administration forms of decellularized amniotic membrane extract towards improving corneal repair. J. Mater. Chem. B 2021, 9, 9347–9357. [Google Scholar] [CrossRef]
- Sullivan, M.M.; Sage, E.H. Hevin/SC1, a matricellular glycoprotein and potential tumor-suppressor of the SPARC/BM-40/Osteonectin family. Int. J. Biochem. Cell Biol. 2004, 36, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, S.S.; Perera, P.R.; Poh, R.; Lim, R.R.; Wong, T.T.; Mehta, J.S. Hevin plays a pivotal role in corneal wound healing. PLoS ONE 2013, 2013, e81544. [Google Scholar] [CrossRef]
- McConnell, B.B.; Ghaleb, A.M.; Nandan, M.O.; Yang, V.W. The diverse functions of Kruppel-like factors 4 and 5 in epithelial biology and pathobiology. BioEssays 2007, 29, 549–557. [Google Scholar] [CrossRef]
- Tiwari, A.; Loughner, C.L.; Swamynathan, S.; Swamynathan, S.K. KLF4 Plays an Essential Role in Corneal Epithelial Homeostasis by Promoting Epithelial Cell Fate and Suppressing Epithelial-Mesenchymal Transition. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2785–2795. [Google Scholar] [CrossRef]
- Fujimoto, S.; Hayashi, R.; Hara, S.; Sasamoto, Y.; Harrington, J.; Tsujikawa, M.; Nishida, K. KLF4 prevents epithelial to mesenchymal transition in human corneal epithelial cells via endogenous TGF-beta2 suppression. Regen. Ther. 2019, 11, 249–257. [Google Scholar] [CrossRef]
- Tarczewska, A.; Greb-Markiewicz, B. The Significance of the Intrinsically Disordered Regions for the Functions of the bHLH Transcription Factors. Int. J. Mol. Sci. 2019, 20, 5306. [Google Scholar] [CrossRef]
- Gupta, S.; Fink, M.K.; Kempuraj, D.; Sinha, N.R.; Martin, L.M.; Keele, L.M.; Sinha, P.R.; Giuliano, E.A.; Hesemann, N.P.; Raikwar, S.P.; et al. Corneal fibrosis abrogation by a localized AAV-mediated inhibitor of differentiation 3 (Id3) gene therapy in rabbit eyes in vivo. Mol. Ther. J. Am. Soc. Gene Ther. 2022, 30, 3257–3269. [Google Scholar] [CrossRef]
- Yan, X.; Liao, H.; Cheng, M.; Shi, X.; Lin, X.; Feng, X.H.; Chen, Y.G. Smad7 Protein Interacts with Receptor-regulated Smads (R-Smads) to Inhibit Transforming Growth Factor-beta (TGF-beta)/Smad Signaling. J. Biol. Chem. 2016, 291, 382–392. [Google Scholar] [CrossRef]
- Hu, H.H.; Chen, D.Q.; Wang, Y.N.; Feng, Y.L.; Cao, G.; Vaziri, N.D.; Zhao, Y.Y. New insights into TGF-beta/Smad signaling in tissue fibrosis. Chem. Biol. Interact. 2018, 292, 76–83. [Google Scholar] [CrossRef]
- Gupta, S.; Rodier, J.T.; Sharma, A.; Giuliano, E.A.; Sinha, P.R.; Hesemann, N.P.; Ghosh, A.; Mohan, R.R. Targeted AAV5-Smad7 gene therapy inhibits corneal scarring in vivo. PLoS ONE 2017, 12, e0172928. [Google Scholar] [CrossRef]
- Chung, J.K.; Park, S.A.; Hwang, H.S.; Kim, K.S.; Cho, Y.J.; You, Y.S.; Kim, Y.S.; Jang, J.W.; Lee, S.J. Effects of exogenous recombinant human bone morphogenic protein-7 on the corneal epithelial mesenchymal transition and fibrosis. Int. J. Ophthalmol. 2017, 10, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Saika, S.; Ikeda, K.; Yamanaka, O.; Flanders, K.C.; Nakajima, Y.; Miyamoto, T.; Ohnishi, Y.; Kao, W.W.; Muragaki, Y.; Ooshima, A. Therapeutic effects of adenoviral gene transfer of bone morphogenic protein-7 on a corneal alkali injury model in mice. Lab. Investig. 2005, 85, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Tandon, A.; Sharma, A.; Rodier, J.T.; Klibanov, A.M.; Rieger, F.G.; Mohan, R.R. BMP7 gene transfer via gold nanoparticles into stroma inhibits corneal fibrosis in vivo. PLoS ONE 2013, 8, e66434. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R.R.; Tovey, J.C.; Gupta, R.; Sharma, A.; Tandon, A. Decorin biology, expression, function and therapy in the cornea. Curr. Mol. Med. 2011, 11, 110–128. [Google Scholar] [CrossRef]
- Hill, L.J.; Moakes, R.J.A.; Vareechon, C.; Butt, G.; Ng, A.; Brock, K.; Chouhan, G.; Vincent, R.C.; Abbondante, S.; Williams, R.L.; et al. Sustained release of decorin to the surface of the eye enables scarless corneal regeneration. NPJ Regen. Med. 2018, 3, 23. [Google Scholar] [CrossRef]
- Grisanti, S.; Szurman, P.; Warga, M.; Kaczmarek, R.; Ziemssen, F.; Tatar, O.; Bartz-Schmidt, K.U. Decorin modulates wound healing in experimental glaucoma filtration surgery: A pilot study. Investig. Ophthalmol. Vis. Sci. 2005, 46, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Jhanji, V.; Billig, I.; Yam, G.H. Cell-Free Biological Approach for Corneal Stromal Wound Healing. Front. Pharmacol. 2021, 12, 671405. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Mehan, M.M.; Sinha, S.; Cowden, J.W.; Mohan, R.R. Trichostatin a inhibits corneal haze in vitro and in vivo. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2695–2701. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, K.S.; Giuliano, E.A.; Sharm, A.; Mohan, R.R. Suberoylanilide hydroxamic acid (vorinostat): Its role on equine corneal fibrosis and matrix metalloproteinase activity. Vet. Ophthalmol. 2014, 17 (Suppl. S1), 61–68. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, R.W.D.; Vickaryous, M.K.; Viloria-Petit, A.M. Signalling by Transforming Growth Factor Beta Isoforms in Wound Healing and Tissue Regeneration. J. Dev. Biol. 2016, 4, 21. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.; Funderburgh, J.; Khandaker, I.; Geary, M.L.; Basu, R.; Funderburgh, M.L.; Du, Y.; Yam, G.H. The anti-scarring effect of corneal stromal stem cell therapy is mediated by transforming growth factor b3. Eye Vis. 2020, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Karamichos, D.; Hutcheon, A.E.; Zieske, J.D. Transforming growth factor-beta3 regulates assembly of a non-fibrotic matrix in a 3D corneal model. J. Tissue Eng. Regen. Med. 2011, 5, e228–e238. [Google Scholar] [CrossRef]
- Karamichos, D.; Rich, C.B.; Zareian, R.; Hutcheon, A.E.; Ruberti, J.W.; Trinkaus-Randall, V.; Zieske, J.D. TGF-beta3 stimulates stromal matrix assembly by human corneal keratocyte-like cells. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6612–6619. [Google Scholar] [CrossRef]
- Satish, L.; Kathju, S. Cellular and Molecular Characteristics of Scarless versus Fibrotic Wound Healing. Dermatol. Res. Pract. 2010, 2010, 790234. [Google Scholar] [CrossRef]
- Schrementi, M.E.; Ferreira, A.M.; Zender, C.; DiPietro, L.A. Site-specific production of TGF-beta in oral mucosal and cutaneous wounds. Wound Repair Regen. 2008, 16, 80–86. [Google Scholar] [CrossRef]
- Rollins, B.J.; O’Connell, T.M.; Bennett, G.; Burton, L.E.; Stiles, C.D.; Rheinwald, J.G. Environment-dependent growth inhibition of human epidermal keratinocytes by recombinant human transforming growth factor-beta. J. Cell. Physiol. 1989, 139, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.J.; Nguyen, D.D.; Lai, J.Y. Poly(l-Histidine)-Mediated On-Demand Therapeutic Delivery of Roughened Ceria Nanocages for Treatment of Chemical Eye Injury. Adv. Sci. 2023, 10, e2302174. [Google Scholar] [CrossRef]
- Sampaio, L.P.; Villabona-Martinez, V.; Shiju, T.M.; Santhiago, M.R.; Wilson, S.E. Topical Losartan Decreases Myofibroblast Generation But Not Corneal Opacity After Surface Blast-Simulating Irregular PTK in Rabbits. Transl. Vis. Sci. Technol. 2023, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, L.P.; Hilgert, G.S.L.; Shiju, T.M.; Santhiago, M.R.; Wilson, S.E. Topical Losartan and corticosteroid additively inhibit corneal stromal myofibroblast generation and scarring fibrosis after alkali burn injury. Transl. Vis. Sci. Technol. 2022, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Souza, A.L.; Ambrosio, R., Jr.; Bandeira, F.; Salomao, M.Q.; Souza Lima, A.; Wilson, S.E. Topical Losartan for treating corneal fibrosis (haze): First clinical experience. J. Refract. Surg. 2022, 38, 741–746. [Google Scholar] [CrossRef]
- Okayama, K.; Azuma, J.; Dosaka, N.; Iekushi, K.; Sanada, F.; Kusunoki, H.; Iwabayashi, M.; Rakugi, H.; Taniyama, Y.; Morishita, R. Hepatocyte growth factor reduces cardiac fibrosis by inhibiting endothelial-mesenchymal transition. Hypertension 2012, 59, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, H.; Thomasy, S.M.; Russell, P.; Murphy, C.J. The role of hepatocyte growth factor in corneal wound healing. Exp. Eye Res. 2018, 166, 49–55. [Google Scholar] [CrossRef]
- Shukla, M.N.; Rose, J.L.; Ray, R.; Lathrop, K.L.; Ray, A.; Ray, P. Hepatocyte growth factor inhibits epithelial to myofibroblast transition in lung cells via Smad7. Am. J. Respir. Cell Mol. Biol. 2009, 40, 643–653. [Google Scholar] [CrossRef]
- Mizuno, S.; Matsumoto, K.; Li, M.Y.; Nakamura, T. HGF reduces advancing lung fibrosis in mice: A potential role for MMP-dependent myofibroblast apoptosis. FASEB J. 2005, 19, 580–582. [Google Scholar] [CrossRef]
- Omoto, M.; Suri, K.; Amouzegar, A.; Li, M.; Katikireddy, K.R.; Mittal, S.K.; Chauhan, S.K. Hepatocyte growth factor suppresses inflammation and promotes epithelium repair in corneal injury. Mol. Ther. 2017, 25, 1881–1888. [Google Scholar] [CrossRef]
- Mittal, S.K.; Omoto, M.; Amouzegar, A.; Sahu, A.; Rezazadeh, A.; Katikireddy, K.R.; Shah, D.I.; Sahu, S.K.; Chauhan, S.K. Restoration of corneal transparency by mesenchymal stem cells. Stem Cell Rep. 2016, 7, 583–590. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, R.C.; Murillo, S.; Saikia, P.; Wilson, S.E. The efficacy of topical HGF on corneal fibrosis and epithelial healing after scar-producing PRK injury in rabbits. Transl. Vis. Sci. Technol. 2020, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Kao, W.W.; Liu, C.Y. Roles of lumican and keratocan on corneal transparency. Glycoconj. J. 2002, 19, 275–285. [Google Scholar] [CrossRef]
- Vij, N.; Roberts, L.; Joyce, S.; Chakravarti, S. Lumican regulates corneal inflammatory responses by modulating Fas-Fas ligand signaling. Investig. Ophthalmol. Vis. Sci. 2005, 46, 88–95. [Google Scholar] [CrossRef]
- Gesteira, T.F.; Coulson-Thomas, V.J.; Yuan, Y.; Zhang, J.; Nader, H.B.; Kao, W.W. Lumican peptides: Rational design targeting ALK5/TGFΒRI. Sci. Rep. 2017, 7, 42057. [Google Scholar] [CrossRef] [PubMed]
- Kao, W.; Zhang, J.; Yasuda, S.; Yuan, Y.; Saika, S.; Liu, C.Y. Role of lumican on TgfβR signaling in the healing of deep corneal epithelial debridement. Investig. Ophthalmol. Vis. Sci. 2023, 64, 506. [Google Scholar]
- Roth, J.; Yam, G.H.; Fan, J.; Hirano, K.; Gaplovska-Kysela, K.; Le Fourn, V.; Guhl, B.; Santimaria, R.; Torossi, T.; Ziak, M.; et al. Protein quality control: The who’s who, the where’s and therapeutic escapes. Histochem. Cell Biol. 2008, 129, 163–177. [Google Scholar] [CrossRef]
- Morishige, N.; Ko, J.A.; Morita, Y.; Nishida, T. Expression of semaphorin 3A in the rat corneal epithelium during wound healing. Biochem. Biophys. Res. Commun. 2010, 395, 451–457. [Google Scholar] [CrossRef]
- Jeon, K.I.; Nehrke, K.; Huxlin, K.R. Semaphorin 3A potentiates the profibrotic effects of transforming growth factor-beta1 in the cornea. Biochem. Biophys. Res. Commun. 2020, 521, 333–339. [Google Scholar] [CrossRef]
- Takayama, K.I.; Suzuki, T.; Fujimura, T.; Takahashi, S.; Inoue, S. Association of USP10 with G3BP2 inhibits p53 signaling and contributes to poor outcome in prostate cancer. Mol. Cancer Res. MCR 2018, 16, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, S.R.; Tedesco, L.J.; Wang, L.; Bernstein, A.M. The deubiquitylase USP10 regulates integrin beta1 and beta5 and fibrotic wound healing. J. Cell Sci. 2017, 130, 3481–3495. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garcia, M.T.; Cidad, P.; Lopez-Lopez, J.R. The secret life of ion channels: Kv1.3 potassium channels and proliferation. Am. J. Physiol. Cell Physiol. 2018, 314, C27–C42. [Google Scholar] [CrossRef]
- Girault, A.; Chebli, J.; Prive, A.; Trinh, N.T.; Maille, E.; Grygorczyk, R.; Brochiero, E. Complementary roles of KCa3.1 channels and beta1-integrin during alveolar epithelial repair. Respir. Res. 2015, 16, 100. [Google Scholar] [CrossRef] [PubMed]
- Anumanthan, G.; Gupta, S.; Fink, M.K.; Hesemann, N.P.; Bowles, D.K.; McDaniel, L.M.; Muhammad, M.; Mohan, R.R. KCa3.1 ion channel: A novel therapeutic target for corneal fibrosis. PLoS ONE 2018, 13, e0192145. [Google Scholar] [CrossRef] [PubMed]
- Sriram, S.; Gibson, D.J.; Robinson, P.; Pi, L.; Tuli, S.; Lewin, A.S.; Schultz, G. Assessment of anti-scarring therapies in ex vivo organ cultured rabbit corneas. Exp. Eye Res. 2014, 125, 173–182. [Google Scholar] [CrossRef]
- Gain, P.; Jullienne, R.; He, Z.; Aldossary, M.; Acquart, S.; Cognasse, F.; Thuret, G. Global survey of corneal transplantation and eye banking. JAMA Ophthalmol. 2016, 134, 167–173. [Google Scholar] [CrossRef]
- Santra, M.; Liu, Y.C.; Jhanji, V.; Yam, G.H. Human SMILE-derived stromal lenticule scaffold for regenerative therapy: Review and perspectives. Int. J. Mol. Sci. 2022, 23, 7967. [Google Scholar] [CrossRef]
- Pradhan, K.R.; Reinstein, D.Z.; Carp, G.I.; Archer, T.J.; Gobbe, M.; Gurung, R. Femtosecond laser-assisted keyhole endokeratophakia: Correction of hyperopia by implantation of an allogeneic lenticule obtained by SMILE from a myopic donor. J. Refract. Surg. 2013, 29, 777–782. [Google Scholar] [CrossRef]
- Li, M.; Zhao, F.; Li, M.; Knorz, M.C.; Zhou, X. Treatment of corneal ectasia by implantation of an allogenic corneal lenticule. J. Refract. Surg. 2018, 34, 347–350. [Google Scholar] [CrossRef]
- Doroodgar, F.; Jabbarvand, M.; Niazi, S.; Karimian, F.; Niazi, F.; Sanginabadi, A.; Ghoreishi, M.; Alinia, C.; Hashemi, H.; Alio, J.L. Customized Stromal Lenticule Implantation for Keratoconus. J. Refract. Surg. 2020, 36, 786–794. [Google Scholar] [CrossRef]
- Damgaard, I.B.; Riau, A.K.; Liu, Y.C.; Tey, M.L.; Yam, G.H.; Mehta, J.S. Reshaping and customization of SMILE-derived biological lenticules for intrastromal implantation. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2555–2563. [Google Scholar] [CrossRef] [PubMed]
- Yam, G.H.; Yusoff, N.Z.; Goh, T.W.; Setiawan, M.; Lee, X.W.; Liu, Y.C.; Mehta, J.S. Decellularization of human stromal refractive lenticules for corneal tissue engineering. Sci. Rep. 2016, 6, 26339. [Google Scholar] [CrossRef] [PubMed]
- Riau, A.K.; Liu, Y.C.; Yam, G.H.F.; Mehta, J.S. Stromal keratophakia: Corneal inlay implantation. Prog. Retin. Eye Res. 2020, 75, 100780. [Google Scholar] [CrossRef] [PubMed]
- Yam, G.H.; Bandeira, F.; Liu, Y.C.; Devarajan, K.; Yusoff, N.; Htoon, H.M.; Mehta, J.S. Effect of corneal stromal lenticule customization on neurite distribution and excitatory property. J. Adv. Res. 2022, 38, 275–284. [Google Scholar] [CrossRef]
- Mimura, T.; Amano, S.; Yokoo, S.; Uchida, S.; Yamagami, S.; Usui, T.; Kimura, Y.; Tabata, Y. Tissue engineering of corneal stroma with rabbit fibroblast precursors and gelatin hydrogels. Mol. Vis. 2008, 14, 1819–1828. [Google Scholar] [PubMed]
- Goodarzi, H.; Jadidi, K.; Pourmotabed, S.; Sharifi, E.; Aghamollaei, H. Preparation and in vitro characterization of cross-linked collagen-gelatin hydrogel using EDC/NHS for corneal tissue engineering applications. Int. J. Biol. Macromol. 2019, 126, 620–632. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, Q.; Li, Q.; Kawazoe, N.; Chen, G. Functional hydrogels with tunable structures and properties for tissue engineering applications. Front. Chem. 2018, 6, 499. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Koole, L.H.; Gao, C.; Yang, D.; Yang, L.; Zhang, C.; Li, H. The potential utility of hybrid photo-crosslinked hydrogels with non-immunogenic component for cartilage repair. NPJ Regen. Med. 2021, 6, 54. [Google Scholar] [CrossRef]
- Nagaraj, A.; Etxeberria, A.E.; Naffa, R.; Zidan, G.; Seyfoddin, A. 3D-printed hybrid collagen/GelMA hydrogels for tissue engineering applications. Biology 2022, 11, 1561. [Google Scholar] [CrossRef]
- Huang, J.; Jiang, T.; Li, J.; Qie, J.; Cheng, X.; Wang, Y.; Zhou, T.; Liu, J.; Han, H.; Yao, K.; et al. Biomimetic corneal stroma for scarless corneal wound healing via structural restoration and microenvironment modulation. Adv. Health Mater. 2024, 13, e2302889. [Google Scholar] [CrossRef]
- Bray, L.J.; George, K.A.; Hutmacher, D.W.; Chirila, T.V.; Harkin, D.G. A dual-layer silk fibroin scaffold for reconstructing the human corneal limbus. Biomaterials 2012, 33, 3529–3538. [Google Scholar] [CrossRef]
- Ghezzi, C.E.; Marelli, B.; Omenetto, F.G.; Funderburgh, J.L.; Kaplan, D.L. 3D functional corneal stromal tissue equivalent based on corneal stromal stem cells and multi-layered silk film architecture. PLoS ONE 2017, 12, e0169504. [Google Scholar] [CrossRef]
- Wang, L.; Ma, R.; Du, G.; Guo, H.; Huang, Y. Biocompatibility of helicoidal multilamellar arginine-glycine-aspartic acid-functionalized silk biomaterials in a rabbit corneal model. J. Biomed. Mater. Res. B Appl. Biomater. 2015, 103, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Raia, N.R.; Bakaysa, S.L.; Ghezzi, C.E.; House, M.D.; Kaplan, D.L. Ex vivo pregnant-like tissue model to assess injectable hydrogel for preterm birth prevention. J. Biomed. Mater. Res. B Appl. Biomater. 2020, 108, 468–474. [Google Scholar] [CrossRef]
- Aranaz, I.; Alcantara, A.R.; Civera, M.C.; Arias, C.; Elorza, B.; Heras Caballero, A.; Acosta, N. Chitosan: An overview of its properties and applications. Polymers 2021, 13, 3256. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Wang, I.J.; Hu, F.R.; Young, T.H. Applications of biomaterials in corneal endothelial tissue engineering. Cornea 2016, 35 (Suppl. S1), S25–S30. [Google Scholar] [CrossRef]
- Guan, L.; Tian, P.; Ge, H.; Tang, X.; Zhang, H.; Du, L.; Liu, P. Chitosan-functionalized silk fibroin 3D scaffold for keratocyte culture. J. Mol. Histol. 2013, 44, 609–618. [Google Scholar] [CrossRef]
- Li, Y.H.; Cheng, C.Y.; Wang, N.K.; Tan, H.Y.; Tsai, Y.J.; Hsiao, C.H.; Ma, D.H.; Yeh, L.K. Characterization of the modified chitosan membrane cross-linked with genipin for the cultured corneal epithelial cells. Colloids Surf. B Biointerfaces 2015, 126, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Zheng, Q.; Luo, H.; Li, X.; Chen, Z.; Song, Z.; Zhou, G.; Hong, C. Exosomal miR-19a from adipose-derived stem cells suppresses differentiation of corneal keratocytes into myofibroblasts. Aging 2020, 12, 4093–4110. [Google Scholar] [CrossRef]
- Al Halawani, A.; Mithieux, S.M.; Yeo, G.C.; Hosseini-Beheshti, E.; Weiss, A.S. Extracellular vesicles: Interplay with the extracellular matrix and modulated cell responses. Int. J. Mol. Sci. 2022, 23, 3389. [Google Scholar] [CrossRef] [PubMed]
- Hogan, K.J.; Perez, M.R.; Mikos, A.G. Extracellular matrix component-derived nanoparticles for drug delivery and tissue engineering. J. Control. Release 2023, 360, 888–912. [Google Scholar] [CrossRef] [PubMed]
- Calonge, M.; Perez, I.; Galindo, S.; Nieto-Miguel, T.; Lopez-Paniagua, M.; Fernandez, I.; Alberca, M.; Garcia-Sancho, J.; Sanchez, A.; Herreras, J.M. A proof-of-concept clinical trial using mesenchymal stem cells for the treatment of corneal epithelial stem cell deficiency. Transl. Res. 2019, 206, 18–40. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.Y.; Du, X.; Geng, S.X.; Peng, Y.W.; Wang, Z.; Lu, Z.S.; Wu, S.J.; Luo, C.W.; Guo, R.; Ling, W.; et al. Mesenchymal stem cell as salvage treatment for refractory chronic GVHD. Bone Marrow Transplant. 2010, 45, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.; He, C.; Lai, P.; Luo, C.; Guo, R.; Wu, S.; Geng, S.; Xiangpeng, A.; Liu, X.; Du, X. Mesenchymal stromal cells treatment attenuates dry eye in patients with chronic graft-versus-host disease. Mol. Ther. 2012, 20, 2347–2354. [Google Scholar] [CrossRef]
- Jeng, B.H.; Hamrah, P.; Kirshner, Z.Z.; Mendez, B.C.; Wessel, H.C.; Brown, L.R.; Steed, D.L. Exploratory phase II multicenter, open-label, clinical trial of ST266, a novel secretome for treatment of persistent corneal epithelial defects. Transl. Vis. Sci. Technol. 2022, 11, 8. [Google Scholar] [CrossRef]
- Moller-Hansen, M.; Larsen, A.C.; Toft, P.B.; Lynggaard, C.D.; Schwartz, C.; Bruunsgaard, H.; Haack-Sorensen, M.; Ekblond, A.; Kastrup, J.; Heegaard, S. Safety and feasibility of mesenchymal stem cell therapy in patients with aqueous deficient dry eye disease. Ocul. Surf. 2021, 19, 43–52. [Google Scholar] [CrossRef]
- Zhou, T.; He, C.; Lai, P.; Yang, Z.; Liu, Y.; Xu, H.; Lin, X.; Ni, B.; Ju, R.; Yi, W.; et al. miR-204-containing exosomes ameliorate GVHD-associated dry eye disease. Sci. Adv. 2022, 8, eabj9617. [Google Scholar] [CrossRef]
- El Zarif, M.; Alio Del Barrio, J.; Abdul Jawad, K.; Palazon-Bru, A.; Abdul Jawad, Z.; De Miguel, M.P.; Makdissy, N. Corneal stromal regeneration therapy for advanced keratoconus: Long-term outcomes at 3 years. Cornea 2021, 40, 741–754. [Google Scholar] [CrossRef]
- Chaurasia, S.S.; Perera, P.; Poh, R.; Wong, T.T.; Mehta, J.S. Hevin deficiency leads to aberrant extracellular matrix: A novel target for wound repair. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5301. [Google Scholar]
- Prelich, G. Gene overexpression: Uses, mechanisms, and interpretation. Genetics 2012, 190, 841–854. [Google Scholar] [CrossRef]
- Moriya, H. Quantitative nature of overexpression experiments. Mol. Biol. Cell 2015, 26, 3932–3939. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.W.; Lian, H.; Zhong, B.; Shu, H.B.; Li, S. Kruppel-like factor 4 negatively regulates cellular antiviral immune response. Cell. Mol. Immunol. 2016, 13, 65–72. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; He, J.; Xie, K. KLF4 transcription factor in tumorigenesis. Cell Death Discov. 2023, 9, 118. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.W.; Wu, J.M. Molecular mechanisms regulating expression and function of transcription regulator inhibitor of differentiation 3. Acta Pharmacol. Sin. 2005, 26, 1409–1420. [Google Scholar] [CrossRef] [PubMed]
- Mody, A.A.; Millar, J.C.; Clark, A.F. ID1 and ID3 are negative regulators of TGFβeta2-induced ocular hypertension and compromised aqueous humor outflow facility in mice. Investig. Ophthalmol. Vis. Sci. 2021, 62, 3. [Google Scholar] [CrossRef] [PubMed]
- Ha Thi, H.T.; Kim, H.Y.; Lee, Y.J.; Kim, S.J.; Hong, S. SMAD7 in keratinocytes promotes skin carcinogenesis by activating ATM-dependent DNA repair and an EGFR-mediated cell proliferation pathway. Carcinogenesis 2019, 40, 112–120. [Google Scholar] [CrossRef] [PubMed]
- de Ceuninck van Capelle, C.; Spit, M.; Ten Dijke, P. Current perspectives on inhibitory SMAD7 in health and disease. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 691–715. [Google Scholar] [CrossRef]
- Alarmo, E.L.; Korhonen, T.; Kuukasjarvi, T.; Huhtala, H.; Holli, K.; Kallioniemi, A. Bone morphogenetic protein 7 expression associates with bone metastasis in breast carcinomas. Ann. Oncol. 2008, 19, 308–314. [Google Scholar] [CrossRef]
- Guo, J.; Lin, Q.; Shao, Y.; Rong, L.; Zhang, D. BMP-7 suppresses excessive scar formation by activating the BMP-7/Smad1/5/8 signaling pathway. Mol. Med. Rep. 2017, 16, 1957–1963. [Google Scholar] [CrossRef]
- Jarvinen, T.A.; Prince, S. Decorin: A growth factor antagonist for tumor growth inhibition. BioMed Res. Int. 2015, 2015, 654765. [Google Scholar] [CrossRef]
- Yoon, S.; Kang, G.; Eom, G.H. HDAC Inhibitors: Therapeutic Potential in Fibrosis-Associated Human Diseases. Int. J. Mol. Sci. 2019, 20, 1329. [Google Scholar] [CrossRef]
- Fu, Y.; Saxu, R.; Ridwan, K.A.; Yao, J.; Chen, X.; Xu, X.; Zheng, W.; Yu, P.; Teng, Y. Losartan Alleviates the Side Effects and Maintains the Anticancer Activity of Axitinib. Molecules 2022, 27, 2764. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Mizuno, S. The discovery of hepatocyte growth factor (HGF) and its significance for cell biology, life sciences and clinical medicine. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 588–610. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, L.M.; Barillas, S.; Weis, S.M.; Gothert, J.R.; Cheresh, D.A. Semaphorin 3A suppresses VEGF-mediated angiogenesis yet acts as a vascular permeability factor. Blood 2008, 111, 2674–2680. [Google Scholar] [CrossRef] [PubMed]
- Shelly, M.; Cancedda, L.; Lim, B.K.; Popescu, A.T.; Cheng, P.L.; Gao, H.; Poo, M.M. Semaphorin3A regulates neuronal polarization by suppressing axon formation and promoting dendrite growth. Neuron 2011, 71, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Han, H. RNA Interference to Knock Down Gene Expression. Methods Mol. Biol. 2018, 1706, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, L.; Ma, W.; Cao, X.; Chen, H.; Feng, D.; Liang, J.; Yin, K.; Jiang, X. The blockage of KCa3.1 channel inhibited proliferation, migration and promoted apoptosis of human hepatocellular carcinoma cells. J. Cancer 2015, 6, 643–651. [Google Scholar] [CrossRef]
- Bachmann, M.; Rossa, A.; Varanita, T.; Fioretti, B.; Biasutto, L.; Milenkovic, S.; Checchetto, V.; Peruzzo, R.; Ahmad, S.A.; Patel, S.H.; et al. Pharmacological targeting of the mitochondrial calcium-dependent potassium channel KCa3.1 triggers cell death and reduces tumor growth and metastasis in vivo. Cell Death Dis. 2022, 13, 1055. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chandran, C.; Santra, M.; Rubin, E.; Geary, M.L.; Yam, G.H.-F. Regenerative Therapy for Corneal Scarring Disorders. Biomedicines 2024, 12, 649. https://doi.org/10.3390/biomedicines12030649
Chandran C, Santra M, Rubin E, Geary ML, Yam GH-F. Regenerative Therapy for Corneal Scarring Disorders. Biomedicines. 2024; 12(3):649. https://doi.org/10.3390/biomedicines12030649
Chicago/Turabian StyleChandran, Christine, Mithun Santra, Elizabeth Rubin, Moira L. Geary, and Gary Hin-Fai Yam. 2024. "Regenerative Therapy for Corneal Scarring Disorders" Biomedicines 12, no. 3: 649. https://doi.org/10.3390/biomedicines12030649