
Oxidative Stress Markers and Heat Shock Proteins in Non-Obese Women with Polycystic Ovary Syndrome Are Not Elevated and Show No Correlation with Vitamin D
 ,
,
Abstract
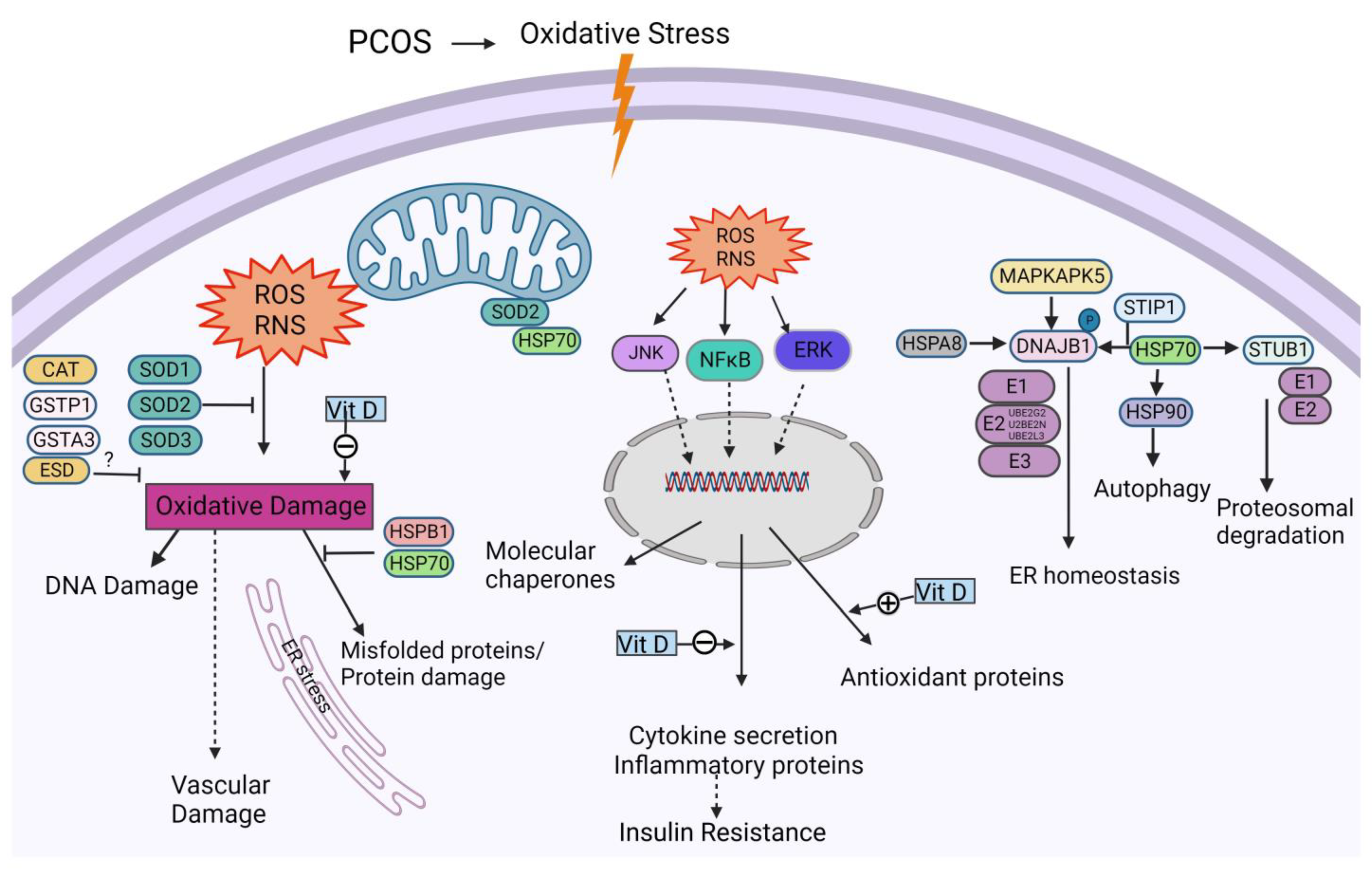
:1. Introduction
2. Materials and Methods
Statistics
3. Results
3.1. Demographic Data
3.2. Oxidative Stress Results
3.3. Heat Shock Protein Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shroff, R.; Kerchner, A.; Maifeld, M.; Van Beek, E.J.R.; Jagasia, D.; Dokras, A. Young obese women with polycystic ovary syndrome have evidence of early coronary atherosclerosis. J. Clin. Endocrinol. Metab. 2007, 92, 4609–4614. [Google Scholar] [CrossRef] [PubMed]
- Legro, R.S.; Arslanian, S.A.; Ehrmann, D.A.; Hoeger, K.M.; Murad, M.H.; Pasquali, R.; Welt, C.K. Diagnosis and treatment of polycystic ovary syndrome: An Endocrine Society clinical practice guideline. J. Clin. Endocrinol. Metab. 2013, 98, 4565–4592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamanti-Kandarakis, E.; Kouli, C.R.; Bergiele, A.T.; Filandra, F.A.; Tsianateli, T.C.; Spina, G.G.; Zapanti, E.D.; Bartzis, M.I. A survey of the polycystic ovary syndrome in the Greek island of Lesbos: Hormonal and metabolic profile. J. Clin. Endocrinol. Metab. 1999, 84, 4006–4011. [Google Scholar] [CrossRef] [PubMed]
- Sathyapalan, T.; Atkin, S.L. Recent advances in cardiovascular aspects of polycystic ovary syndrome. Eur. J. Endocrinol. 2012, 166, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, M. Oxidative Stress and Polycystic Ovary Syndrome: A Brief Review. Int. J. Prev. Med. 2019, 10, 86. [Google Scholar] [CrossRef]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [Green Version]
- Sathyapalan, T.; Shepherd, J.; Coady, A.M.; Kilpatrick, E.S.; Atkin, S.L. Atorvastatin reduces malondialdehyde concentrations in patients with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2012, 97, 3951–3955. [Google Scholar] [CrossRef] [Green Version]
- Bannigida, D.M.; Nayak, B.S.; Vijayaraghavan, R. Insulin resistance and oxidative marker in women with PCOS. Arch. Physiol. Biochem. 2020, 126, 183–186. [Google Scholar] [CrossRef]
- Niinuma, S.A.; Lubbad, L.; Lubbad, W.; Moin, A.S.M.; Butler, A.E. The Role of Heat Shock Proteins in the Pathogenesis of Polycystic Ovarian Syndrome: A Review of the Literature. Int. J. Mol. Sci. 2023, 24, 1838. [Google Scholar] [CrossRef]
- Moin, A.S.M.; Sathyapalan, T.; Butler, A.E.; Atkin, S.L. Coagulation factor dysregulation in polycystic ovary syndrome is an epiphenomenon of obesity. Clin. Endocrinol. 2023, 98, 796–802. [Google Scholar] [CrossRef]
- Lewis, R.D.; Narayanaswamy, A.K.; Farewell, D.; Rees, D.A. Complement activation in polycystic ovary syndrome occurs in the postprandial and fasted state and is influenced by obesity and insulin sensitivity. Clin. Endocrinol. 2021, 94, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, Q.; Song, Y.; Tian, B.; Cheng, Q.; Qing, H.; Zhong, L.; Xia, W. Serum complement C3 has a stronger association with insulin resistance than high-sensitivity C-reactive protein in women with polycystic ovary syndrome. Fertil. Steril. 2011, 95, 1749–1753. [Google Scholar] [CrossRef]
- Moin, A.S.M.; Sathyapalan, T.; Butler, A.E.; Atkin, S.L. Classical and alternate complement factor overexpression in non-obese weight matched women with polycystic ovary syndrome does not correlate with vitamin D. Front. Endocrinol. 2022, 13, 935750. [Google Scholar] [CrossRef]
- Butler, A.E.; Moin, A.S.M.; Sathyapalan, T.; Atkin, S.L. Components of the Complement Cascade Differ in Polycystic Ovary Syndrome. Int. J. Mol. Sci. 2022, 23, 12232. [Google Scholar] [CrossRef] [PubMed]
- Legro, R.S. Obesity and PCOS: Implications for diagnosis and treatment. In Seminars in Reproductive Medicine; Thieme Medical Publishers: New York, NY, USA, 2012; Volume 30, pp. 496–506. [Google Scholar]
- Hu, L.; Ma, L.; Xia, X.; Ying, T.; Zhou, M.; Zou, S.; Yu, H.; Yin, J. Efficacy of Bariatric Surgery in the Treatment of Women with Obesity and Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2022, 107, e3217–e3229. [Google Scholar] [CrossRef] [PubMed]
- Alalami, H.; Sathyapalan, T.; Atkin, S.L. Cardiovascular profile of pharmacological agents used for the management of polycystic ovary syndrome. Ther. Adv. Endocrinol. Metab. 2019, 10, 2042018818805674. [Google Scholar] [CrossRef] [PubMed]
- Bastard, J.-P.; Jardel, C.; Bruckert, E.; Blondy, P.; Capeau, J.; Laville, M.; Vidal, H.; Hainque, B. Elevated levels of interleukin 6 are reduced in serum and subcutaneous adipose tissue of obese women after weight loss. J. Clin. Endocrinol. Metab. 2000, 85, 3338–3342. [Google Scholar]
- O’Rourke, R.W.; Kay, T.; Lyle, E.A.; Traxler, S.A.; Deveney, C.W.; Jobe, B.A.; Roberts, C.T.; Marks, D.; Rosenbaum, J.T. Alterations in peripheral blood lymphocyte cytokine expression in obesity. Clin. Exp. Immunol. 2006, 146, 39–46. [Google Scholar] [CrossRef]
- Festa, A.; D’Agostino, R., Jr.; Williams, K.; Karter, A.J.; Mayer-Davis, E.J.; Tracy, R.P.; Haffner, S.M. The relation of body fat mass and distribution to markers of chronic inflammation. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 1407–1415. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.C.; Lyall, H.; Petrie, J.R.; Gould, G.W.; Connell, J.M.; Sattar, N. Low grade chronic inflammation in women with polycystic ovarian syndrome. J. Clin. Endocrinol. Metab. 2001, 86, 2453–2455. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Shah, N.; Deshmukh, H.; Sahebkar, A.; Östlundh, L.; Al-Rifai, R.H.; Atkin, S.L.; Sathyapalan, T. Effect of pharmacological interventions on lipid profiles and C-reactive protein in polycystic ovary syndrome: A systematic review and meta-analysis. Clin. Endocrinol. 2022, 96, 443–459. [Google Scholar] [CrossRef]
- Ridker, P.M.; Hennekens, C.H.; Buring, J.E.; Rifai, N. C-reactive protein and other markers of inflammation in the prediction of cardiovascular disease in women. N. Engl. J. Med. 2000, 342, 836–843. [Google Scholar] [CrossRef]
- Fauser, B.C.; Tarlatzis, B.C.; Rebar, R.W.; Legro, R.S.; Balen, A.H.; Lobo, R.; Carmina, E.; Chang, J.; Yildiz, B.O.; Laven, J.S.; et al. Consensus on women’s health aspects of polycystic ovary syndrome (PCOS): The Amsterdam ESHRE/ASRM-Sponsored 3rd PCOS Consensus Workshop Group. Fertil. Steril. 2012, 97, 28–38.e25. [Google Scholar] [CrossRef]
- Setji, T.L.; Holland, N.D.; Sanders, L.L.; Pereira, K.C.; Diehl, A.M.; Brown, A.J. Nonalcoholic steatohepatitis and nonalcoholic Fatty liver disease in young women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 1741–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baptiste, C.G.; Battista, M.C.; Trottier, A.; Baillargeon, J.P. Insulin and hyperandrogenism in women with polycystic ovary syndrome. J. Steroid Biochem. Mol. Biol. 2010, 122, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Chantrapanichkul, P.; Indhavivadhana, S.; Wongwananuruk, T.; Techatraisak, K.; Dangrat, C.; Sa-Nga-Areekul, N. Prevalence of type 2 diabetes mellitus compared between lean and overweight/obese patients with polycystic ovarian syndrome: A 5-year follow-up study. Arch. Gynecol. Obstet. 2020, 301, 809–816. [Google Scholar] [CrossRef]
- Osibogun, O.; Ogunmoroti, O.; Michos, E.D. Polycystic ovary syndrome and cardiometabolic risk: Opportunities for cardiovascular disease prevention. Trends Cardiovasc. Med. 2020, 30, 399–404. [Google Scholar] [CrossRef]
- Murri, M.; Luque-Ramírez, M.; Insenser, M.; Ojeda-Ojeda, M.; Escobar-Morreale, H.F. Circulating markers of oxidative stress and polycystic ovary syndrome (PCOS): A systematic review and meta-analysis. Hum. Reprod. Update 2013, 19, 268–288. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative Stress in Atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 42. [Google Scholar] [CrossRef]
- Liu, J.F.; Chen, P.C.; Ling, T.Y.; Hou, C.H. Hyperthermia increases HSP production in human PDMCs by stimulating ROS formation, p38 MAPK and Akt signaling, and increasing HSF1 activity. Stem Cell Res. Ther. 2022, 13, 236. [Google Scholar] [CrossRef]
- Tsan, M.F.; Gao, B. Heat shock protein and innate immunity. Cell. Mol. Immunol. 2004, 1, 274–279. [Google Scholar] [PubMed]
- Gao, H.; Meng, J.; Xu, M.; Zhang, S.; Ghose, B.; Liu, J.; Yao, P.; Yan, H.; Wang, D.; Liu, L. Serum Heat Shock Protein 70 Concentration in Relation to Polycystic Ovary Syndrome in a Non-Obese Chinese Population. PLoS ONE 2013, 8, e67727. [Google Scholar] [CrossRef] [PubMed]
- Costes, S.; Huang, C.-J.; Gurlo, T.; Daval, M.; Matveyenko, A.V.; Rizza, R.A.; Butler, A.E.; Butler, P.C. β-cell dysfunctional ERAD/ubiquitin/proteasome system in type 2 diabetes mediated by islet amyloid polypeptide-induced UCH-L1 deficiency. Diabetes 2011, 60, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigouroux, S.; Briand, M.; Briand, Y. Linkage between the proteasome pathway and neurodegenerative diseases and aging. Mol. Neurobiol. 2004, 30, 201–221. [Google Scholar] [CrossRef]
- Hooper, P.L.; Hooper, P.L. Inflammation, heat shock proteins, and type 2 diabetes. Cell Stress Chaperones 2009, 14, 113–115. [Google Scholar] [CrossRef] [Green Version]
- Qi, D.; Liu, H.; Niu, J.; Fan, X.; Wen, X.; Du, Y.; Mou, J.; Pei, D.; Liu, Z.; Zong, Z.; et al. Heat shock protein 72 inhibits c-Jun N-terminal kinase 3 signaling pathway via Akt1 during cerebral ischemia. J. Neurol. Sci. 2012, 317, 123–129. [Google Scholar] [CrossRef]
- Hahn, S.; Haselhorst, U.; Tan, S.; Quadbeck, B.; Schmidt, M.; Roesler, S.; Kimmig, R.; Mann, K.; Janssen, O. Low serum 25-hydroxyvitamin D concentrations are associated with insulin resistance and obesity in women with polycystic ovary syndrome. Exp. Clin. Endocrinol. Diabetes 2006, 114, 577–583. [Google Scholar] [CrossRef]
- Li, H.W.; Brereton, R.E.; Anderson, R.A.; Wallace, A.M.; Ho, C.K. Vitamin D deficiency is common and associated with metabolic risk factors in patients with polycystic ovary syndrome. Metabolism Clin. Exp. 2011, 60, 1475–1481. [Google Scholar] [CrossRef]
- Butler, A.E.; Dargham, S.R.; Abouseif, A.; El Shewehy, A.; Atkin, S.L. Vitamin D deficiency effects on cardiovascular parameters in women with polycystic ovary syndrome: A retrospective, cross-sectional study. J. Steroid Biochem. Mol. Biol. 2021, 211, 105892. [Google Scholar] [CrossRef]
- Lerchbaum, E.; Obermayer-Pietsch, B. Vitamin D and fertility: A systematic review. Eur. J. Endocrinol./Eur. Fed. Endocr. Soc. 2012, 166, 765–778. [Google Scholar] [CrossRef] [Green Version]
- Selimoglu, H.; Duran, C.; Kiyici, S.; Ersoy, C.; Guclu, M.; Ozkaya, G.; Tuncel, E.; Erturk, E.; Imamoglu, S. The effect of vitamin D replacement therapy on insulin resistance and androgen levels in women with polycystic ovary syndrome. J. Endocrinol. Investig. 2010, 33, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Krul-Poel, Y.H.M.; Snackey, C.; Louwers, Y.; Lips, P.T.A.M.; Lambalk, C.B.; Laven, J.S.E.; Simsek, S. The role of vitamin D in metabolic disturbances in polycystic ovary syndrome (PCOS): A systematic review. Eur. J. Endocrinol./Eur. Fed. Endocr. Soc. 2013, 169, 853–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wimalawansa, S.J. Vitamin D Deficiency: Effects on Oxidative Stress, Epigenetics, Gene Regulation, and Aging. Biology 2019, 8, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, T.K.; Allgar, V.; Dargham, S.R.; Kilpatrick, E.; Sathyapalan, T.; Maguiness, S.; Mokhtar Rudin, H.R.; Abdul Ghani, N.M.; Latiff, A.; Atkin, S.L. Association of Vitamin D Metabolites with Embryo Development and Fertilization in Women with and without PCOS Undergoing Subfertility Treatment. Front. Endocrinol. 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome (PCOS). Human Reprod. 2004, 19, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.R.; Kazantzidis, A.; Kift, R.C.; Farrar, M.D.; Wilkinson, J.; Rhodes, L.E. Meeting Vitamin D Requirements in White Caucasians at UK Latitudes: Providing a Choice. Nutrients 2018, 10, 497. [Google Scholar] [CrossRef] [Green Version]
- Kahal, H.; Halama, A.; Aburima, A.; Bhagwat, A.M.; Butler, A.E.; Graumann, J.; Suhre, K.; Sathyapalan, T.; Atkin, S.L. Effect of induced hypoglycemia on inflammation and oxidative stress in type 2 diabetes and control subjects. Sci. Rep. 2020, 10, 4750. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, S.; Vaught, J.D.; Bock, C.; Gold, L.; Katilius, E.; Keeney, T.R.; Kim, N.; Saccomano, N.A.; Wilcox, S.K.; Zichi, D.; et al. From SOMAmer-based biomarker discovery to diagnostic and clinical applications: A SOMAmer-based, streamlined multiplex proteomic assay. PLoS ONE 2011, 6, e26332. [Google Scholar] [CrossRef] [Green Version]
- Torun, A.N.; Vural, M.; Cece, H.; Camuzcuoglu, H.; Toy, H.; Aksoy, N. Paraoxonase-1 is not affected in polycystic ovary syndrome without metabolic syndrome and insulin resistance, but oxidative stress is altered. Gynecol. Endocrinol. 2011, 27, 988–992. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, X.; Yao, W.; Cui, X.; Li, N.; Lin, Z.; Zhao, B.; Miao, J. Esterase D stabilizes FKBP25 to suppress mTORC1. Cell. Mol. Biol. Lett. 2021, 26, 50. [Google Scholar] [CrossRef]
- Yao, W.; Yang, Y.; Chen, X.; Cui, X.; Zhou, B.; Zhao, B.; Lin, Z.; Miao, J. Activation of Esterase D by FPD5 Inhibits Growth of A549 Lung Cancer Cells via JAB1/p53 Pathway. Genes 2022, 13, 786. [Google Scholar] [CrossRef] [PubMed]
- Kuşçu, N.K.; Var, A. Oxidative stress but not endothelial dysfunction exists in non-obese, young group of patients with polycystic ovary syndrome. Acta Obstet. Gynecol. Scand. 2009, 88, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Uçkan, K.; Demir, H.; Başkiran, Y.; Demir, C. Investigation of activities enzyme prolidase (pro) and glutathione s-transferase (gst) in polycystic ovary syndrome (pcos) patients. J. Sci. Rep. A 2022, 050, 20–31. [Google Scholar]
- Fatima, Q.; Amin, S.; Kawa, I.A.; Jeelani, H.; Manzoor, S.; Rizvi, S.M.; Rashid, F. Evaluation of antioxidant defense markers in relation to hormonal and insulin parameters in women with polycystic ovary syndrome (PCOS): A case-control study. Diabetes Metab. Syndr. 2019, 13, 1957–1961. [Google Scholar] [CrossRef] [PubMed]
- Sabuncu, T.; Vural, H.; Harma, M.; Harma, M. Oxidative stress in polycystic ovary syndrome and its contribution to the risk of cardiovascular disease. Clin. Biochem. 2001, 34, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. Int. J. Mol. Sci. 2021, 22, 1786. [Google Scholar] [CrossRef]
- Yazıcı, D.; Sezer, H. Insulin Resistance, Obesity and Lipotoxicity. Adv. Exp. Med. Biol. 2017, 960, 277–304. [Google Scholar]
- Gobato, A.O.; Vasques, A.C.; Zambon, M.P.; Barros Filho Ade, A.; Hessel, G. Metabolic syndrome and insulin resistance in obese adolescents. Rev. Paul. Pediatr. 2014, 32, 55–62. [Google Scholar] [CrossRef]
- Moin, A.S.M.; Sathyapalan, T.; Diboun, I.; Elrayess, M.A.; Butler, A.E.; Atkin, S.L. Metabolic consequences of obesity on the hypercoagulable state of polycystic ovary syndrome. Sci. Rep. 2021, 11, 5320. [Google Scholar] [CrossRef]
- Butler, A.E.; Abouseif, A.; Dargham, S.R.; Sathyapalan, T.; Atkin, S.L. Metabolic comparison of polycystic ovarian syndrome and control women in Middle Eastern and UK Caucasian populations. Sci. Rep. 2020, 10, 18895. [Google Scholar] [CrossRef]
- Akbari, M.; Ostadmohammadi, V.; Lankarani, K.B.; Tabrizi, R.; Kolahdooz, F.; Heydari, S.T.; Kavari, S.H.; Mirhosseini, N.; Mafi, A.; Dastorani, M.; et al. The Effects of Vitamin D Supplementation on Biomarkers of Inflammation and Oxidative Stress among Women with Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Horm. Metab. Res. 2018, 50, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.S.; Ramin, J.; Rafison, B.; Windon, C.; Windon, A.; Liu, P.T. Redefining Human Vitamin D Sufficiency: Back to the Basics. Bone Res. 2013, 1, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.S.; Rafison, B.; Witzel, S.; Reyes, R.E.; Shieh, A.; Chun, R.; Zavala, K.; Hewison, M.; Liu, P.T. Regulation of the extrarenal CYP27B1-hydroxylase. J. Steroid Biochem. Mol. Biol. 2014, 144 Pt A, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Adams, I.P.; Dack, S.; Dickinson, F.M.; Ratledge, C. The distinctiveness of ATP:citrate lyase from Aspergillus nidulans. Biochim. Biophys. Acta 2002, 1597, 36–41. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Control (n = 24) | PCOS (n = 24) | |
|---|---|---|
| Age (years) | 32.5 ± 4.1 | 31 ± 6.4 |
| BMI (kg/m2) | 24.8 ± 1.1 | 25.9 ± 1.8 |
| Fasting glucose (mmol/L) | 4.9 ± 0.4 | 4.7 ± 0.8 |
| HbA1C (mmol/mol) | 30.9 ± 6.5 | 31.8 ± 3.0 |
| HOMA-IR | 1.8 ± 1.0 | 1.9 ± 1.6 |
| SHBG (nmol/L) | 104 ± 80 | 72 ± 62 |
| Free androgen index (FAI) | 1.3 ± 0.5 | 4.1 ± 2.9 ** |
| CRP (mg L−1) | 2.3 ± 2.3 | 2.7 ± 2.5 |
| AMH (ng/mL) | 24 ± 13 | 57 ± 14 ** |
| 25 hydroxy vitamin D3 (nmol/l) | 46.2± 23.5 | 54.0 ± 27.4 |
| 1,25 Dihydroxy vitamin D3 (ng/mL) | 0.03 ± 0.02 | 0.04 ± 0.2 |
| Control | PCOS | p-Value | ||
|---|---|---|---|---|
| Heat shock proteins | HSP90AA1/AB1 | 6566 (11,881) | 5160 (6109) | 0.57 |
| HSP70 (HSPA8) | 2140 (2361) | 1606 (348) | 0.23 | |
| HSP27 (HSPB1) | 2350 (1727) | 3773 (4633) | 0.13 | |
| MAPKAPK5 | 985 (2314) | 625 (504) | 0.42 | |
| DNAJB1 (HSP40) | 447 (296) | 485 (337) | 0.66 | |
| STIP1 | 7563 (7799) | 6785 (4837) | 0.65 | |
| STUB1 | 1660 (3163) | 1212 (1138) | 0.48 | |
| UBE2L3 | 1212 (635) | 1322 (764) | 0.55 | |
| UBE2N | 7696 (8053) | 7014 (6284) | 0.72 | |
| UBE2G2 | 4642 (1853) | 4401 (1002) | 0.54 | |
| Oxidative stress proteins | SOD1 | 973 (621) | 914 (325) | 0.64 |
| GSTP1 | 3358 (2260) | 3594 (763) | 0.65 | |
| GSTA3 | 209 (214) | 182 (119) | 0.53 | |
| ESD | 4606 (1887) | 6184 (2632) | 0.01 | |
| CAT | 22,445 (12,953) | 22,749 (8629) | 0.89 | |
| SOD2 | 48,222 (10,590) | 51,647 (12,763) | 0.26 | |
| SOD3 | 506 (455) | 625 (710) | 0.46 |
| 25(OH)D3 | 1,25(OH)2D3 | ||||
|---|---|---|---|---|---|
| r-Value | p-Value | r-Value | p-Value | ||
| Heat shock proteins | HSP90AA1/AB1 | 0.2 | 0.17 | 0.02 | 0.9 |
| HSP70 (HSPA8) | 0.06 | 0.67 | 0.02 | 0.93 | |
| HSP27 (HSPB1) | 0.22 | 0.14 | 0.09 | 0.59 | |
| MAPKAPK5 | 0.08 | 0.6 | 0.26 | 0.12 | |
| DNAJB1 (HSP40) | 0.24 | 0.1 | 0.0005 | 0.98 | |
| STIP1 | 0.06 | 0.69 | 0.03 | 0.87 | |
| STUB1 | 0.05 | 0.73 | 0.06 | 0.73 | |
| UBE2L3 | 0.0002 | 0.96 | 0.03 | 0.91 | |
| UBE2N | 0.09 | 0.53 | 0.03 | 0.85 | |
| UBE2G2 | 0.06 | 0.71 | 0.16 | 0.34 | |
| Oxidative stress proteins | SOD1 | 0.05 | 0.74 | 0.09 | 0.61 |
| GSTP1 | 0.11 | 0.44 | 0.11 | 0.52 | |
| GSTA3 | 0.12 | 0.43 | 0.11 | 0.53 | |
| ESD | 0.06 | 0.69 | 0.14 | 0.39 | |
| CAT | 0.25 | 0.09 | 0.0003 | 0.99 | |
| SOD2 | 0.11 | 0.48 | 0.08 | 0.66 | |
| SOD3 | 0.17 | 0.25 | 0.21 | 0.22 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nandakumar, M.; Sathyapalan, T.; Butler, A.E.; Atkin, S.L. Oxidative Stress Markers and Heat Shock Proteins in Non-Obese Women with Polycystic Ovary Syndrome Are Not Elevated and Show No Correlation with Vitamin D. Biomedicines 2023, 11, 2044. https://doi.org/10.3390/biomedicines11072044
Nandakumar M, Sathyapalan T, Butler AE, Atkin SL. Oxidative Stress Markers and Heat Shock Proteins in Non-Obese Women with Polycystic Ovary Syndrome Are Not Elevated and Show No Correlation with Vitamin D. Biomedicines. 2023; 11(7):2044. https://doi.org/10.3390/biomedicines11072044
Chicago/Turabian StyleNandakumar, Manjula, Thozhukat Sathyapalan, Alexandra E. Butler, and Stephen L. Atkin. 2023. "Oxidative Stress Markers and Heat Shock Proteins in Non-Obese Women with Polycystic Ovary Syndrome Are Not Elevated and Show No Correlation with Vitamin D" Biomedicines 11, no. 7: 2044. https://doi.org/10.3390/biomedicines11072044