
Detailed Mechanisms Underlying Neutrophil Bactericidal Activity against Streptococcus pneumoniae
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Streptococcus Pneumoniae Culture
2.2. Bacterial Opsonization
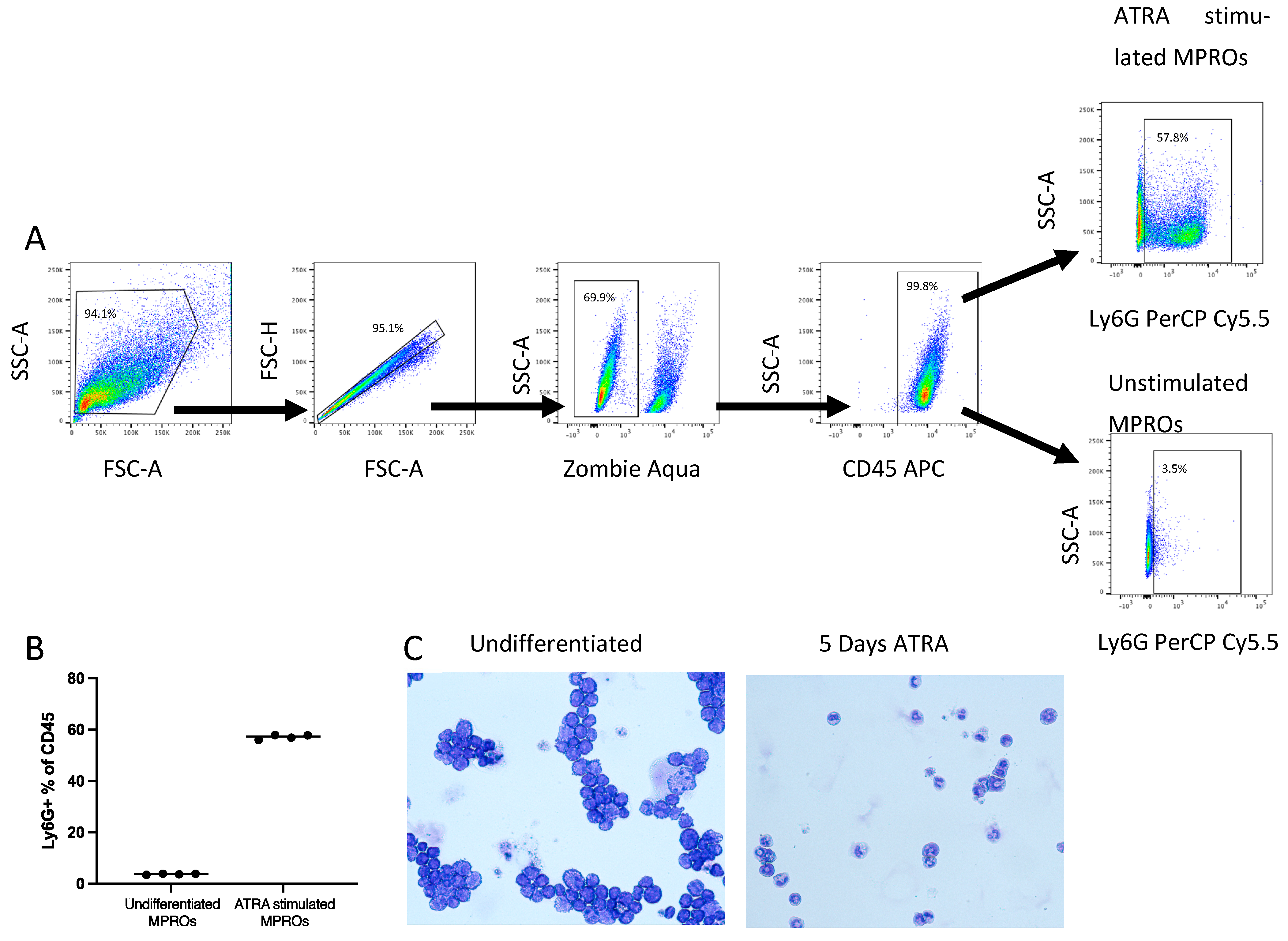
2.3. MPRO Cells Culture and Differentiation
2.4. Primary Murine Neutrophils Isolation
2.5. Phagocytosis Assay and Bactericidal Assay
2.6. ROS Generation via CM-H2DCFDA
2.7. NETs Release
2.8. Assessment of Bactericidal Activity of Neutrophil Supernatants
2.9. Statistical Analysis
3. Results
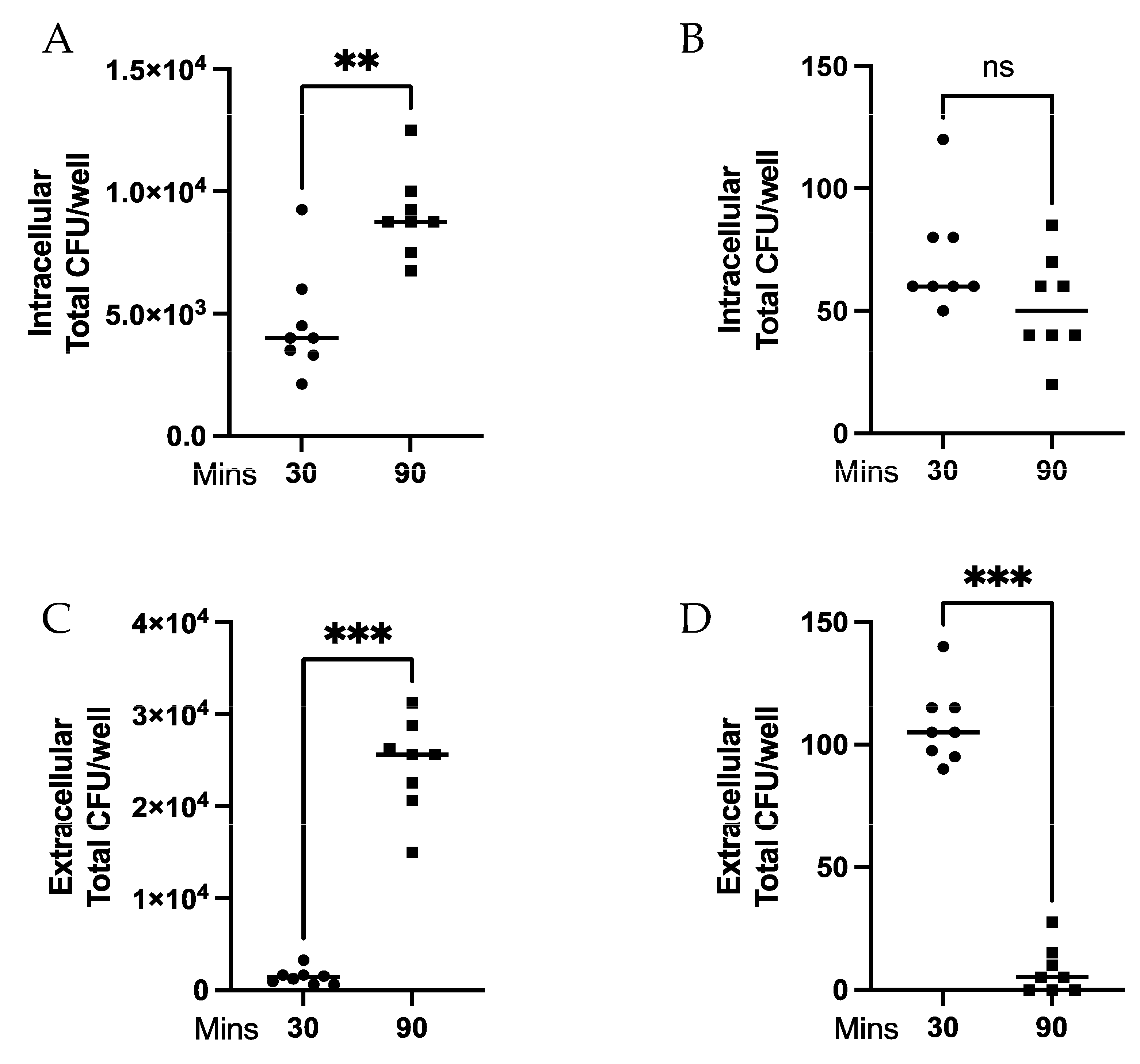
3.1. MPRO Killing of Extracellular but Not Intracellular Spn Is Dependent upon Bacteria:Neutrophil Ratio
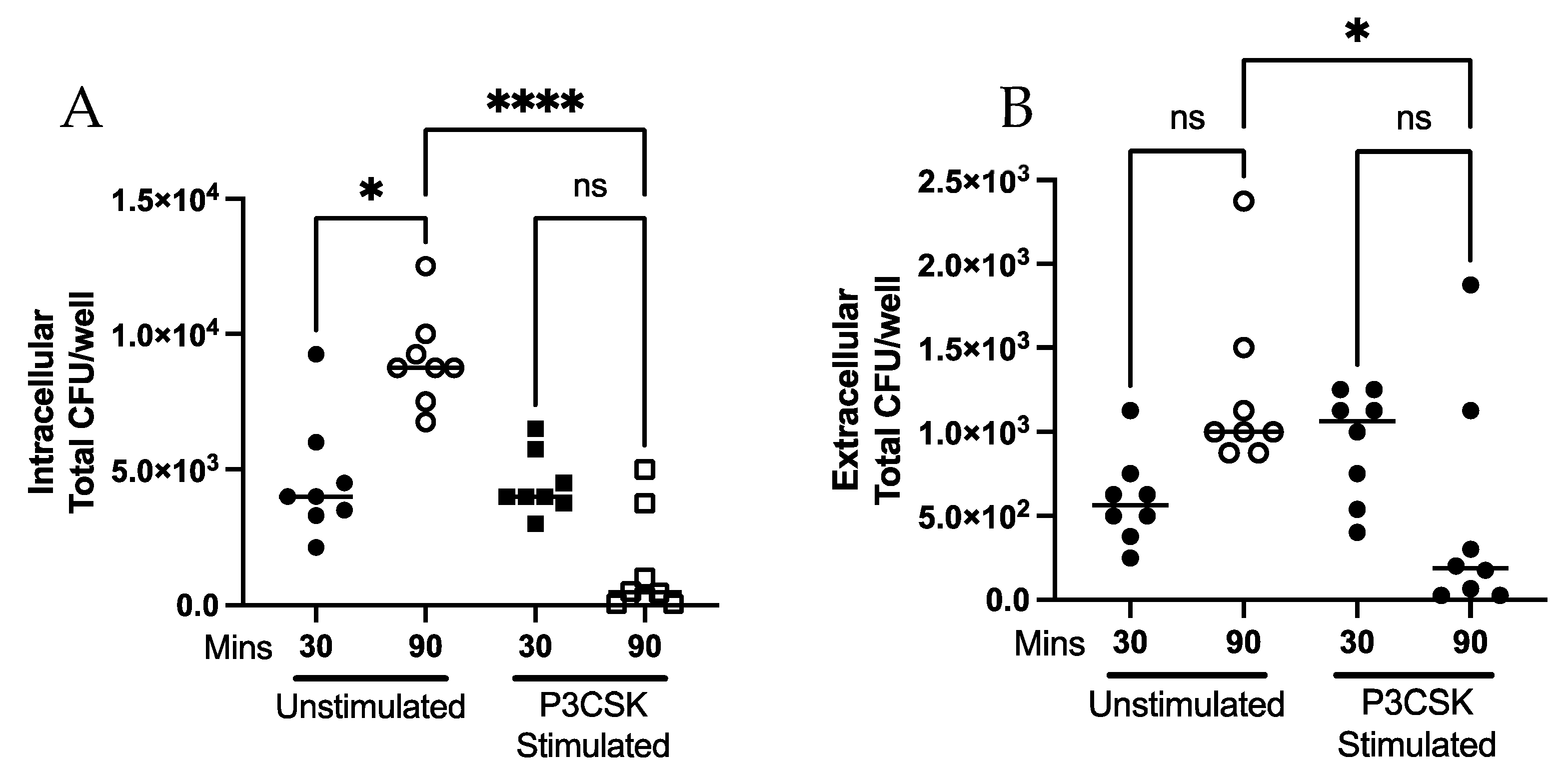
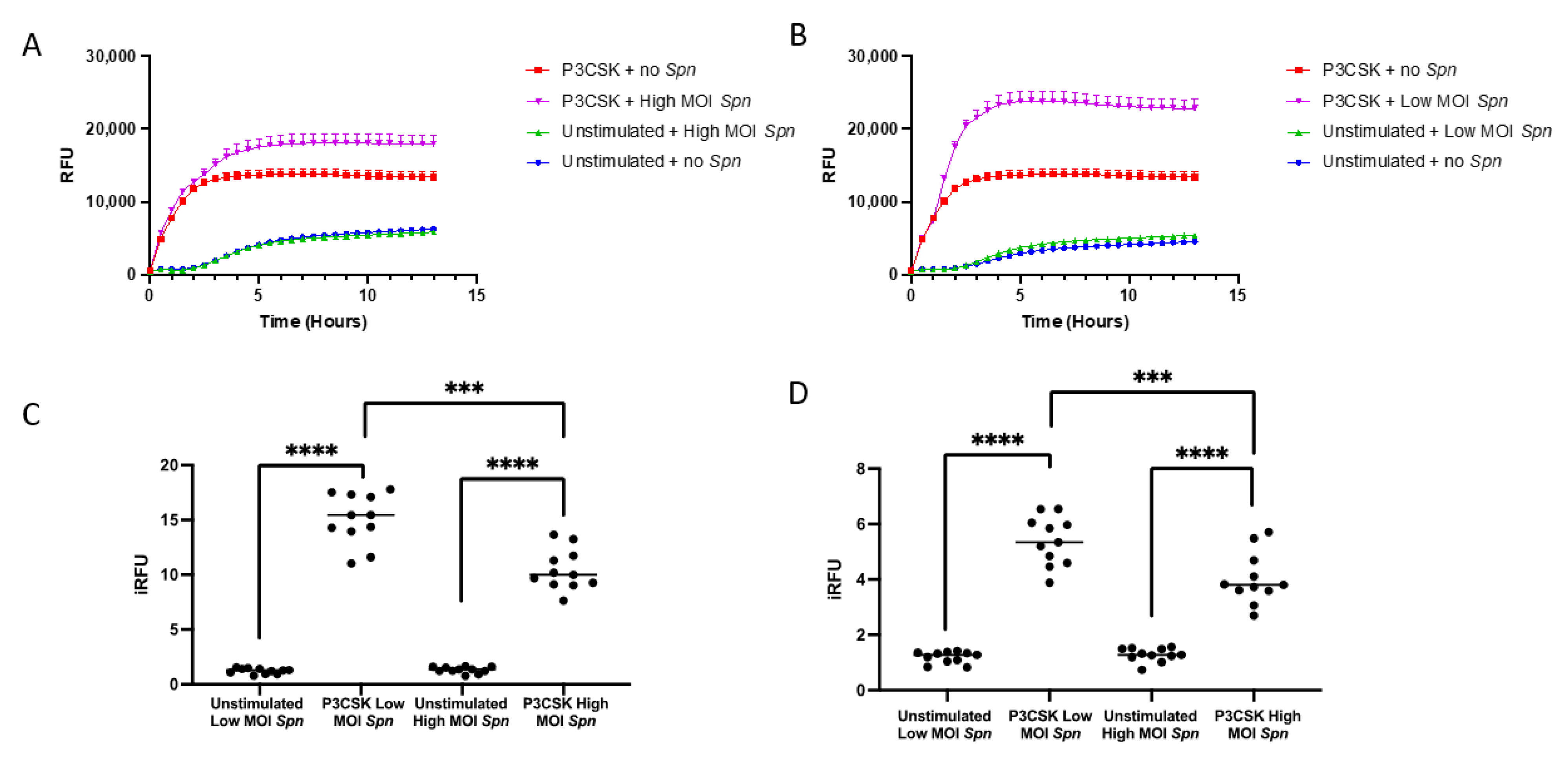
3.2. Ability of MPROs to Kill a Greater Bacterial Challenge, Intra- and Extracellularly, Is Dependent upon TLR2 Activation
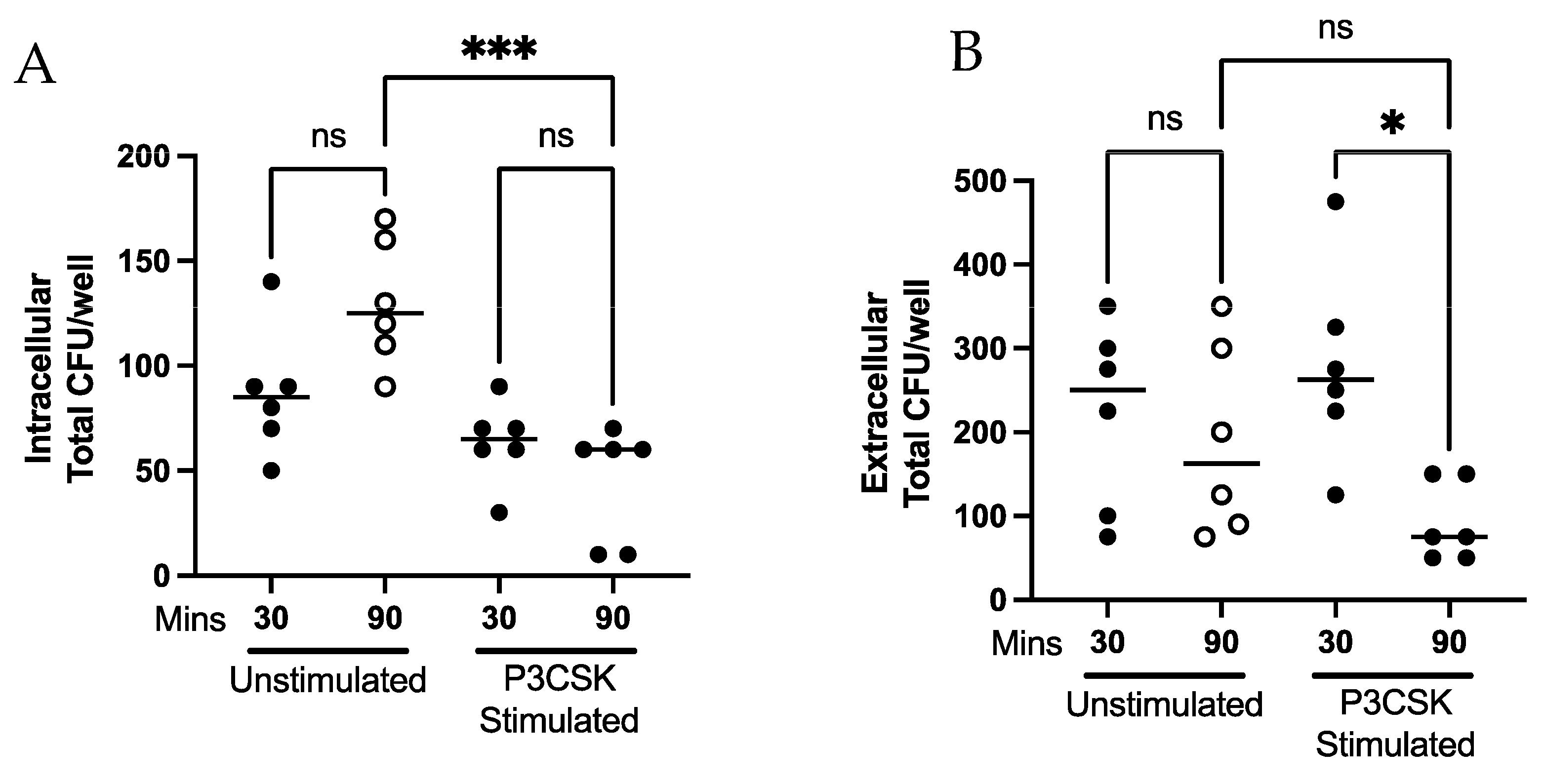
3.3. Ability of Primary Mouse Neutrophils to Contain Intracellular and Kill Extracellular Bacteria Is Dependent upon TLR2 Activation
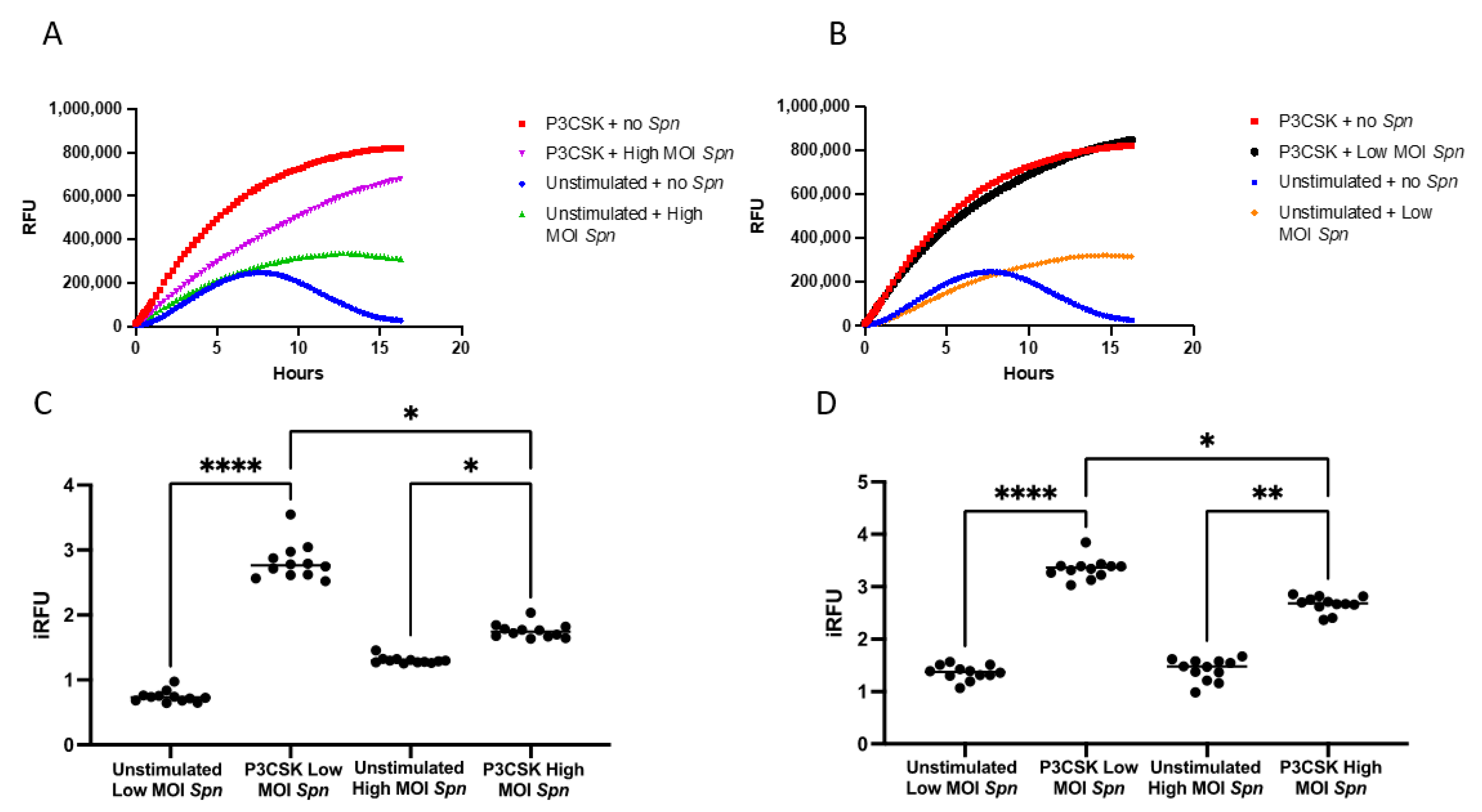
3.4. Primary Mouse Neutrophils Induce ROS Differently When Exposed to Low vs. High MOI of Spn
3.5. Primary Mouse Neutrophils Induce NETs Differently When Exposed to Low vs. High MOI of Spn
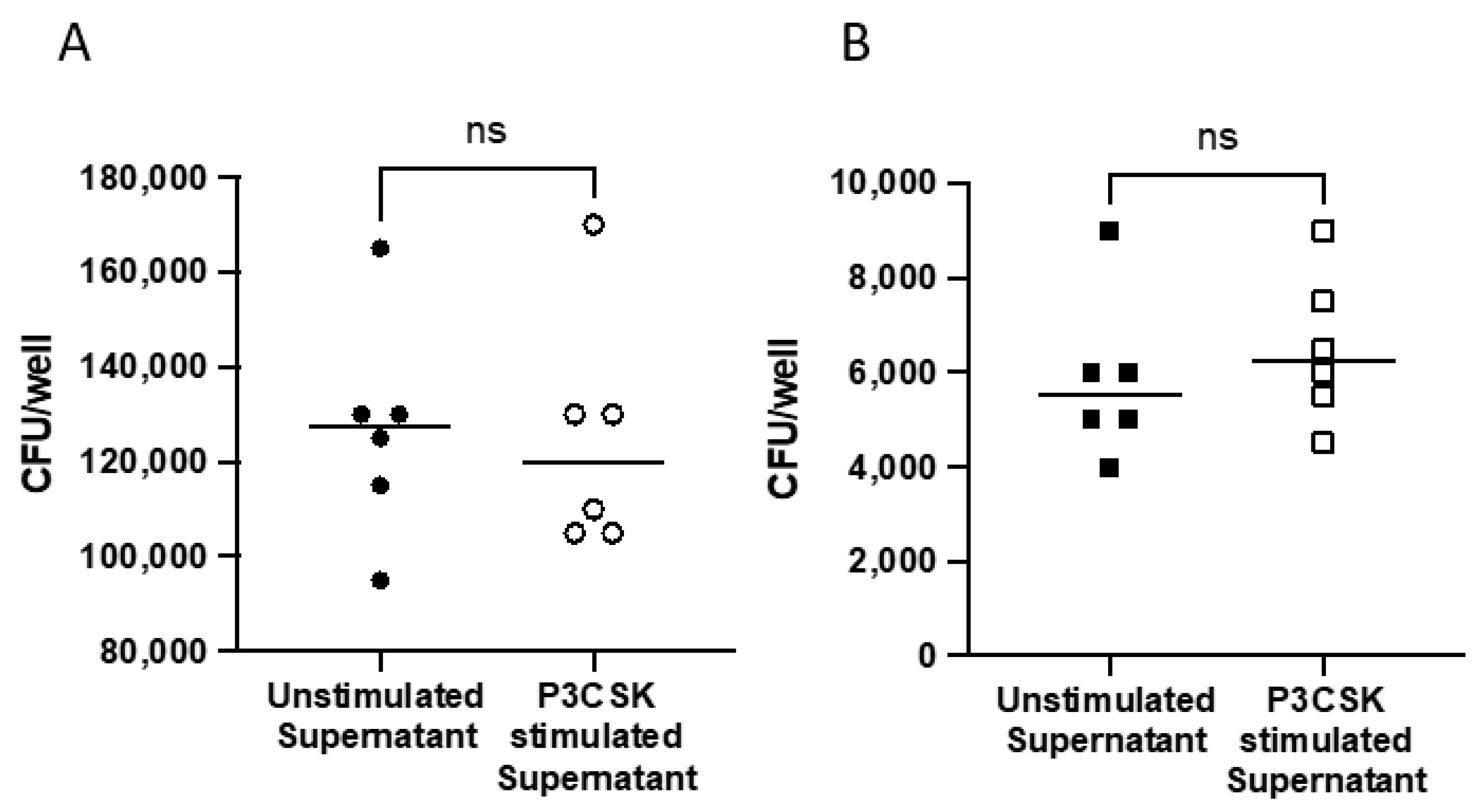
3.6. Extracellular Killing Is Not Mediated via Soluble Factors
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Huang, S.S.; Johnson, K.M.; Ray, G.T.; Wroe, P.; Lieu, T.A.; Moore, M.R.; Zell, E.R.; Linder, J.A.; Grijalva, C.G.; Metlay, J.P.; et al. Healthcare Utilization and Cost of Pneumococcal Disease in the United States. Vaccine 2011, 29, 3398–3412. [Google Scholar] [CrossRef] [Green Version]
- Kim, L.; McGee, L.; Tomczyk, S.; Beall, B. Biological and Epidemiological Features of Antibiotic-Resistant Streptococcus Pneumoniae in Pre- and Post-Conjugate Vaccine Eras: A United States Perspective. Clin. Microbiol. Rev. 2016, 29, 525–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raddaoui, A.; Tanfous, F.B.; Chebbi, Y.; Achour, W.; Baaboura, R.; Benhassen, A. High Prevalence of Multidrug-Resistant International Clones among Macrolide-Resistant Streptococcus Pneumoniae Isolates in Immunocompromised Patients in Tunisia. Int. J. Antimicrob. Agents 2018, 52, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Branzk, N.; Lubojemska, A.; Hardison, S.E.; Wang, Q.; Gutierrez, M.G.; Brown, G.D.; Papayannopoulos, V. Neutrophils Sense Microbe Size and Selectively Release Neutrophil Extracellular Traps in Response to Large Pathogens. Nat. Immunol. 2014, 15, 1017–1025. [Google Scholar] [CrossRef]
- Worley, M.J.; Fei, K.; Lopez-Denman, A.J.; Kelleher, A.D.; Kent, S.J.; Chung, A.W. Neutrophils mediate HIV-specific antibody-dependent phagocytosis and ADCC. J. Immunol. Methods 2018, 457, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Witter, A.R.; Okunnu, B.M.; Berg, R.E. The Essential Role of Neutrophils during Infection with the Intracellular Bacterial Pathogen Listeria Monocytogenes. J. Immunol. 2016, 197, 1557–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balamayooran, G.; Batra, S.; Fessler, M.B.; Happel, K.I.; Jeyaseelan, S. Mechanisms of Neutrophil Accumulation in the Lungs against Bacteria. Am. J. Respir. Cell Mol. Biol. 2010, 43, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Mehta, H.M.; Malandra, M.; Corey, S.J. G-CSF and GM-CSF in Neutropenia. J. Immunol. 2015, 195, 1341–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leijh, P.C.; Barselaar, M.T.; Zwet, T.L.; Dubbeldeman-Rempt, I.; Furth, R. Kinetics of phagocytosis of Staphylococcus aureus and Escherichia coli by human granulocytes. Immunology 1979, 37, 453–465. [Google Scholar]
- Esposito, A.L.; Clark, C.A.; Poirier, W.J. An Assessment of the Factors Contributing to the Killing of Type 3 Streptococcus Pneumoniae by Human Polymorphonuclear Leukocytes in Vitro. Apmis 1990, 98, 111–121. [Google Scholar] [CrossRef]
- Li, Y.; Karlin, A.; Loike, J.D.; Silverstein, S.C. A Critical Concentration of Neutrophils Is Required for Effective Bacterial Killing in Suspension. Proc. Natl. Acad. Sci. USA 2002, 99, 8289–8294. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, F.; Means, T.K.; Luster, A.D. Toll-like Receptors Stimulate Human Neutrophil Function. Blood 2003, 102, 2660–2669. [Google Scholar] [CrossRef] [Green Version]
- Letiembre, M.; Echchannaoui, H.; Bachmann, P.; Ferracin, F.; Nieto, C.; Espinosa, M.; Landmann, R. Toll-like Receptor 2 Deficiency Delays Pneumococcal Phagocytosis and Impairs Oxidative Killing by Granulocytes. Infect. Immun. 2005, 73, 8397–8401. [Google Scholar] [CrossRef] [Green Version]
- Koedel, U.; Angele, B.; Rupprecht, T.; Wagner, H.; Roggenkamp, A.; Pfister, H.W.; Kirschning, C.J. Toll-like receptor 2 participates in mediation of immune response in experimental pneumococcal meningitis. J. Immunol. 2003, 170, 438–444. [Google Scholar] [CrossRef] [Green Version]
- Knapp, S.; Wieland, C.W.; van ’t Veer, C.; Takeuchi, O.; Akira, S.; Florquin, S.; van der Poll, T. Toll-like Receptor 2 Plays a Role in the Early Inflammatory Response to Murine Pneumococcal Pneumonia but Does Not Contribute to Antibacterial Defense. J. Immunol. 2004, 172, 3132–3138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miralda, I.; Uriarte, S.M.; McLeish, K.R. Multiple Phenotypic Changes Define Neutrophil Priming. Front. Cell. Infect. Microbiol. 2017, 7, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, M.S. Molecular Events in the Activation of Human Neutrophils for Microbial Killing. Clin Infect Dis 1994, 18 (Suppl. S2), 170–179. [Google Scholar] [CrossRef]
- Standish, A.J.; Weiser, J.N. Human Neutrophils Kill Streptococcus Pneumoniae via Serine Proteases. J. Immunol. 2009, 183, 2602–2609. [Google Scholar] [CrossRef] [Green Version]
- Azzouz, L.; Cherry, A.; Riedl, M.; Khan, M.; Pluthero, F.G.; Kahr, W.H.A.; Palaniyar, N.; Licht, C. Relative antibacterial functions of complement and NETs: NETs trap and complement effectively kills bacteria. Mol. Immunol. 2018, 97, 71–81. [Google Scholar] [CrossRef]
- Lu, T.; Porter, A.R.; Kennedy, A.D.; Kobayashi, S.D.; DeLeo, F.R. Phagocytosis and Killing of Staphylococcus Aureus by Human Neutrophils. J. Innate Immun. 2014, 6, 639–649. [Google Scholar] [CrossRef]
- Yost, C.C.; Cody, M.J.; Harris, E.S.; Thornton, N.L.; McInturff, A.M.; Martinez, M.L.; Chandler, N.B.; Rodesch, C.K.; Albertine, K.H.; Petti, C.A.; et al. Impaired neutrophil extracellular trap (NET) formation: A novel innate immune deficiency of human neonates. Blood 2009, 113, 6419–6427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazeldine, J.; Harris, P.; Chapple, I.L.; Grant, M.; Greenwood, H.; Livesey, A.; Sapey, E.; Lord, J.M. Impaired Neutrophil Extracellular Trap Formation: A Novel Defect in the Innate Immune System of Aged Individuals. Aging Cell 2014, 13, 690–698. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Pilsczek, F.H.; Salina, D.; Poon, K.K.; Fahey, C.; Yipp, B.G.; Sibley, C.D.; Robbins, S.M.; Green, F.H.; Surette, M.G.; Sugai, M.; et al. A novel mechanism of rapid nuclear neutrophil extracellular trap formation in response to Staphylococcus aureus. J. Immunol. 2010, 185, 7413–7425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenny, E.F.; Herzig, A.; Kruger, R.; Muth, A.; Mondal, S.; Thompson, P.R.; Brinkmann, V.; Bernuth, H.V.; Zychlinsky, A. Diverse stimuli engage different neutrophil extracellular trap pathways. eLife 2017, 6, e24437. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.G.; Zhang, Y.; Deng, L.Q.; Chen, H.; Zhang, Y.J.; Zhou, N.J.; Yuan, K.; Yu, L.Z.; Xiong, Z.H.; Gui, X.M.; et al. Control of Methicillin-Resistant Staphylococcus aureus Pneumonia Utilizing TLR2 Agonist Pam3CSK4. PLoS ONE 2016, 11, 0149233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, N.D.; Krause, D.S.; Berliner, N. Normal neutrophil differentiation and secondary granule gene expression in the EML and MPRO cell lines. Exp. Hematol. 1998, 26, 1178–1185. [Google Scholar]
- Gaines, P.; Chi, J.; Berliner, N. Heterogeneity of Functional Responses in Differentiated Myeloid Cell Lines Reveals EPRO Cells as a Valid Model of Murine Neutrophil Functional Activation. J. Leukoc. Biol. 2005, 77, 669–679. [Google Scholar] [CrossRef]
- Tsai, S.; Collins, S.J. A Dominant Negative Retinoic Acid Receptor Blocks Neutrophil Differentiation at the Promyelocyte Stage. Proc. Natl. Acad. Sci. USA 1993, 90, 7153–7157. [Google Scholar] [CrossRef]
- Gong, H.H.; Worley, M.J.; Carver, K.A.; Goldstein, D.R.; Deng, J.C. Neutrophils Drive Pulmonary Vascular Leakage in MHV-1 Infection of Susceptible A/J Mice. Front. Immunol. 2022, 13, 1089064. [Google Scholar] [CrossRef] [PubMed]
- Mullarkey, C.E.; Bailey, M.J.; Golubeva, D.A.; Tan, G.S.; Nachbagauer, R.; He, W.; Novakowski, K.E.; Bowdish, D.M.; Miller, M.S.; Palese, P. Broadly Neutralizing Hemagglutinin Stalk-Specific Antibodies Induce Potent Phagocytosis of Immune Complexes by Neutrophils in an Fc-Dependent Manner. mBio 2016, 7, 10–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llewellyn, D.; Cassan, S.C.; Williams, A.R.; Douglas, A.D.; Forbes, E.K.; Adame-Gallegos, J.R.; Shi, J.; Pleass, R.J.; Draper, S.J. Assessment of Antibody-Dependent Respiratory Burst Activity from Mouse Neutrophils on Plasmodium Yoelii Malaria Challenge Outcome. J. Leukoc. Biol. 2014, 95, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Ip, W.K.; Takahashi, K.; Moore, K.J.; Stuart, L.M.; Ezekowitz, R.A. Mannose-Binding Lectin Enhances Toll-like Receptors 2 and 6 Signaling from the Phagosome. J. Exp. Med. 2008, 205, 169–181. [Google Scholar] [CrossRef]
- Torres, D.; Barrier, M.; Bihl, F.; Quesniaux, V.J.; Maillet, I.; Akira, S.; Ryffel, B.; Erard, F. Toll-like Receptor 2 Is Required for Optimal Control of Listeria Monocytogenes Infection. Infect. Immun. 2004, 72, 2131–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assinger, A.; Laky, M.; Schabbauer, G.; Hirschl, A.M.; Buchberger, E.; Binder, B.R.; Volf, I. Efficient Phagocytosis of Periodontopathogens by Neutrophils Requires Plasma Factors, Platelets and TLR2. J. Thromb. Haemost. 2011, 9, 799–809. [Google Scholar] [CrossRef]
- Fang, L.; Wu, H.M.; Ding, P.S.; Liu, R.Y. TLR2 Mediates Phagocytosis and Autophagy through JNK Signaling Pathway in Staphylococcus Aureus-Stimulated RAW264.7 Cells. Cell. Signal. 2014, 26, 806–814. [Google Scholar] [CrossRef]
- Delgado-Rizo, V.; Martinez-Guzman, M.A.; Iniguez-Gutierrez, L.; Garcia-Orozco, A.; Alvarado-Navarro, A.; Fafutis-Morris, M. Neutrophil Extracellular Traps and Its Implications in Inflammation: An Overview. Front. Immunol. 2017, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- Awasthi, D.; Nagarkoti, S.; Kumar, A.; Dubey, M.; Singh, A.K.; Pathak, P.; Chandra, T.; Barthwal, M.K.; Dikshit, M. Oxidized LDL Induced Extracellular Trap Formation in Human Neutrophils via TLR-PKC-IRAK-MAPK and NADPH-Oxidase Activation. Free Radic. Biol. Med. 2016, 93, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Tamarozzi, F.; Turner, J.D.; Pionnier, N.; Midgley, A.; Guimaraes, A.F.; Johnston, K.L.; Edwards, S.W.; Taylor, M.J. Wolbachia Endosymbionts Induce Neutrophil Extracellular Trap Formation in Human Onchocerciasis. Sci. Rep. 2016, 6, 35559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, F.; Chang, X.; Wang, G.; Zhou, H.; Ma, Z.; Lin, H.; Fan, H. Streptococcus Suis Serotype 2 Stimulates Neutrophil Extracellular Traps Formation via Activation of P38 MAPK and ERK1/2. Front. Immunol. 2018, 9, 2854. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, Z.; Carver, K.A.; Gong, H.H.; Kosai, K.; Deng, J.C.; Worley, M.J. Detailed Mechanisms Underlying Neutrophil Bactericidal Activity against Streptococcus pneumoniae. Biomedicines 2023, 11, 2252. https://doi.org/10.3390/biomedicines11082252
Tsai Z, Carver KA, Gong HH, Kosai K, Deng JC, Worley MJ. Detailed Mechanisms Underlying Neutrophil Bactericidal Activity against Streptococcus pneumoniae. Biomedicines. 2023; 11(8):2252. https://doi.org/10.3390/biomedicines11082252
Chicago/Turabian StyleTsai, Zachary, Kyle A. Carver, Henry H. Gong, Kosuke Kosai, Jane C. Deng, and Matthew J. Worley. 2023. "Detailed Mechanisms Underlying Neutrophil Bactericidal Activity against Streptococcus pneumoniae" Biomedicines 11, no. 8: 2252. https://doi.org/10.3390/biomedicines11082252