

Expression of CCR8 and CCX-CKR on Basophils in Chronic Urticaria Is Amplified by IgE-Mediated Activation
Abstract
:1. Introduction
2. Methods
2.1. Study Design
2.2. Flow Cytometric Analysis of Basophils
2.3. Statistical Analysis
3. Results
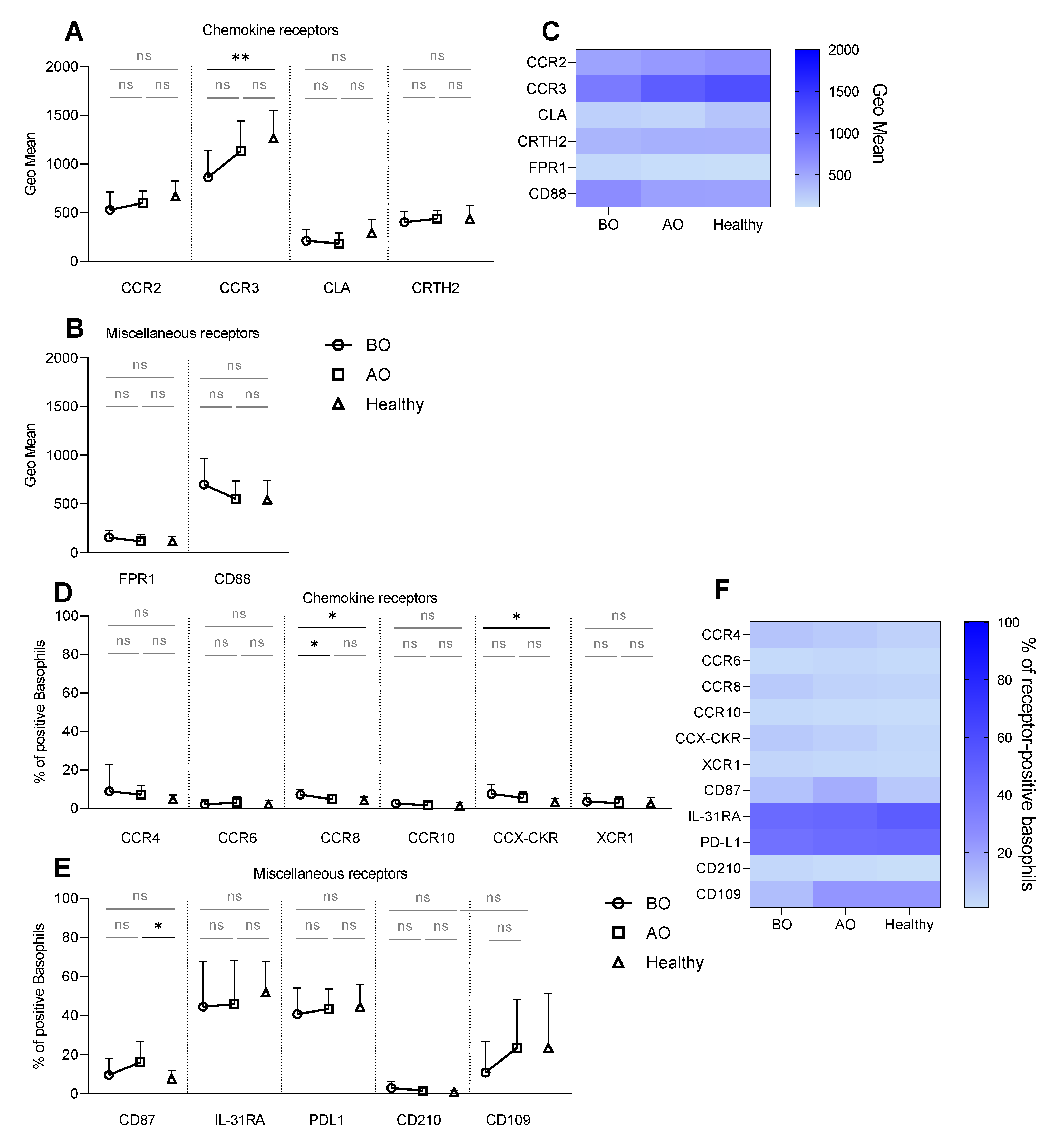
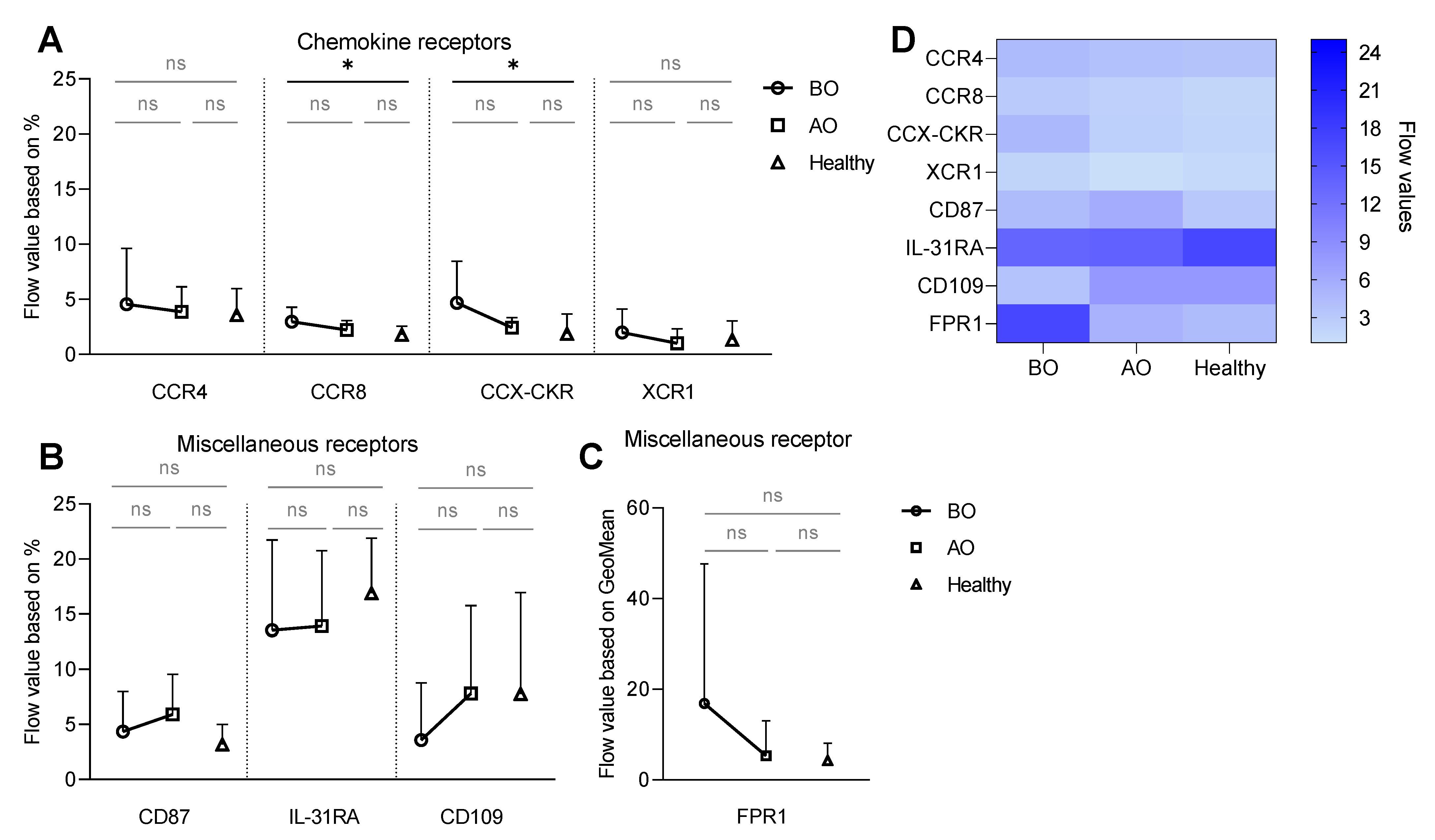
3.1. Receptor Expression on Resting Basophils—Increased Expression of CCR8 and CCX-CKR and Decreased Expression of CCR3 on Unstimulated (Resting) Basophils in CU
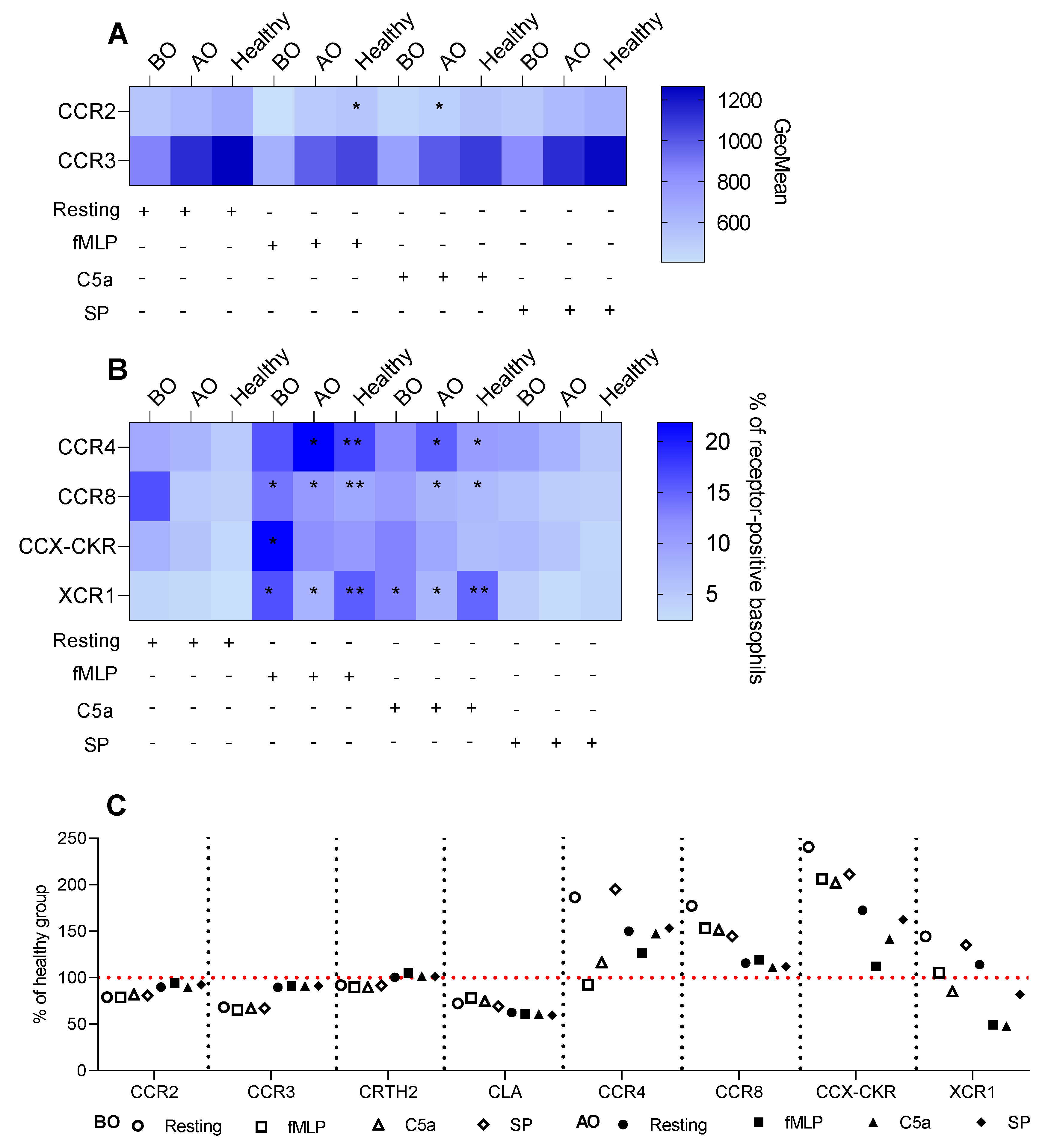
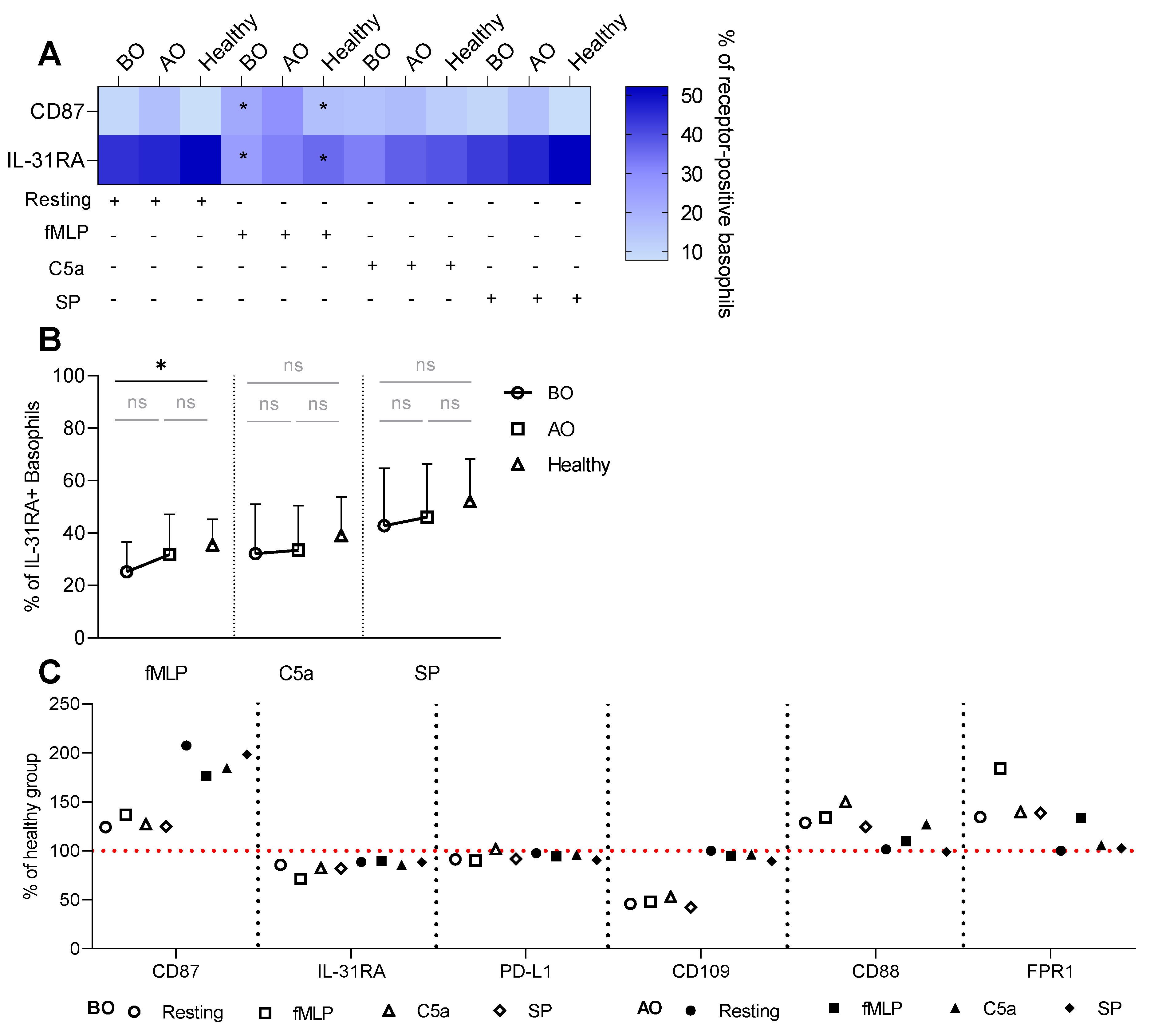
3.2. IgE-Mediated Stimulation Elevates the Expression of CCR8 and CCX-CKR, Boosting the Chemokine Response Potential of Basophils in CU
3.3. Non-IgE-Mediated Stimulation Mimics the Effect of the IgE-FcεRI Axis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Saini, S.S.; Omachi, T.A.; Trzaskoma, B.; Hulter, H.N.; Rosén, K.; Sterba, P.M.; Courneya, J.-P.; Lackey, A.; Chen, H. Effect of Omalizumab on Blood Basophil Counts in Patients with Chronic Idiopathic/Spontaneous Urticaria. J. Investig. Dermatol. 2017, 137, 958–961. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Satoh, T.; Takayama, K.; Miyagishi, C.; Walls, A.; Yokozeki, H. Basophil recruitment and activation in inflammatory skin diseases. Allergy 2011, 66, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Deza, G.; Bertolín-Colilla, M.; Pujol, R.; Curto-Barredo, L.; Soto, D.; García, M.; Hernández, P.; Gimeno, R.; Giménez-Arnau, A. Basophil FcεRI Expression in Chronic Spontaneous Urticaria: A Potential Immunological Predictor of Response to Omalizumab Therapy. Acta Derm.-Venereol. 2017, 97, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wang, J.; Zhu, W.; Xu, C.; He, S. Upregulated expression of substance P in basophils of the patients with chronic spontaneous urticaria: Induction of histamine release and basophil accumulation by substance P. Cell Biol. Toxicol. 2016, 32, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Vasagar, K.; Vonakis, B.M.; Gober, L.M.; Viksman, A.; Gibbons, S.P.; Saini, S.S. Evidence of in vivo basophil activation in chronic idiopathic urticaria. Clin. Exp. Allergy 2006, 36, 770–776. [Google Scholar] [CrossRef]
- Oliver, E.; Sterba, P.M.; Saini, S.S.; Vonakis, B.M.; Devine, K. The Expression of CRTh2 on Blood Basophils and Eosinophils in Chronic Idiopathic Urticaria. J. Allergy Clin. Immunol. 2015, 135, AB126. [Google Scholar] [CrossRef]
- Iikura, M.; Miyamasu, M.; Yamaguchi, M.; Kawasaki, H.; Matsushima, K. Chemokine receptors in human basophils: Inducible expression of functional CXCR4. J. Leukoc. Biol. 2001, 70, 113–120. [Google Scholar] [CrossRef]
- Soler, D.; Humphreys, T.L.; Spinola, S.M.; Campbell, J.J. CCR4 versus CCR10 in human cutaneous TH lymphocyte trafficking. Blood 2003, 101, 1677–1682. [Google Scholar] [CrossRef]
- Furue, K.; Ito, T.; Tsuji, G.; Nakahara, T.; Furue, M. The CCL20 and CCR6 axis in psoriasis. Scand. J. Immunol. 2020, 91, e12846. [Google Scholar] [CrossRef]
- Gombert, M.; Dieu-Nosjean, M.-C.; Winterberg, F.; Bünemann, E.; Kubitza, R.C.; Da Cunha, L.; Haahtela, A.; Lehtimäki, S.; Muüller, A.; Rieker, J.; et al. CCL1-CCR8 Interactions: An Axis Mediating the Recruitment of T Cells and Langerhans-Type Dendritic Cells to Sites of Atopic Skin Inflammation. J. Immunol. 2005, 174, 5082–5091. [Google Scholar] [CrossRef]
- Hudak, S.; Hagen, M.; Liu, Y.; Catron, D.; Oldham, E.; McEvoy, L.M.; Bowman, E.P. Immune Surveillance and Effector Functions of CCR10+ Skin Homing T Cells. J. Immunol. 2002, 169, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Comerford, I.; Milasta, S.; Morrow, V.; Milligan, G.; Nibbs, R. The chemokine receptor CCX-CKR mediates effective scavenging of CCL19 In Vitro. Eur. J. Immunol. 2006, 36, 1904–1916. [Google Scholar] [CrossRef] [PubMed]
- Blom, L.H.; Bartko, E.A.; Møller, T.K.R.; Poulsen, L.K.; Jensen, B.M. FcεRI-activated basophils express CCR4, CCR8, CCR9, CCX-CKR and XCR1. Allergy 2022, 78, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Luquin, E.; Kaplan, A.P.; Ferrer, M. Increased responsiveness of basophils of patients with chronic urticaria to sera but hypo-responsiveness to other stimuli. Clin. Exp. Allergy 2005, 35, 456–460. [Google Scholar] [CrossRef]
- Rauber, M.M.; Pickert, J.; Holiangu, L.; Möbs, C.; Pfützner, W. Functional and phenotypic analysis of basophils allows determining distinct subtypes in patients with chronic urticaria. Allergy Eur. J. Allergy Clin. Immunol. 2017, 72, 1904–1911. [Google Scholar] [CrossRef]
- Raap, U.; Gehring, M.; Kleiner, S.; Rüdrich, U.; Eiz-Vesper, B.; Haas, H.; Kapp, A.; Gibbs, B.F. Human basophils are a source of—And are differentially activated by—IL-31. Clin. Exp. Allergy 2017, 47, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Zhou, Q.; Liu, C.; Ying, M.; Xu, S. Increased plasma IL-17, IL-31, and IL-33 levels in chronic spontaneous urticaria. Sci. Rep. 2017, 7, 17797. [Google Scholar] [CrossRef]
- Tsai, Y.-L.; Ha, D.P.; Zhao, H.; Carlos, A.J.; Wei, S.; Pun, T.K.; Wu, K.; Zandi, E.; Kelly, K.; Lee, A.S. Endoplasmic reticulum stress activates SRC, relocating chaperones to the cell surface where GRP78/CD109 blocks TGF-β signaling. Proc. Natl. Acad. Sci. USA 2018, 115, E4245–E4254. [Google Scholar] [CrossRef]
- Béné, M.C.; Castoldi, G.; Knapp, W.; Rigolin, G.M.; Escribano, L.; Lemez, P.; Ludwig, W.-D.; Matutes, E.; Orfao, A.; Lanza, F.; et al. CD87 (urokinase-type plasminogen activator receptor), function and pathology in hematological disorders: A review. Leukemia 2004, 18, 394–400. [Google Scholar] [CrossRef]
- Bartko, E.A.; Elberling, J.; Blom, L.H.; Poulsen, L.K.; Jensen, B.M. Elevated, FcεRI-dependent MRGPRX2 expression on basophils in chronic urticaria. Ski. Health Dis. 2022, e195. [Google Scholar] [CrossRef]
- Pan, Q.; Feng, Y.; Peng, Y.; Zhou, H.; Deng, Z.; Li, L.; Han, H.; Lin, J.; Shi, L.; Wang, S.; et al. Basophil Recruitment to Skin Lesions of Patients with Systemic Lupus Erythematosus Mediated by CCR1 and CCR2. Cell. Physiol. Biochem. 2017, 43, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Korosec, P.; Turner, P.; Silar, M.; Kopac, P.; Kosnik, M.; Gibbs, B.F.; Shamji, M.H.; Custovic, A.; Rijavec, M. Basophils, high-affinity IgE receptors, and CCL2 in human anaphylaxis. J. Allergy Clin. Immunol. 2017, 140, 750–758.e15. [Google Scholar] [CrossRef]
- Johal, K.J.; Chichester, K.L.; Oliver, E.T.; Devine, K.C.; Bieneman, A.P.; Schroeder, J.T.; MacGlashan, D.W.; Saini, S.S. The efficacy of omalizumab treatment in chronic spontaneous urticaria is associated with basophil phenotypes. J. Allergy Clin. Immunol. 2021, 147, 2271–2280.e8. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Luquin, E.; Sanchez-Ibarrola, A.; Moreno, C.; Sanz, M.L.; Kaplan, A.P. Secretion of Cytokines, Histamine and Leukotrienes in Chronic Urticaria. Int. Arch. Allergy Immunol. 2002, 129, 254–260. [Google Scholar] [CrossRef]
- Uguccioni, M.; Mackay, C.R.; Ochensberger, B.; Loetscher, P.; Rhis, S.; LaRosa, G.J.; Rao, P.; Ponath, P.D.; Baggiolini, M.; Dahinden, C.A. High expression of the chemokine receptor CCR3 in human blood basophils. Role in activation by eotaxin, MCP-4, and other chemokines. J. Clin. Investig. 1997, 100, 1137–1143. [Google Scholar] [CrossRef]
- Vestergaard, C.; Deleuran, M.; Gesser, B.; Larsen, C.G. Expression of the T-helper 2-specific chemokine receptor CCR4 on CCR10-positive lymphocytes in atopic dermatitis skin but not in psoriasis skin. Br. J. Dermatol. 2003, 149, 457–463. [Google Scholar] [CrossRef]
- Homey, B.; Alenius, H.; Müller, A.; Soto, H.; Bowman, E.P.; Yuan, W.; McEvoy, L.; Lauerma, A.I.; Assmann, T.; Bünemann, E.; et al. CCL27–CCR10 interactions regulate T cell–mediated skin inflammation. Nat. Med. 2002, 8, 157. [Google Scholar] [CrossRef]
- Velazquez, J.R.; Teran, L.M. Chemokines and Their Receptors in the Allergic Airway Inflammatory Process. Clin. Rev. Allergy Immunol. 2011, 41, 76–88. [Google Scholar] [CrossRef]
- Borroni, E.M.; Bonecchi, R.; Buracchi, C.; Savino, B.; Mantovani, A.; Locati, M. Chemokine Decoy Receptors: New Players in Reproductive Immunology. Immunol. Investig. 2008, 37, 483–497. [Google Scholar] [CrossRef]
- Comerford, I.; Litchfield, W.; Harata-Lee, Y.; Nibbs, R.J.; McColl, S.R. Regulation of chemotactic networks by ‘atypical’ receptors. Bioessays 2007, 29, 237–247. [Google Scholar] [CrossRef]
- Song, G.; Feng, T.; Zhao, R.; Lu, Q.; Diao, Y.; Guo, Q.; Wang, Z.; Zhang, Y.; Ge, L.; Pan, J.; et al. CD109 regulates the inflammatory response and is required for the pathogenesis of rheumatoid arthritis. Ann. Rheum. Dis. 2019, 78, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Blom, L.H.; Martel, B.C.; Larsen, L.F.; Hansen, C.V.; Christensen, M.P.; Juel-Berg, N.; Litman, T.; Poulsen, L.K. The immunoglobulin superfamily member CD200R identifies cells involved in type 2 immune responses. Allergy Eur. J. Allergy Clin. Immunol. 2017, 72, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Raap, U.; Wieczorek, D.; Gehring, M.; Pauls, I.; Ständer, S.; Kapp, A.; Wedi, B. Increased levels of serum IL-31 in chronic spontaneous urticaria. Exp. Dermatol. 2010, 19, 464–466. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.E.; Nijmeh, H.S.; Brightling, C.E.; Sayers, I. uPAR regulates bronchial epithelial repair in vitro and is elevated in asthmatic epithelium. Thorax 2012, 67, 477–487. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % | Geo Mean | |
|---|---|---|
| Chemokine receptors | CCR4 | CCR2 |
| CCR6 | CCR3 | |
| CCR8 | CLA | |
| CCR10 | CRTH2 | |
| CCX-CKR | ||
| XCR1 | ||
| Miscellaneous receptors | CD87 | FPR1 |
| IL-31RA | CD88 | |
| PD-L1 | ||
| CD210 | ||
| CD109 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartko, E.A.; Blom, L.H.; Elberling, J.; Poulsen, L.K.; Jensen, B.M. Expression of CCR8 and CCX-CKR on Basophils in Chronic Urticaria Is Amplified by IgE-Mediated Activation. Biomedicines 2023, 11, 1537. https://doi.org/10.3390/biomedicines11061537
Bartko EA, Blom LH, Elberling J, Poulsen LK, Jensen BM. Expression of CCR8 and CCX-CKR on Basophils in Chronic Urticaria Is Amplified by IgE-Mediated Activation. Biomedicines. 2023; 11(6):1537. https://doi.org/10.3390/biomedicines11061537
Chicago/Turabian StyleBartko, Ewa A., Lars H. Blom, Jesper Elberling, Lars K. Poulsen, and Bettina M. Jensen. 2023. "Expression of CCR8 and CCX-CKR on Basophils in Chronic Urticaria Is Amplified by IgE-Mediated Activation" Biomedicines 11, no. 6: 1537. https://doi.org/10.3390/biomedicines11061537