
Colitis Is Associated with Loss of the Histidine Phosphatase LHPP and Upregulation of Histidine Phosphorylation in Intestinal Epithelial Cells
 and
and 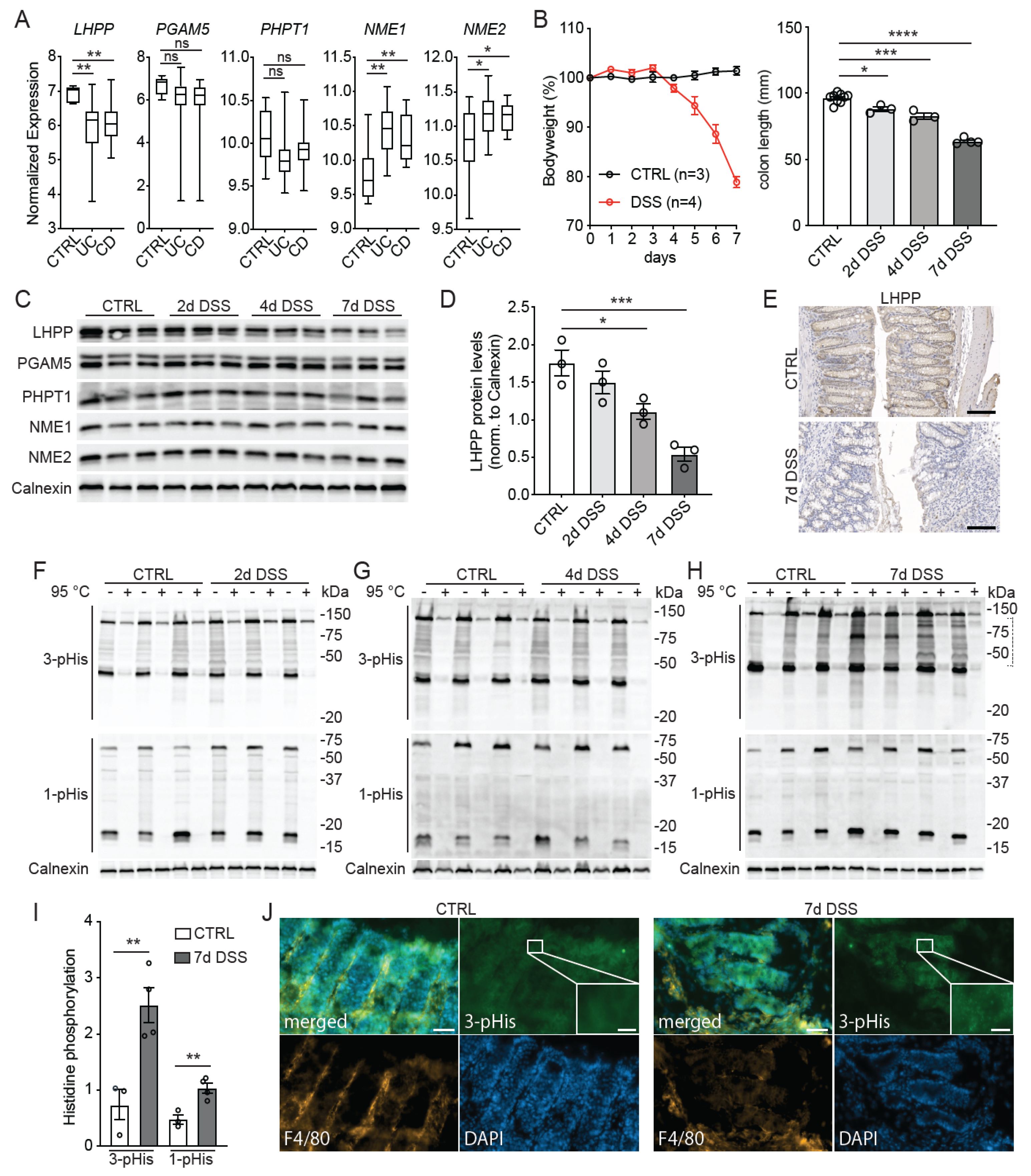{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Histology
2.3. Immunoblotting
2.4. Analysis of the Publicly Available Transcriptomic Dataset
2.5. Statistical Analysis
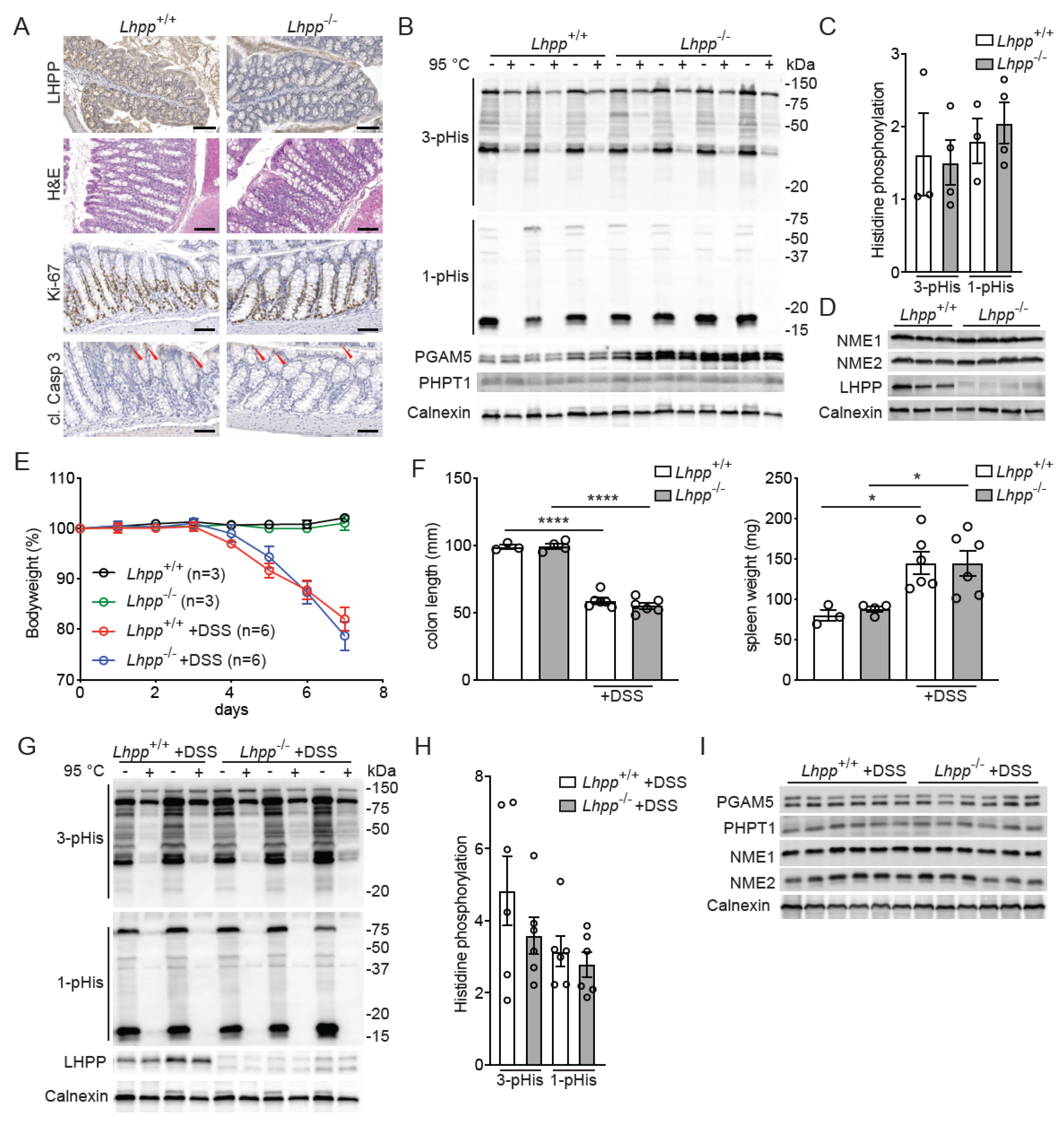
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuhs, S.R.; Hunter, T. pHisphorylation: The emergence of histidine phosphorylation as a reversible regulatory modification. Curr. Opin. Cell Biol. 2017, 45, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Fuhs, S.R.; Meisenhelder, J.; Aslanian, A.; Ma, L.; Zagorska, A.; Stankova, M.; Binnie, A.; Al-Obeidi, F.; Mauger, J.; Lemke, G.; et al. Monoclonal 1- and 3-Phosphohistidine Antibodies: New Tools to Study Histidine Phosphorylation. Cell 2015, 162, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Hindupur, S.K.; Colombi, M.; Fuhs, S.R.; Matter, M.S.; Guri, Y.; Adam, K.; Cornu, M.; Piscuoglio, S.; Ng, C.K.Y.; Betz, C.; et al. The protein histidine phosphatase LHPP is a tumour suppressor. Nature 2018, 555, 678–682. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Chen, Y.; Zhu, J. LHPP suppresses proliferation, migration, and invasion and promotes apoptosis in pancreatic cancer. Biosci. Rep. 2020, 40, BSR20194142. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.J.; Chen, L.H.; Wang, J.; Wang, Z.T.; Wu, C.Y.; Sun, K.; Ding, B.Y.; Liu, N.; Xu, R.X. LHPP impedes energy metabolism by inducing ubiquitin-mediated degradation of PKM2 in glioblastoma. Am. J. Cancer Res. 2021, 11, 1369–1390. [Google Scholar] [PubMed]
- Liu, S.; Gao, W.; Lu, Y.; Zhou, Q.; Su, R.; Hasegawa, T.; Du, J.; Li, M. As a Novel Tumor Suppressor, LHPP Promotes Apoptosis by Inhibiting the PI3K/AKT Signaling Pathway in Oral Squamous Cell Carcinoma. Int. J. Biol. Sci. 2022, 18, 491–506. [Google Scholar] [CrossRef] [PubMed]
- Hou, B.; Li, W.; Li, J.; Ma, J.; Xia, P.; Liu, Z.; Zeng, Q.; Zhang, X.; Chang, D. Tumor suppressor LHPP regulates the proliferation of colorectal cancer cells via the PI3K/AKT pathway. Oncol. Rep. 2020, 43, 536–548. [Google Scholar] [CrossRef]
- Percario, R.; Panaccio, P.; di Mola, F.F.; Grottola, T.; Di Sebastiano, P. The Complex Network between Inflammation and Colorectal Cancer: A Systematic Review of the Literature. Cancers 2021, 13, 6237. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.R.; Bakir, I.A.; Hart, A.L.; Graham, T.A. Clonal evolution of colorectal cancer in IBD. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 218–229. [Google Scholar] [CrossRef] [PubMed]
- de Souza, H.S.P.; Fiocchi, C.; Iliopoulos, D. The IBD interactome: An integrated view of aetiology, pathogenesis and therapy. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 739–749. [Google Scholar] [CrossRef]
- Di, L.; Srivastava, S.; Zhdanova, O.; Ding, Y.; Li, Z.; Wulff, H.; Lafaille, M.; Skolnik, E.Y. Inhibition of the K+ channel KCa3.1 ameliorates T cell-mediated colitis. Proc. Natl. Acad. Sci. USA 2010, 107, 1541–1546. [Google Scholar] [CrossRef]
- Panda, S.; Srivastava, S.; Li, Z.; Vaeth, M.; Fuhs, S.R.; Hunter, T.; Skolnik, E.Y. Identification of PGAM5 as a Mammalian Protein Histidine Phosphatase that Plays a Central Role to Negatively Regulate CD4(+) T Cells. Mol. Cell 2016, 63, 457–469. [Google Scholar] [CrossRef] [Green Version]
- Concordet, J.P.; Haeussler, M. CRISPOR: Intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic Acids Res. 2018, 46, W242–W245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Kutny, P.M.; Byers, S.L.; Longstaff, C.J.; DaCosta, M.J.; Pang, C.; Zhang, Y.; Taft, R.A.; Buaas, F.W.; Wang, H. Delivery of Cas9 Protein into Mouse Zygotes through a Series of Electroporation Dramatically Increases the Efficiency of Model Creation. J. Genet. Genom. 2016, 43, 319–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirtz, S.; Popp, V.; Kindermann, M.; Gerlach, K.; Weigmann, B.; Fichtner-Feigl, S.; Neurath, M.F. Chemically induced mouse models of acute and chronic intestinal inflammation. Nat. Protoc. 2017, 12, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.; Glitzner, E.; Srivatsa, S.; Bakiri, L.; Matsuoka, K.; Shahrouzi, P.; Dumanic, M.; Novoszel, P.; Mohr, T.; Langer, O.; et al. EGFR is required for FOS-dependent bone tumor development via RSK2/CREB signaling. EMBO Mol. Med. 2018, 10, e9408. [Google Scholar] [CrossRef]
- Arijs, I.; De Hertogh, G.; Lemaire, K.; Quintens, R.; Van Lommel, L.; Van Steen, K.; Leemans, P.; Cleynen, I.; Van Assche, G.; Vermeire, S.; et al. Mucosal gene expression of antimicrobial peptides in inflammatory bowel disease before and after first infliximab treatment. PLoS ONE 2009, 4, e7984. [Google Scholar] [CrossRef] [PubMed]
- Laroui, H.; Ingersoll, S.A.; Liu, H.C.; Baker, M.T.; Ayyadurai, S.; Charania, M.A.; Laroui, F.; Yan, Y.; Sitaraman, S.V.; Merlin, D. Dextran sodium sulfate (DSS) induces colitis in mice by forming nano-lipocomplexes with medium-chain-length fatty acids in the colon. PLoS ONE 2012, 7, e32084. [Google Scholar] [CrossRef]
- Srivatsa, S.; Paul, M.C.; Cardone, C.; Holcmann, M.; Amberg, N.; Pathria, P.; Diamanti, M.A.; Linder, M.; Timelthaler, G.; Dienes, H.P.; et al. EGFR in Tumor-Associated Myeloid Cells Promotes Development of Colorectal Cancer in Mice and Associates with Outcomes of Patients. Gastroenterology 2017, 153, 178–190.e10. [Google Scholar] [CrossRef] [Green Version]
- Eaden, J.A.; Abrams, K.R.; Mayberry, J.F. The risk of colorectal cancer in ulcerative colitis: A meta-analysis. Gut 2001, 48, 526–535. [Google Scholar] [CrossRef] [Green Version]
- Lutgens, M.W.; Vleggaar, F.P.; Schipper, M.E.; Stokkers, P.C.; van der Woude, C.J.; Hommes, D.W.; de Jong, D.J.; Dijkstra, G.; van Bodegraven, A.A.; Oldenburg, B.; et al. High frequency of early colorectal cancer in inflammatory bowel disease. Gut 2008, 57, 1246–1251. [Google Scholar] [CrossRef] [PubMed]
- Hou, B.; Li, W.; Xia, P.; Zhao, F.; Liu, Z.; Zeng, Q.; Wang, S.; Chang, D. LHPP suppresses colorectal cancer cell migration and invasion in vitro and in vivo by inhibiting Smad3 phosphorylation in the TGF-beta pathway. Cell Death Discov. 2021, 7, 273. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, X.; Zhu, H.; Song, X.; Gao, H.; Niu, Z.; Lu, J. Purpurin binding interacts with LHPP protein that inhibits PI3K/AKT phosphorylation and induces apoptosis in colon cancer cells HCT-116. J. Biochem. Mol. Toxicol. 2021, 35, e22665. [Google Scholar] [CrossRef]
- Guo, C.; Liu, X.; Xu, Y.; Han, X.; Xie, R.; Meng, X.; Li, Y.; Chen, T.; Cheng, Z.; Fu, X. Exploring the Mechanism of Action of Canmei Formula against Colorectal Adenoma through Multi-Omics Technique. Front. Cell Dev. Biol. 2021, 9, 778826. [Google Scholar] [CrossRef]
- Cader, M.Z.; Kaser, A. Recent advances in inflammatory bowel disease: Mucosal immune cells in intestinal inflammation. Gut 2013, 62, 1653–1664. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linder, M.; Liko, D.; Kancherla, V.; Piscuoglio, S.; Hall, M.N. Colitis Is Associated with Loss of the Histidine Phosphatase LHPP and Upregulation of Histidine Phosphorylation in Intestinal Epithelial Cells. Biomedicines 2023, 11, 2158. https://doi.org/10.3390/biomedicines11082158
Linder M, Liko D, Kancherla V, Piscuoglio S, Hall MN. Colitis Is Associated with Loss of the Histidine Phosphatase LHPP and Upregulation of Histidine Phosphorylation in Intestinal Epithelial Cells. Biomedicines. 2023; 11(8):2158. https://doi.org/10.3390/biomedicines11082158
Chicago/Turabian StyleLinder, Markus, Dritan Liko, Venkatesh Kancherla, Salvatore Piscuoglio, and Michael N. Hall. 2023. "Colitis Is Associated with Loss of the Histidine Phosphatase LHPP and Upregulation of Histidine Phosphorylation in Intestinal Epithelial Cells" Biomedicines 11, no. 8: 2158. https://doi.org/10.3390/biomedicines11082158