
Impaired Response to Mismatch Novelty in the Li2+-Pilocarpine Rat Model of TLE: Correlation with Hippocampal Monoaminergic Inputs



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
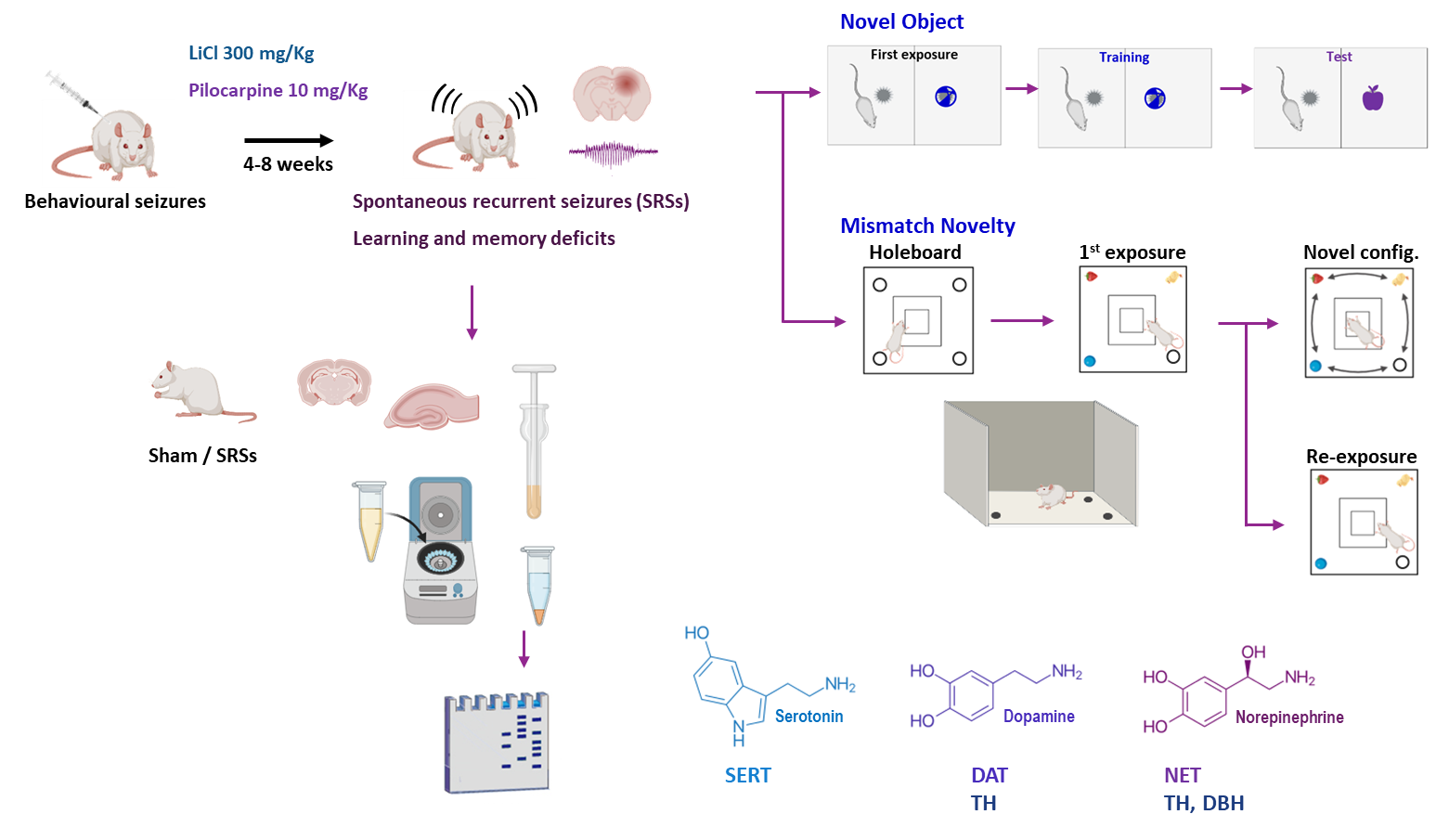
2.1. Animals and Induction of SRSs
2.2. Evaluation of Anxiety and Locomotion Using the EPM and OF
2.3. Evaluation of Cognitive Performance in the RAM
2.4. Novel Object Recognition (NOR)
2.5. Mismatch Novelty Test
2.6. Western Blot Analysis of Monoaminergic Markers and Synaptic Proteins
2.7. Statistics
3. Results
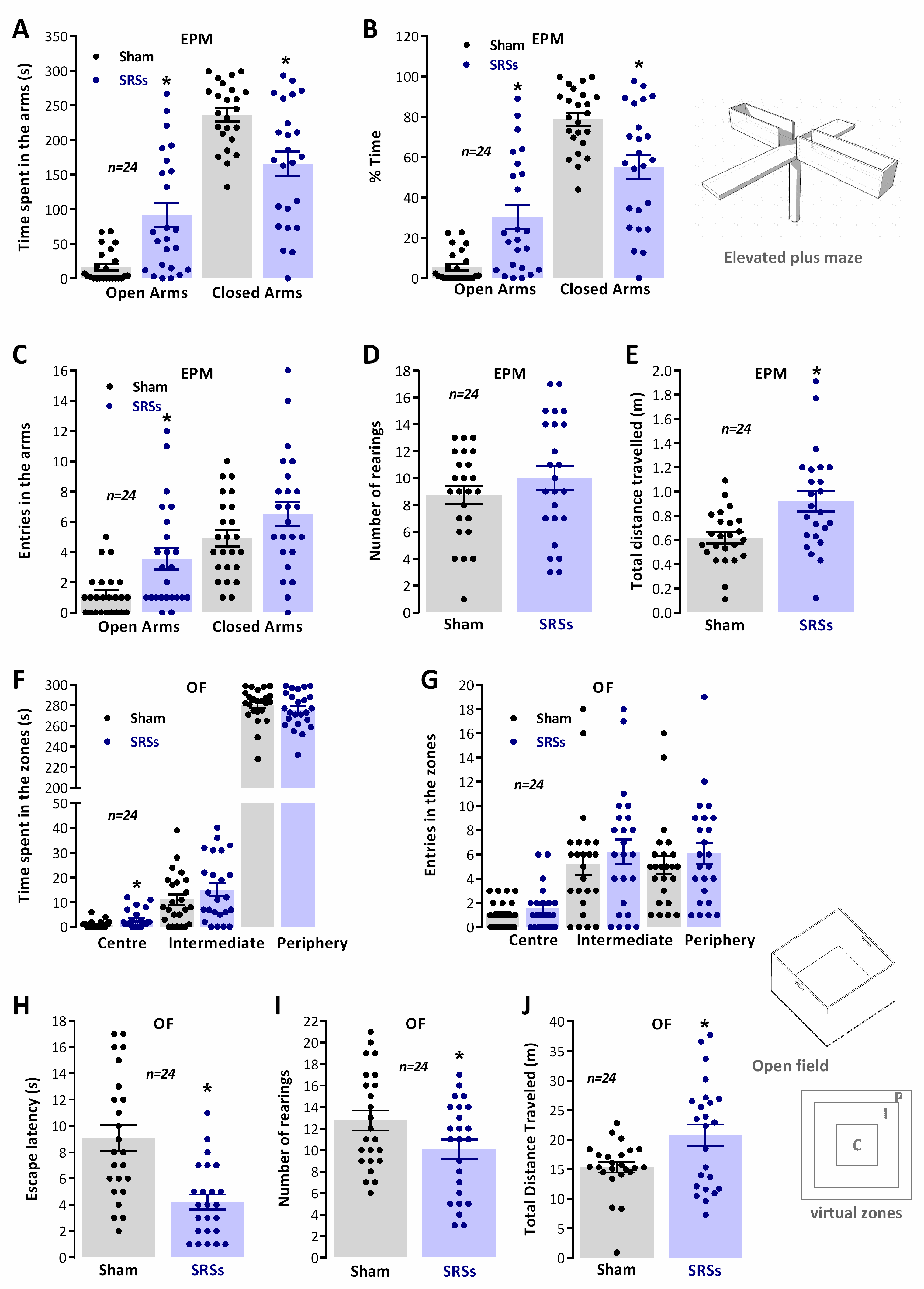
3.1. Elevated Plus Maze (EPM) and Open-Field (OF) Tests
3.2. Spatial Learning in the RAM
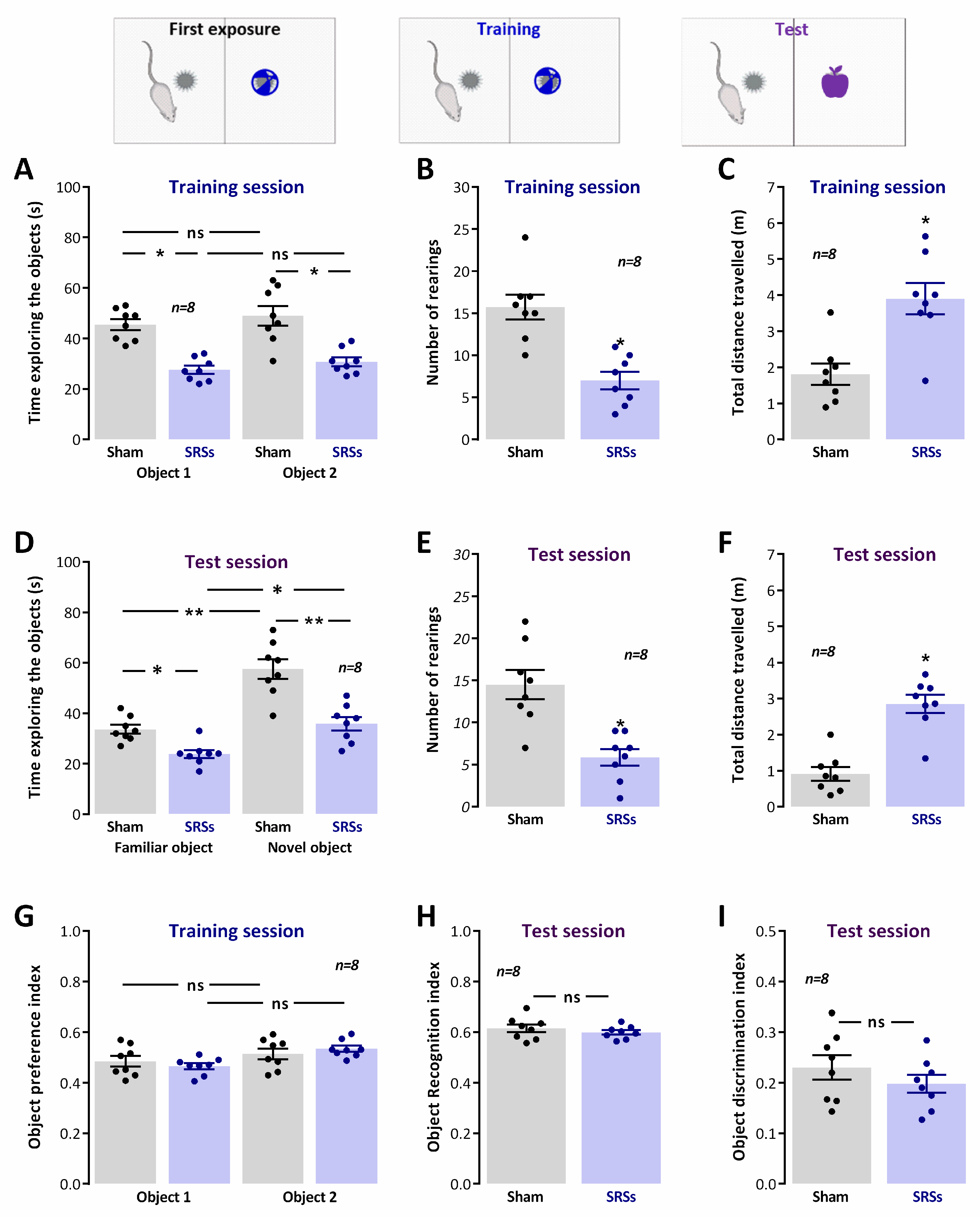
3.3. Novel Object Recognition
3.4. Mismatch Novelty Detection
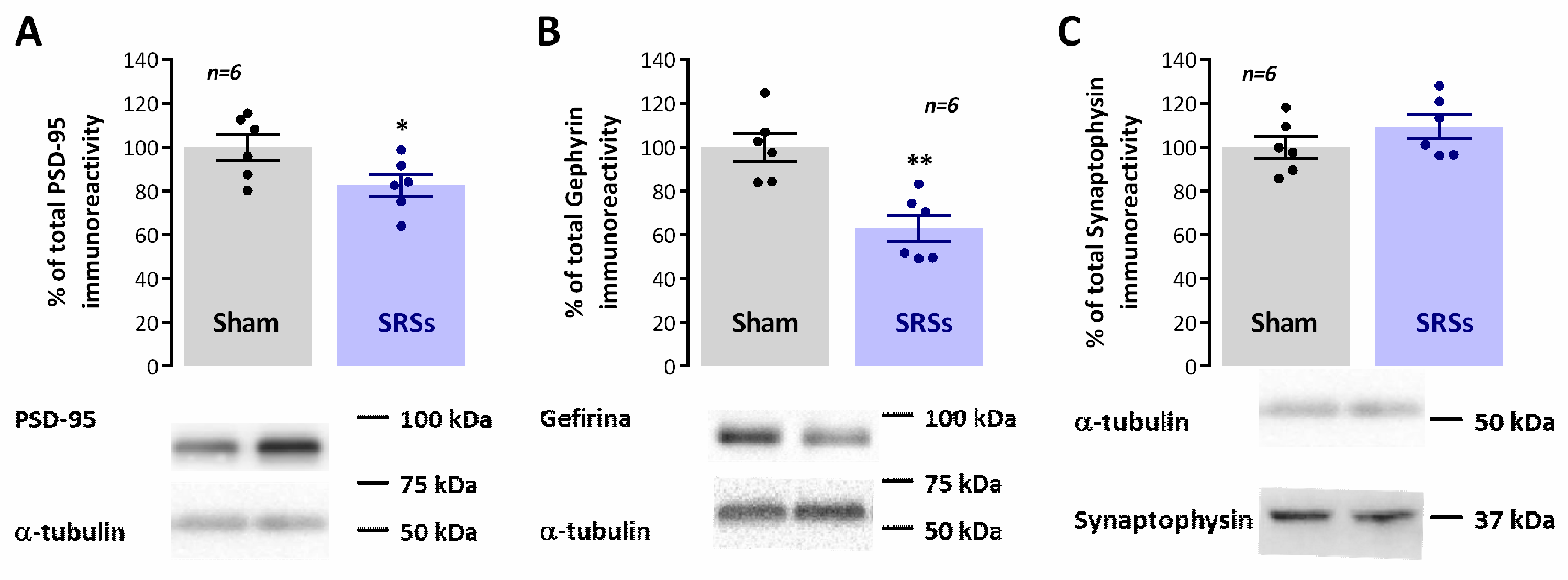
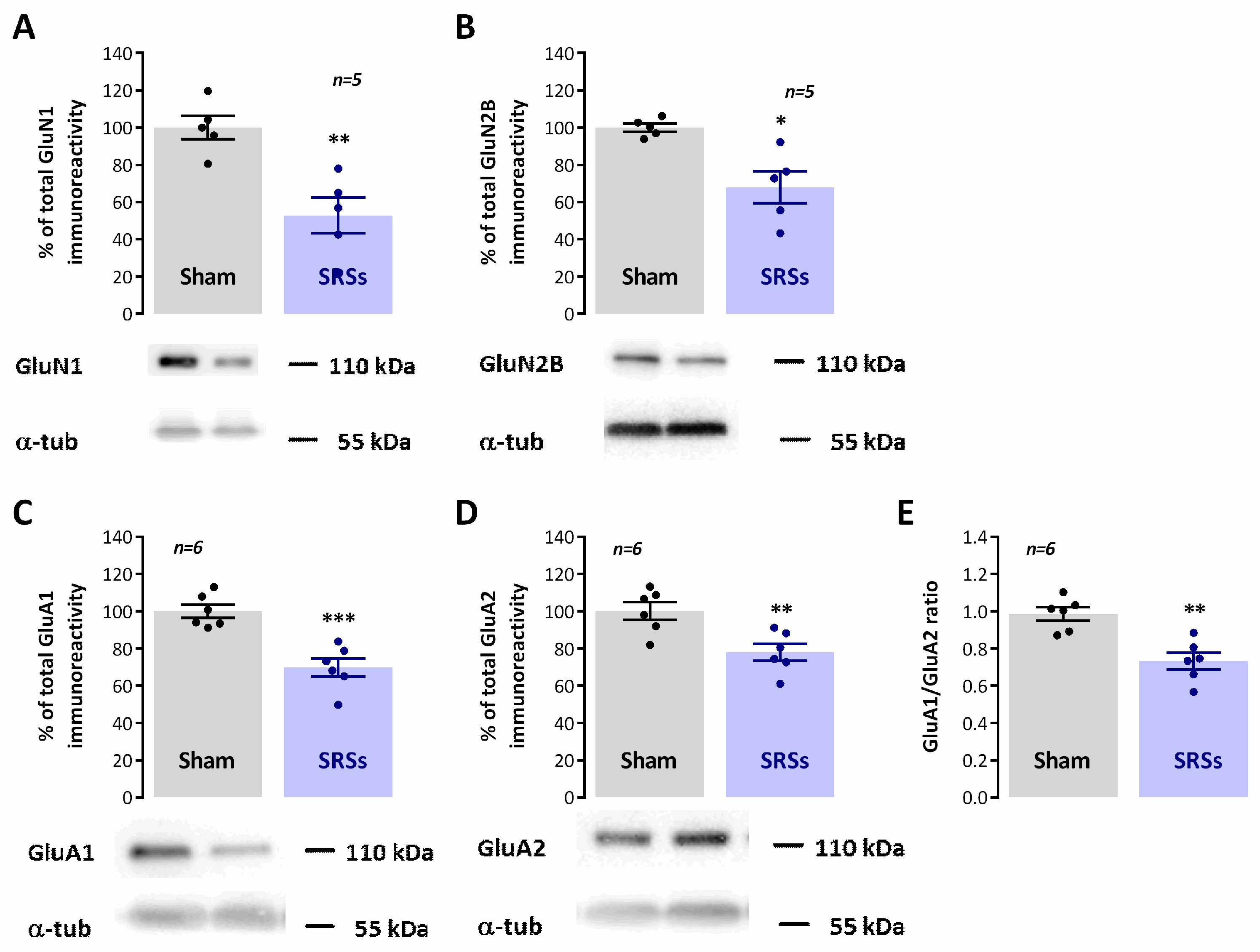
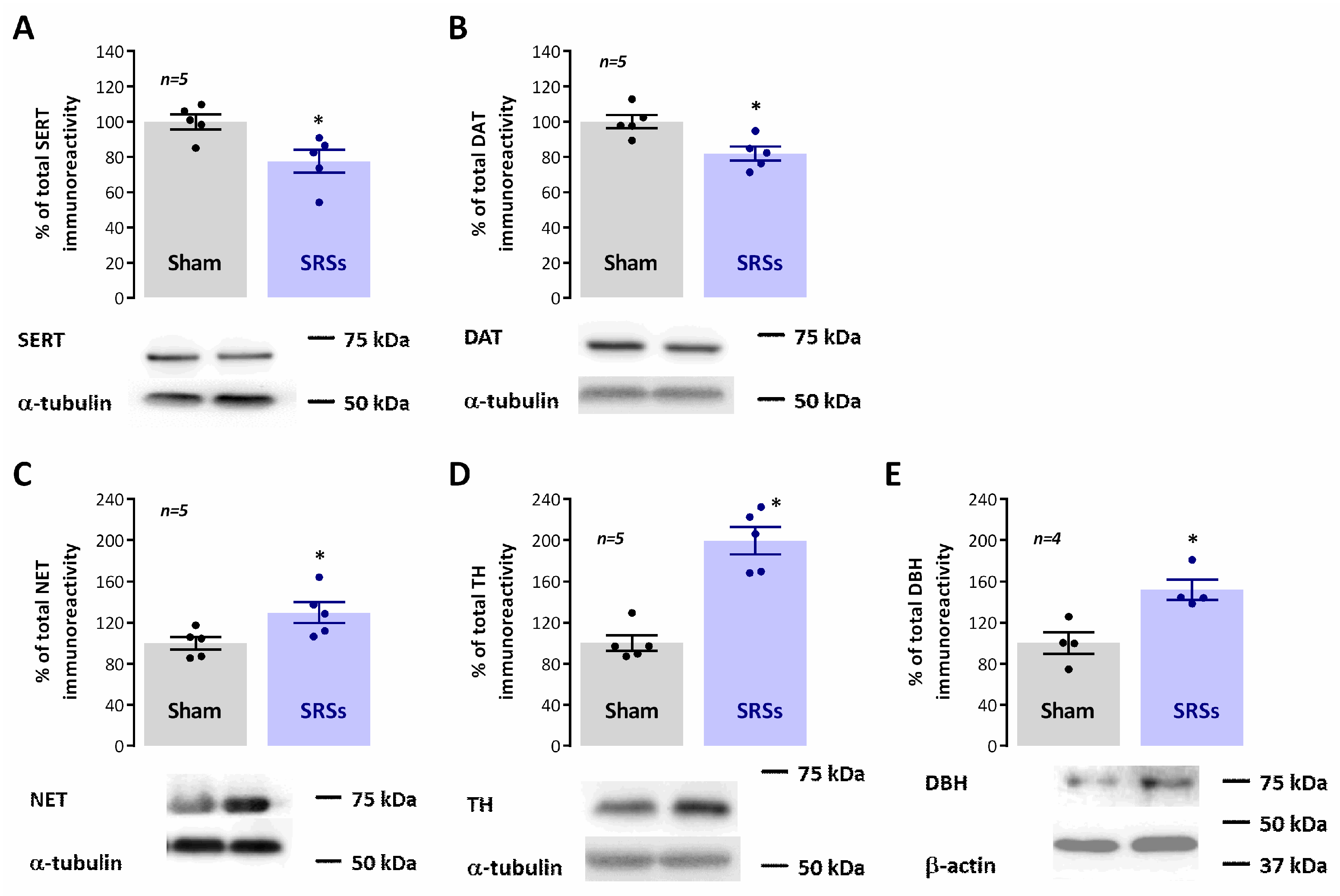
3.5. Evaluation of Hippocampal Levels of Synaptic and Monoaminergic Markers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devinsky, O.; Vezzani, A.; O’Brien, T.J.; Jette, N.; Scheffer, I.E.; De Curtis, M.; Perucca, P. Epilepsy. Nat. Rev. Dis. Prim. 2018, 4, 18024. [Google Scholar] [CrossRef]
- Moshé, S.L.; Perucca, E.; Ryvlin, P.; Tomson, T. Epilepsy: New Advances. Lancet 2015, 385, 884–898. [Google Scholar] [CrossRef]
- Vezzani, A.; Balosso, S.; Ravizza, T. Neuroinflammatory Pathways as Treatment Targets and Biomarkers in Epilepsy. Nat. Rev. Neurol. 2019, 15, 459–472. [Google Scholar] [CrossRef]
- Cunha-Reis, D.; Caulino-Rocha, A.; Correia-de-Sá, P. VIPergic Neuroprotection in Epileptogenesis: Challenges and Opportunities. Pharmacol. Res. 2021, 164, 105356. [Google Scholar] [CrossRef] [PubMed]
- Thom, M. Review: Hippocampal Sclerosis in Epilepsy: A Neuropathology Review. Neuropathol. Appl. Neurobiol. 2014, 40, 520–543. [Google Scholar] [CrossRef] [PubMed]
- Helmstaedter, C.; Kockelmann, E. Cognitive Outcomes in Patients with Chronic Temporal Lobe Epilepsy. Epilepsia 2006, 47 (Suppl. S2), 96–98. [Google Scholar] [CrossRef] [PubMed]
- Schomaker, J.; Grouls, M.M.E.; van der Linden, C.G.M.; Rau, E.M.; Hendriks, M.; Colon, A.; Meeter, M. Novelty Processing Depends on Medial Temporal Lobe Structures. Neurobiol. Learn. Mem. 2021, 183, 107464. [Google Scholar] [CrossRef] [PubMed]
- Celiker Uslu, S.; Yuksel, B.; Tekin, B.; Sariahmetoglu, H.; Atakli, D. Cognitive Impairment and Drug Responsiveness in Mesial Temporal Lobe Epilepsy. Epilepsy Behav. 2019, 90, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Poprelka, K.; Patrikelis, P.; Takousi, M.; Messinis, L.; Fasilis, T.; Margariti, S.; Ntinopoulou, E.; Verentzioti, A.; Stefanatou, M.; Alexoudi, A.; et al. Arousal Deregulation in the Co-Shaping of Neuropsychological Dysfunction in Frontal and Mesial Temporal Lobe Epilepsy. Epilepsy Res. 2023, 194, 107189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lu, G.; Zhong, Y.; Tan, Q.; Yang, Z.; Liao, W.; Chen, Z.; Shi, J.; Liu, Y. Impaired Attention Network in Temporal Lobe Epilepsy: A Resting FMRI Study. Neurosci. Lett. 2009, 458, 97–101. [Google Scholar] [CrossRef]
- Kemp, A.; Manahan-Vaughan, D. Hippocampal Long-Term Depression and Long-Term Potentiation Encode Different Aspects of Novelty Acquisition. Proc. Natl. Acad. Sci. USA 2004, 101, 8192–8197. [Google Scholar] [CrossRef] [PubMed]
- Kemp, A.; Manahan-Vaughan, D. Hippocampal Long-Term Depression: Master or Minion in Declarative Memory Processes? Trends Neurosci. 2007, 30, 111–118. [Google Scholar] [CrossRef]
- Dong, Z.; Gong, B.; Li, H.; Bai, Y.; Wu, X.; Huang, Y.; He, W.; Li, T.; Wang, Y.T. Mechanisms of Hippocampal Long-Term Depression Are Required for Memory Enhancement by Novelty Exploration. J. Neurosci. 2012, 32, 11980–11990. [Google Scholar] [CrossRef]
- Ge, Y.; Dong, Z.; Bagot, R.C.; Howland, J.G.; Phillips, A.G.; Wong, T.P.; Wang, Y.T. Hippocampal Long-Term Depression Is Required for the Consolidation of Spatial Memory. Proc. Natl. Acad. Sci. USA 2010, 107, 16697–16702. [Google Scholar] [CrossRef] [PubMed]
- Abraham, W.C.; Bear, M.F. Metaplasticity: The Plasticity of Synaptic Plasticity. Trends Neurosci. 1996, 19, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, L.A.; Viola, H.; Barros, D.M.; Alonso, M.; Vianna, M.R.M.; Furman, M.; Levi De Stein, M.; Szapiro, G.; Rodrigues, C.; Choi, H.; et al. Novelty Enhances Retrieval: Molecular Mechanisms Involved in Rat Hippocampus. Eur. J. Neurosci. 2001, 13, 1464–1467. [Google Scholar] [CrossRef]
- Stepankova, K.; Fenton, A.A.; Pastalkova, E.; Kalina, M.; Bohbot, V.É.D. Object-Location Memory Impairment in Patients with Thermal Lesions to the Right or Left Hippocampus. Neuropsychologia 2004, 42, 1017–1028. [Google Scholar] [CrossRef]
- Bohbot, V.D.; Jech, R.; Růžička, E.; Nadel, L.; Kalina, M.; Stěpánková, K.; Bureš, J. Rat Spatial Memory Tasks Adapted for Humans: Characterization in Subjects with Intact Brain and Subjects with Medial Temporal Lobe Lesions. Physiol. Res. 2002, 51, 49–64. [Google Scholar] [CrossRef]
- Aidil-Carvalho, M.F.; Carmo, A.J.S.; Ribeiro, J.A.; Cunha-Reis, D. Mismatch Novelty Exploration Training Enhances Hippocampal Synaptic Plasticity: A Tool for Cognitive Stimulation? Neurobiol. Learn. Mem. 2017, 145, 240–250. [Google Scholar] [CrossRef]
- Kempadoo, K.A.; Mosharov, E.V.; Choi, S.J.; Sulzer, D.; Kandel, E.R. Dopamine Release from the Locus Coeruleus to the Dorsal Hippocampus Promotes Spatial Learning and Memory. Proc. Natl. Acad. Sci. USA 2016, 113, 14835–14840. [Google Scholar] [CrossRef]
- Takeuchi, T.; Duszkiewicz, A.J.; Sonneborn, A.; Spooner, P.A.; Yamasaki, M.; Watanabe, M.; Smith, C.C.; Fernández, G.; Deisseroth, K.; Greene, R.W.; et al. Locus Coeruleus and Dopaminergic Consolidation of Everyday Memory. Nature 2016, 537, 357–362. [Google Scholar] [CrossRef]
- Twarkowski, H.; Manahan-Vaughan, D. Loss of Catecholaminergic Neuromodulation of Persistent Forms of Hippocampal Synaptic Plasticity with Increasing Age. Front. Synaptic Neurosci. 2016, 8, 30. [Google Scholar] [CrossRef]
- Titulaer, J.; Björkholm, C.; Feltmann, K.; Malmlöf, T.; Mishra, D.; Bengtsson Gonzales, C.; Schilström, B.; Konradsson-Geuken, Å. The Importance of Ventral Hippocampal Dopamine and Norepinephrine in Recognition Memory. Front. Behav. Neurosci. 2021, 73, 667244. [Google Scholar] [CrossRef]
- Kurian, M.A.; Gissen, P.; Smith, M.; Heales, S.J.R.; Clayton, P.T. The Monoamine Neurotransmitter Disorders: An Expanding Range of Neurological Syndromes. Lancet Neurol. 2011, 10, 721–733. [Google Scholar] [CrossRef]
- Marinho, A.M.d.N.; Lobão-Soares, B.; Targino, H.A.S.; Vasconcellos, Y.J.F.; Guarnieri, R.; Carlotti, C.G. Decreased Hippocampal Serotonin 5HT1A Expression in Mesial Temporal Lobe of Epilepsy Patients. Epilepsy Behav. 2022, 129, 108574. [Google Scholar] [CrossRef] [PubMed]
- Maia, G.H.; Soares, J.I.; Andrade, P.A.; Leite, J.F.; Luz, L.L.; Andrade, J.P.; Lukoyanov, N.V. Altered Taste Preference and Loss of Limbic-Projecting Serotonergic Neurons in the Dorsal Raphe Nucleus of Chronically Epileptic Rats. Behav. Brain Res. 2016, 297, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Medel-Matus, J.S.; Shin, D.; Sankar, R.; Mazarati, A. Inherent Vulnerabilities in Monoaminergic Pathways Predict the Emergence of Depressive Impairments in an Animal Model of Chronic Epilepsy. Epilepsia 2017, 58, e116–e121. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Cao, Q.; Li, J.; Zhang, Y.; Liu, X.; Wang, Z.; Guo, F.; Chen, Y.; Chen, Y.; Chen, G.; et al. Altered Expression of Vesicular Monoamine Transporter 2 in Epileptic Patients and Experimental Rats. Synapse 2013, 67, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, C.; Guerreiro-Pinto, V.; Pawlak, S.; Caulino-Rocha, A.; Amat-Garcia, L.; Cunha-Reis, D. Impaired Response to Mismatch Novelty in the Li2+-Pilocarpine Rat Model of TLE: Correlation with Hippocampal Monoaminergic Inputs. bioRxiv 2024, 2024.02.07.579299. [Google Scholar] [CrossRef]
- Caulino-Rocha, A.; Nascimento, C.; Amaro-Leal, A.; Serpa, A.; Cunha-Reis, D. Impaired Response to Mismatch Novelty in the Li2+-Pilocarpine Rat Model of TLE: Correlation with Synaptic Remodeling and Hippocampal Monoamines. Epilepsia 2023, 64, 227–228. [Google Scholar] [CrossRef]
- Glien, M.; Brandt, C.; Potschka, H.; Voigt, H.; Ebert, U.; Löscher, W. Repeated Low-Dose Treatment of Rats with Pilocarpine: Low Mortality but High Proportion of Rats Developing Epilepsy. Epilepsy Res. 2001, 46, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Lüttjohann, A.; Fabene, P.F.; van Luijtelaar, G. A Revised Racine’s Scale for PTZ-Induced Seizures in Rats. Physiol. Behav. 2009, 98, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Crusio, W.E.; Schwegler, H.; van Abeelen, J.H.F. Behavioral Responses to Novelty and Structural Variation of the Hippocampus in Mice. II. Multivariate Genetic Analysis. Behav. Brain Res. 1989, 32, 81–88. [Google Scholar] [CrossRef]
- Schneider, P.; Ho, Y.-J.; Spanagel, R.; Pawlak, C.R. A Novel Elevated Plus-Maze Procedure to Avoid the One-Trial Tolerance Problem. Front. Behav. Neurosci. 2011, 5, 43. [Google Scholar] [CrossRef]
- Goh, J.J.; Manahan-Vaughan, D. Spatial Object Recognition Enables Endogenous LTD That Curtails LTP in the Mouse Hippocampus. Cereb. Cortex 2013, 23, 1118–1125. [Google Scholar] [CrossRef]
- Rodrigues, N.C.; Silva-Cruz, A.; Caulino-Rocha, A.; Bento-Oliveira, A.; Alexandre Ribeiro, J.; Cunha-Reis, D. Hippocampal CA1 Theta Burst-Induced LTP from Weaning to Adulthood: Cellular and Molecular Mechanisms in Young Male Rats Revisited. Eur. J. Neurosci. 2021, 54, 5272–5292. [Google Scholar] [CrossRef] [PubMed]
- Ramos, F.O.; Carreiro, L.R.R.; Scorza, F.A.; Cysneiros, R.M. Impaired Executive Functions in Experimental Model of Temporal Lobe Epilepsy. Arq. Neuropsiquiatr. 2016, 74, 470–477. [Google Scholar] [CrossRef]
- Pineda, E.; Jentsch, J.D.; Shin, D.; Griesbach, G.; Sankar, R.; Mazarati, A. Behavioral Impairments in Rats with Chronic Epilepsy Suggest Comorbidity between Epilepsy and Attention Deficit/Hyperactivity Disorder. Epilepsy Behav. 2014, 31, 267–275. [Google Scholar] [CrossRef]
- Dong, Z.; Bai, Y.; Wu, X.; Li, H.; Gong, B.; Howland, J.G.; Huang, Y.; He, W.; Li, T.; Wang, Y.T. Hippocampal Long-Term Depression Mediates Spatial Reversal Learning in the Morris Water Maze. Neuropharmacology 2013, 64, 65–73. [Google Scholar] [CrossRef]
- Valenti, O.; Mikus, N.; Klausberger, T. The Cognitive Nuances of Surprising Events: Exposure to Unexpected Stimuli Elicits Firing Variations in Neurons of the Dorsal CA1 Hippocampus. Brain Struct. Funct. 2018, 223, 3183–3211. [Google Scholar] [CrossRef]
- Curia, G.; Longo, D.; Biagini, G.; Jones, R.S.G.; Avoli, M. The Pilocarpine Model of Temporal Lobe Epilepsy. J. Neurosci. Methods 2008, 172, 143–157. [Google Scholar] [CrossRef]
- Serpa, A.; Bento, M.; Caulino-Rocha, A.; Pawlak, S.; Cunha-Reis, D. Opposing Reduced VPAC1 and Enhanced VPAC2 VIP Receptors in the Hippocampus of the Li2+-Pilocarpine Rat Model of Temporal Lobe Epilepsy. Neurochem. Int. 2022, 158, 105383. [Google Scholar] [CrossRef]
- Vinogradova, O.S.; Kitchigina, V.F.; Kudina, T.A.; Zenchenko, K.I. Spontaneous Activity and Sensory Responses of Hippocampal Neurons during Persistent Theta-Rhythm Evoked by Median Raphe Nucleus Blockade in Rabbit. Neuroscience 1999, 94, 745–753. [Google Scholar] [CrossRef]
- Borhegyi, Z.; Varga, V.; Szilágyi, N.; Fabo, D.; Freund, T.F. Phase Segregation of Medial Septal GABAergic Neurons during Hippocampal Theta Activity. J. Neurosci. 2004, 24, 8470–8479. [Google Scholar] [CrossRef]
- Vandecasteele, M.; Varga, V.; Berényi, A.; Papp, E.; Barthó, P.; Venance, L.; Freund, T.F.; Buzsáki, G. Optogenetic Activation of Septal Cholinergic Neurons Suppresses Sharp Wave Ripples and Enhances Theta Oscillations in the Hippocampus. Proc. Natl. Acad. Sci. USA 2014, 111, 13535–13540. [Google Scholar] [CrossRef]
- Teixeira, C.M.; Rosen, Z.B.; Suri, D.; Sun, Q.; Hersh, M.; Sargin, D.; Dincheva, I.; Morgan, A.A.; Spivack, S.; Krok, A.C.; et al. Hippocampal 5-HT Input Regulates Memory Formation and Schaffer Collateral Excitation. Neuron 2018, 98, 992–1004.e4. [Google Scholar] [CrossRef] [PubMed]
- De Leo, G.; Gulino, R.; Coradazzi, M.; Leanza, G. Acetylcholine and Noradrenaline Differentially Regulate Hippocampus-Dependent Spatial Learning and Memory. Brain Commun. 2022, 5, fcac338. [Google Scholar] [CrossRef] [PubMed]
- Twarkowski, H.; Hagena, H.; Manahan-Vaughan, D. The 5-Hydroxytryptamine 4 Receptor Enables Differentiation of Informational Content and Encoding in the Hippocampus. Hippocampus 2016, 26, 875–891. [Google Scholar] [CrossRef] [PubMed]
- Nachtigall, E.G.; Furini, C.R.G.; Behling, J.A.K.; Farias, C.P.; Izquierdo, I.; Myskiw, J.d.C. Facilitation of Fear Extinction by Novelty Is Modulated by β-Adrenergic and 5-HT1A Serotoninergic Receptors in Hippocampus. Neurobiol. Learn. Mem. 2019, 166, 107101. [Google Scholar] [CrossRef] [PubMed]
- Pimpinella, D.; Mastrorilli, V.; Giorgi, C.; Coemans, S.; Lecca, S.; Lalive, A.L.; Ostermann, H.; Fuchs, E.C.; Monyer, H.; Mele, A.; et al. Septal Cholinergic Input to Ca2 Hippocampal Region Controls Social Novelty Discrimination via Nicotinic Receptor-Mediated Disinhibition. Elife 2021, 10, 65580. [Google Scholar] [CrossRef] [PubMed]
- Garrido Sanabria, E.R.; Castañeda, M.T.; Banuelos, C.; Perez-Cordova, M.G.; Hernandez, S.; Colom, L.V. Septal GABAergic Neurons Are Selectively Vulnerable to Pilocarpine-Induced Status Epilepticus and Chronic Spontaneous Seizures. Neuroscience 2006, 142, 871–883. [Google Scholar] [CrossRef]
- Soares, J.I.; Valente, M.C.; Andrade, P.A.; Maia, G.H.; Lukoyanov, N.V. Reorganization of the Septohippocampal Cholinergic Fiber System in Experimental Epilepsy. J. Comp. Neurol. 2017, 525, 2690–2705. [Google Scholar] [CrossRef]
- Patodia, S.; Somani, A.; Liu, J.; Cattaneo, A.; Paradiso, B.; Garcia, M.; Othman, M.; Diehl, B.; Devinsky, O.; Mills, J.D.; et al. Serotonin Transporter in the Temporal Lobe, Hippocampus and Amygdala in SUDEP. Brain Pathol. 2022, 32, e13074. [Google Scholar] [CrossRef]
- Kamiński, J.; Mamelak, A.N.; Birch, K.; Mosher, C.P.; Tagliati, M.; Rutishauser, U. Novelty-Sensitive Dopaminergic Neurons in the Human Substantia Nigra Predict Success of Declarative Memory Formation. Curr. Biol. 2018, 28, 1333–1343.e4. [Google Scholar] [CrossRef] [PubMed]
- Wideman, C.E.; Jardine, K.H.; Winters, B.D. Involvement of Classical Neurotransmitter Systems in Memory Reconsolidation: Focus on Destabilization. Neurobiol. Learn. Mem. 2018, 156, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Ahern, T.H.; Javors, M.A.; Eagles, D.A.; Martillotti, J.; Mitchell, H.A.; Liles, L.C.; Weinshenker, D. The Effects of Chronic Norepinephrine Transporter Inactivation on Seizure Susceptibility in Mice. Neuropsychopharmacology 2005, 31, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Bengzon, J.; Hansson, S.R.; Hoffman, B.J.; Lindvall, O. Regulation of Norepinephrine Transporter and Tyrosine Hydroxylase MRNAs after Kainic Acid-Induced Seizures. Brain Res. 1999, 842, 239–242. [Google Scholar] [CrossRef]
- Akyuz, E.; Doganyigit, Z.; Paudel, Y.N.; Koklu, B.; Kaymak, E.; Villa, C.; Arulsamy, A.; Shaikh, M.F.; Devinsky, O. Immunoreactivity of Muscarinic Acetylcholine M2 and Serotonin 5-HT2B Receptors, Norepinephrine Transporter and Kir Channels in a Model of Epilepsy. Life 2021, 11, 276. [Google Scholar] [CrossRef] [PubMed]
- Cossart, R.; Dinocourt, C.; Hirsch, J.C.; Merchan-Perez, A.; De Felipe, J.; Ben-Ari, Y.; Esclapez, M.; Bernard, C. Dendritic but Not Somatic GABAergic Inhibition Is Decreased in Experimental Epilepsy. Nat. Neurosci. 2001, 4, 52–62. [Google Scholar] [CrossRef]
- Wyeth, M.; Buckmaster, P.S. Lack of Hyperinhibition of Oriens Lacunosum-Moleculare Cells by Vasoactive Intestinal Peptide-Expressing Cells in a Model of Temporal Lobe Epilepsy. eNeuro 2021, 8, 0299-21.2021. [Google Scholar] [CrossRef]
- David, L.S.; Topolnik, L. Target-Specific Alterations in the VIP Inhibitory Drive to Hippocampal GABAergic Cells after Status Epilepticus. Exp. Neurol. 2017, 292, 102–112. [Google Scholar] [CrossRef]
- Rahimi, S.; Salami, P.; Matulewicz, P.; Schmuck, A.; Bukovac, A.; Ramos-Prats, A.; Tasan, R.O.; Drexel, M. The Role of Subicular VIP-Expressing Interneurons on Seizure Dynamics in the Intrahippocampal Kainic Acid Model of Temporal Lobe Epilepsy. Exp. Neurol. 2023, 370, 114580. [Google Scholar] [CrossRef]
- Sun, Q.-J.J.; Duan, R.-S.S.; Wang, A.-H.H.; Shang, W.; Zhang, T.; Zhang, X.-Q.Q.; Chi, Z.-F.F. Alterations of NR2B and PSD-95 Expression in Hippocampus of Kainic Acid-Exposed Rats with Behavioural Deficits. Behav. Brain Res. 2009, 201, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Ren, E.; Curia, G. Synaptic Reshaping and Neuronal Outcomes in the Temporal Lobe Epilepsy. Int. J. Mol. Sci. 2021, 22, 3860. [Google Scholar] [CrossRef]
- Keith, D.; El-Husseini, A. Excitation Control: Balancing PSD-95 Function at the Synapse. Front. Mol. Neurosci. 2008, 1, 4. [Google Scholar] [CrossRef]
- Hamamoto, O.; Tirapelli, D.P.d.C.; Lizarte Neto, F.S.; Freitas-Lima, P.; Saggioro, F.P.; Cirino, M.L.d.A.; Assirati, J.A.; Serafini, L.N.; Velasco, T.R.; Sakamoto, A.C.; et al. Modulation of NMDA Receptor by MiR-219 in the Amygdala and Hippocampus of Patients with Mesial Temporal Lobe Epilepsy. J. Clin. Neurosci. 2020, 74, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Hanada, T. Ionotropic Glutamate Receptors in Epilepsy: A Review Focusing on AMPA and NMDA Receptors. Biomolecules 2020, 10, 464. [Google Scholar] [CrossRef] [PubMed]
- Curia, G. Hebbian and Homeostatic Synaptic Plasticity of AMPA Receptors in Epileptogenesis. Cell Rep. Med. 2023, 4, 101047. [Google Scholar] [CrossRef]
- Issa, N.P.; Nunn, K.C.; Wu, S.; Haider, H.A.; Tao, J.X. Putative Roles for Homeostatic Plasticity in Epileptogenesis. Epilepsia 2023, 64, 539–552. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, P.; Yan, F.; Luo, Y.; Zhao, G. Animal Models of Epilepsy: A Phenotype-Oriented Review. Aging Dis. 2022, 13, 215–231. [Google Scholar] [CrossRef]
- Kandratavicius, L.; Balista, P.A.; Lopes-Aguiar, C.; Ruggiero, R.N.; Umeoka, E.H.; Garcia-Cairasco, N.; Bueno-Junior, L.S.; Leite, J.P. Animal Models of Epilepsy: Use and Limitations. Neuropsychiatr. Dis. Treat. 2014, 10, 1693–1705. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nascimento, C.; Guerreiro-Pinto, V.; Pawlak, S.; Caulino-Rocha, A.; Amat-Garcia, L.; Cunha-Reis, D. Impaired Response to Mismatch Novelty in the Li2+-Pilocarpine Rat Model of TLE: Correlation with Hippocampal Monoaminergic Inputs. Biomedicines 2024, 12, 631. https://doi.org/10.3390/biomedicines12030631
Nascimento C, Guerreiro-Pinto V, Pawlak S, Caulino-Rocha A, Amat-Garcia L, Cunha-Reis D. Impaired Response to Mismatch Novelty in the Li2+-Pilocarpine Rat Model of TLE: Correlation with Hippocampal Monoaminergic Inputs. Biomedicines. 2024; 12(3):631. https://doi.org/10.3390/biomedicines12030631
Chicago/Turabian StyleNascimento, Carlos, Vasco Guerreiro-Pinto, Seweryn Pawlak, Ana Caulino-Rocha, Laia Amat-Garcia, and Diana Cunha-Reis. 2024. "Impaired Response to Mismatch Novelty in the Li2+-Pilocarpine Rat Model of TLE: Correlation with Hippocampal Monoaminergic Inputs" Biomedicines 12, no. 3: 631. https://doi.org/10.3390/biomedicines12030631