
Microvascular Fragments Protect Ischemic Musculocutaneous Flap Tissue from Necrosis by Improving Nutritive Tissue Perfusion and Suppressing Apoptosis
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
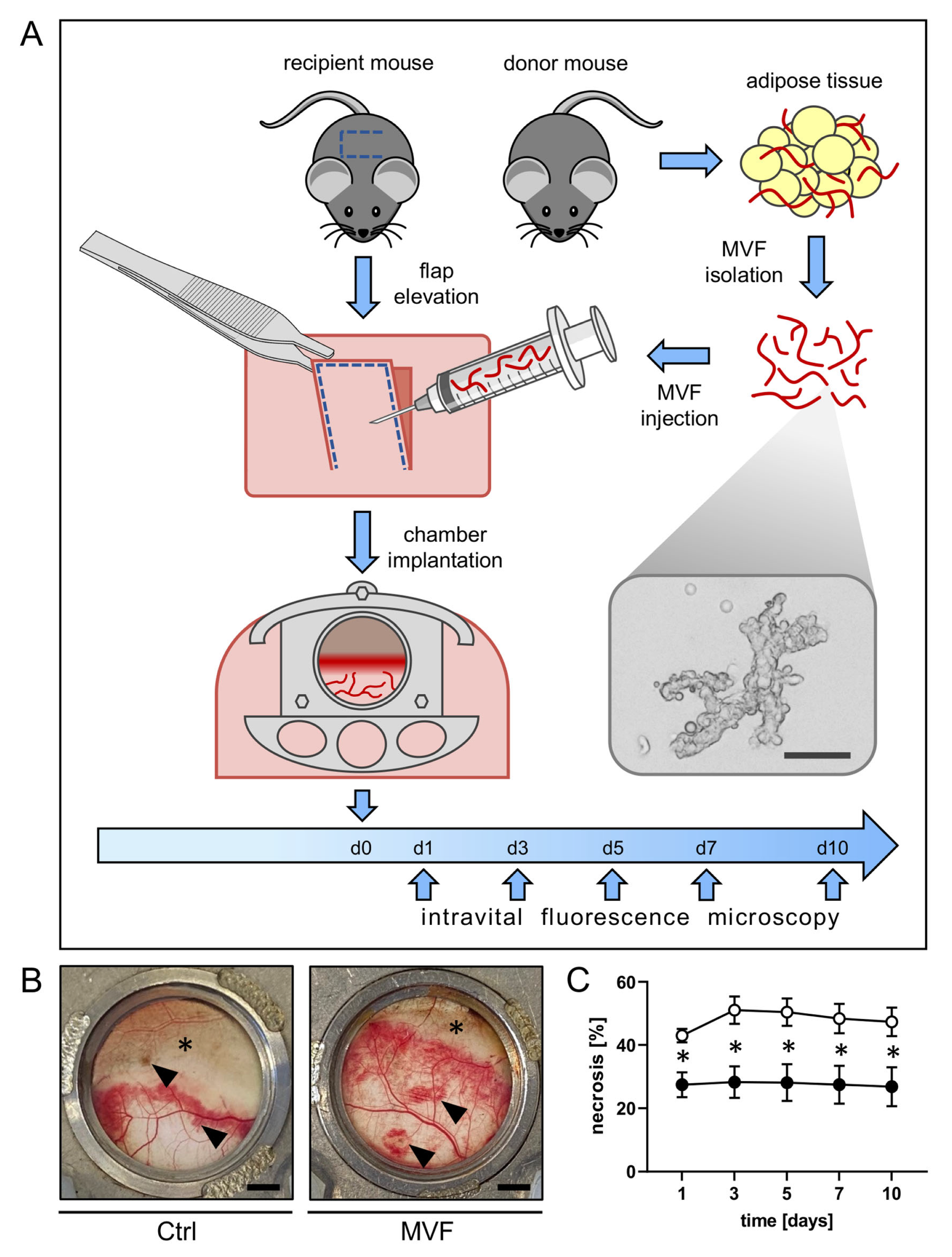
2.1. Animals
2.2. MVF Isolation
2.3. Anesthesia
2.4. Dorsal Skinfold Chamber
2.5. Intravital Fluorescence Microscopy
2.6. Histology and Immunohistochemistry
2.7. Statistical Analysis
3. Results
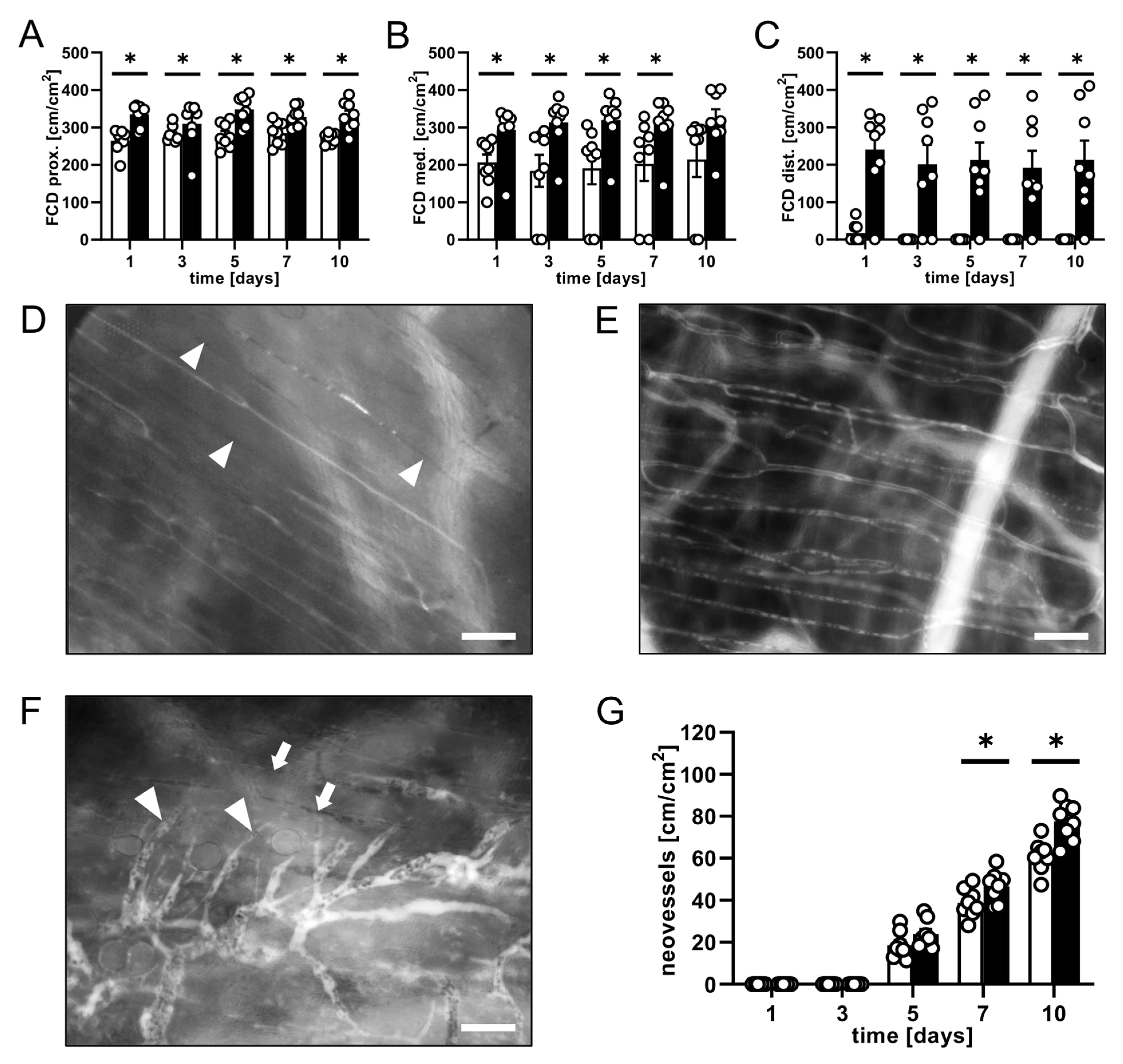
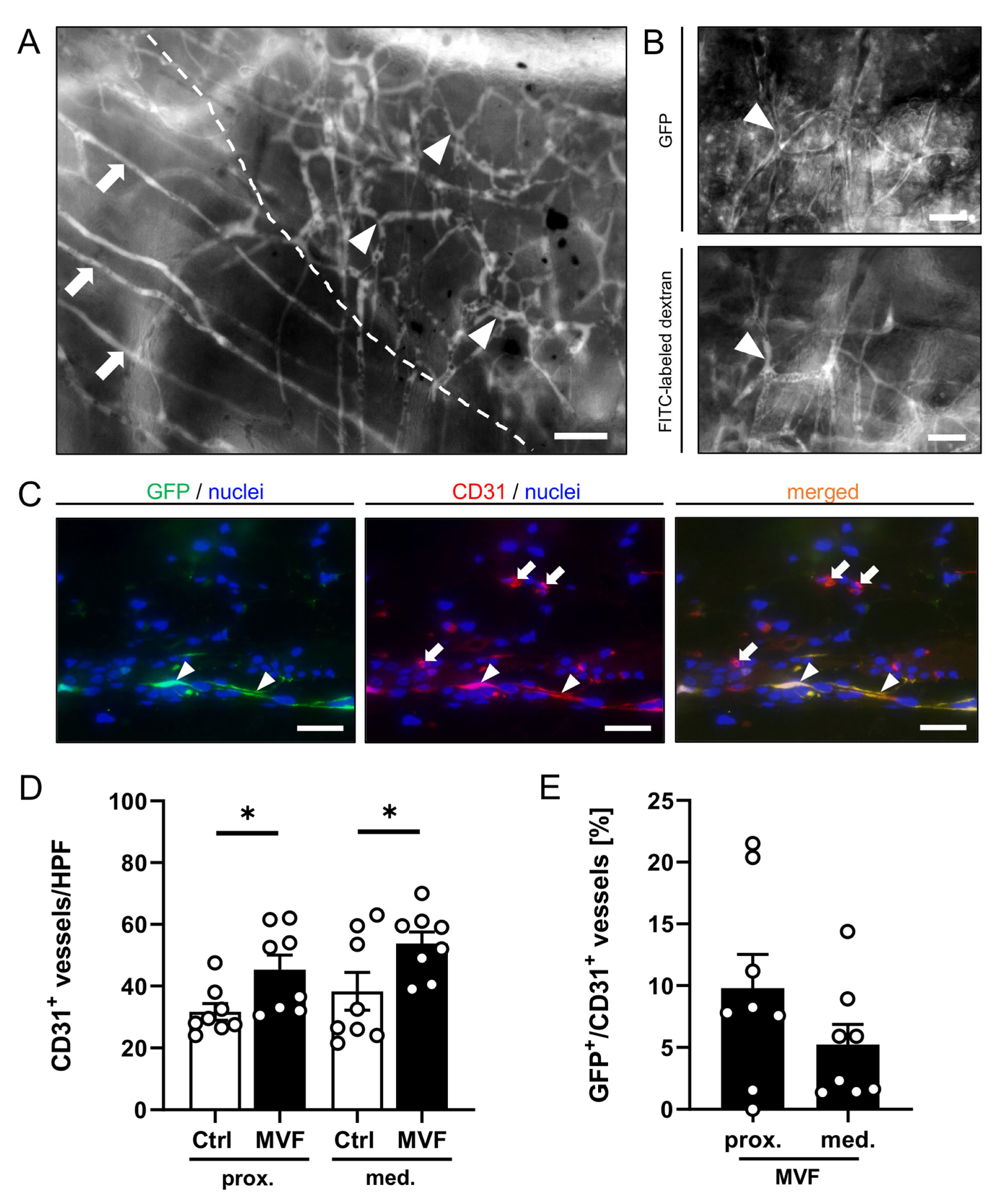
3.1. Intravital Fluorescence Microscopy
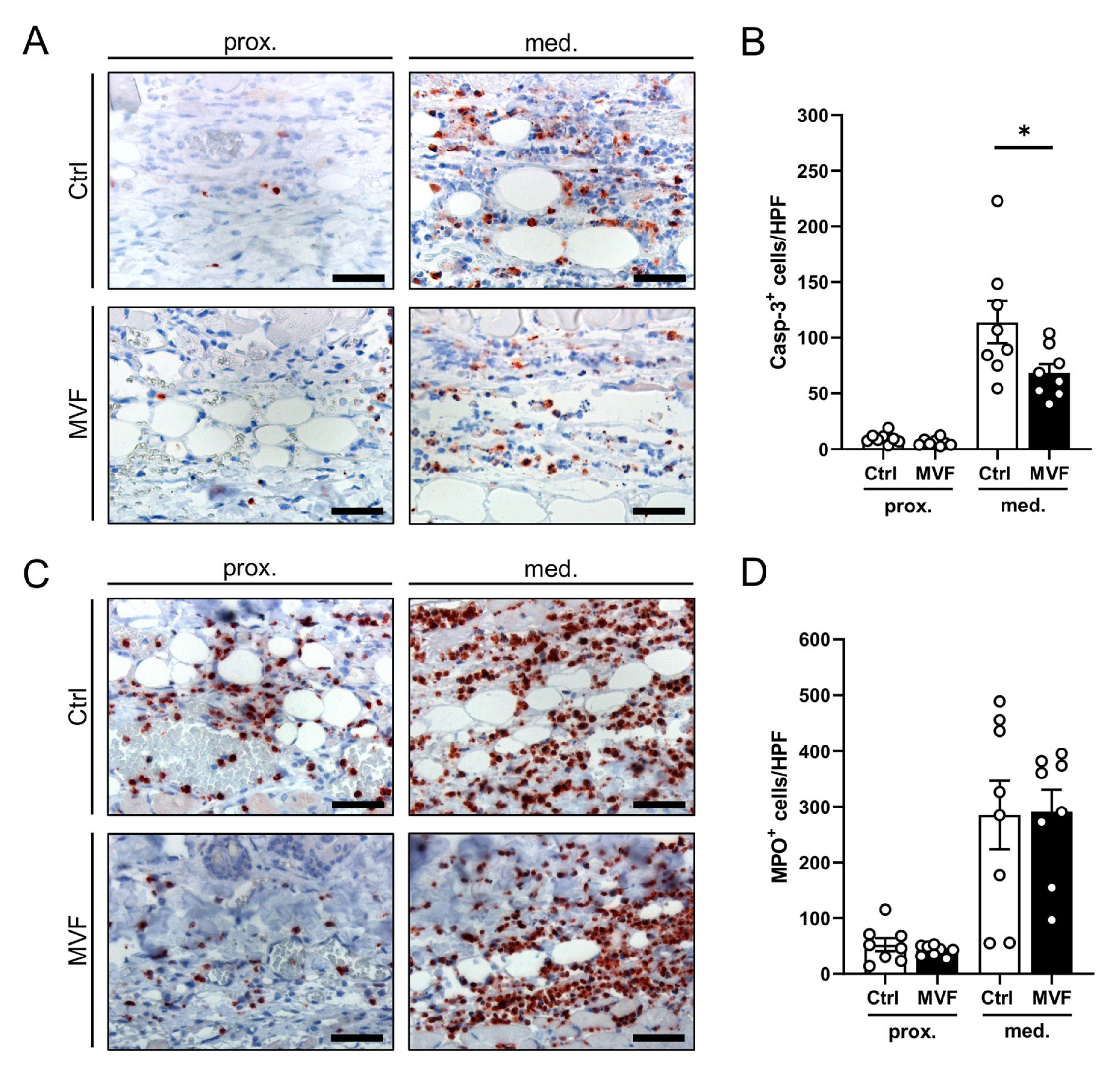
3.2. Histological and Immunohistochemical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kalogeris, T.; Bao, Y.; Korthuis, R.J. Mitochondrial reactive oxygen species: A double edged sword in ischemia/reperfusion vs preconditioning. Redox Biol. 2014, 2, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.B. The Physiology and Biomechanics of Skin Flaps. Facial Plast. Surg. Clin. N. Am. 2017, 25, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Z.; Baynosa, R.C.; Zamboni, W.A. Update on Ischemia-Reperfusion Injury for the Plastic Surgeon. Plast. Reconstr. Surg. 2011, 128, 685e–692e. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.; Rathbone, C.R. Microvascular Fragment Transplantation Improves Rat Dorsal Skin Flap Survival. Plast. Reconstr. Surg. Glob. Open 2016, 4, e1140. [Google Scholar] [CrossRef]
- Nakano, M.; Nakajima, Y.; Kudo, S.; Tsuchida, Y.; Nakamura, H.; Fukuda, O. Effect of Autotransplantation of Microvessel Fragments on Experimental Random-Pattern Flaps in the Rat. Eur. Surg. Res. 1998, 30, 149–160. [Google Scholar] [CrossRef]
- Frueh, F.S.; Später, T.; Scheuer, C.; Menger, M.D.; Laschke, M.W. Isolation of Murine Adipose Tissue-derived Microvascular Fragments as Vascularization Units for Tissue Engineering. J. Vis. Exp. 2017, 122, e55721. [Google Scholar] [CrossRef]
- Laschke, M.W.; Menger, M.D. Adipose Tissue-Derived Microvascular Fragments: Natural Vascularization Units for Regenerative Medicine. Trends Biotechnol. 2015, 33, 442–448. [Google Scholar] [CrossRef]
- McDaniel, J.S.; Pilia, M.; Ward, C.L.; Pollot, B.E.; Rathbone, C.R. Characterization and multilineage potential of cells derived from isolated microvascular fragments. J. Surg. Res. 2014, 192, 214–222. [Google Scholar] [CrossRef]
- Laschke, M.; Kleer, S.; Scheuer, C.; Schuler, S.; Garcia, P.; Eglin, D.; Alini, M.; Menger, M.D. Vascularisation of porous scaffolds is improved by incorporation of adipose tissue-derived microvascular fragments. Eur. Cells Mater. 2012, 24, 266–277. [Google Scholar] [CrossRef]
- Pilia, M.; McDaniel, J.S.; Guda, T.; Chen, X.K.; Rhoads, R.P.; Allen, R.E.; Corona, B.T.; Rathbone, C.R. Transplantation and perfusion of microvascular fragments in a rodent model of volumetric muscle loss injury. Eur. Cells Mater. 2014, 28, 11–24. [Google Scholar] [CrossRef]
- Shepherd, B.R.; Chen, H.Y.; Smith, C.M.; Gruionu, G.; Williams, S.K.; Hoying, J.B. Rapid Perfusion and Network Remodeling in a Microvascular Construct after Implantation. Arter. Thromb. Vasc. Biol. 2004, 24, 898–904. [Google Scholar] [CrossRef]
- Später, T.; Frueh, F.; Menger, M.D.; Laschke, M.W. Potentials and limitations of Integra® flowable wound matrix seeded with adipose tissue-derived microvascular fragments. Eur. Cells Mater. 2017, 33, 268–278. [Google Scholar] [CrossRef]
- Später, T.; Frueh, F.S.; Nickels, R.M.; Menger, M.D.; Laschke, M.W. Prevascularization of collagen-glycosaminoglycan scaffolds: Stromal vascular fraction versus adipose tissue-derived microvascular fragments. J. Biol. Eng. 2018, 12, 24. [Google Scholar] [CrossRef]
- Shepherd, B.R.; Hoying, J.B.; Williams, S.K. Microvascular Transplantation after Acute Myocardial Infarction. Tissue Eng. 2007, 13, 2871–2879. [Google Scholar] [CrossRef]
- Laschke, M.W.; Menger, M.D. Microvascular Fragments in Microcirculation Research and Regenerative Medicine. Tissue Eng. Part B Rev. 2022, 28, 1109–1120. [Google Scholar] [CrossRef]
- Frueh, F.S.; Später, T.; Lindenblatt, N.; Calcagni, M.; Giovanoli, P.; Scheuer, C.; Menger, M.D.; Laschke, M.W. Adipose Tissue-Derived Microvascular Fragments Improve Vascularization, Lymphangiogenesis, and Integration of Dermal Skin Substitutes. J. Investig. Dermatol. 2017, 137, 217–227. [Google Scholar] [CrossRef]
- Harder, Y.; Amon, M.; Erni, D.; Menger, M. Evolution of ischemic tissue injury in a random pattern flap: A new mouse model using intravital microscopy. J. Surg. Res. 2004, 121, 197–205. [Google Scholar] [CrossRef]
- Weinzierl, A.; Harder, Y.; Schmauss, D.; Menger, M.D.; Laschke, M.W. Boosting Tissue Vascularization: Nanofat as a Potential Source of Functional Microvessel Segments. Front. Bioeng. Biotechnol. 2022, 10, 820835. [Google Scholar] [CrossRef]
- Harder, Y.; Schmauss, D.; Wettstein, R.; Egaña, J.T.; Weiss, F.; Weinzierl, A.; Schuldt, A.; Machens, H.-G.; Menger, M.D.; Rezaeian, F. Ischemic Tissue Injury in the Dorsal Skinfold Chamber of the Mouse: A Skin Flap Model to Investigate Acute Persistent Ischemia. J. Vis. Exp. 2014, 93, e51900. [Google Scholar] [CrossRef]
- De Vriese, A.S.; Verbeuren, T.J.; Vallez, M.-O.; Lameire, N.H.; De Buyzere, M.; Vanhoutte, P.M. Off-Line Analysis of Red Blood Cell Velocity in Renal Arterioles. J. Vasc. Res. 2000, 37, 26–31. [Google Scholar] [CrossRef]
- Baker, M.; Wayland, H. On-line volume flow rate and velocity profile measurement for blood in microvessels. Microvasc. Res. 1974, 7, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Menger, M.D.; Lehr, H.-A. Scope and perspectives of intravital microscopy—Bridge over from in vitro to in vivo. Immunol. Today 1993, 14, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Porras, M.A.G.; Stojkova, K.; Acosta, F.M.; Rathbone, C.R.; Brey, E.M. Engineering Human Beige Adipose Tissue. Front. Bioeng. Biotechnol. 2022, 10, 906395. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liang, C.; Gao, X.; Huang, H.; Xing, X.; Tang, Q.; Yang, J.; Wu, Y.; Li, M.; Li, H.; et al. Adipose Tissue–derived Microvascular Fragments as Vascularization Units for Dental Pulp Regeneration. J. Endod. 2021, 47, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Laschke, M.W.; Später, T.; Menger, M.D. Microvascular Fragments: More Than Just Natural Vascularization Units. Trends Biotechnol. 2021, 39, 24–33. [Google Scholar] [CrossRef]
- Uysal, A.C.; Mizuno, H.; Tobita, M.; Ogawa, R.; Hyakusoku, H. The Effect of Adipose-Derived Stem Cells on Ischemia-Reperfusion Injury: Immunohistochemical and Ultrastructural Evaluation. Plast. Reconstr. Surg. 2009, 124, 804–815. [Google Scholar] [CrossRef]
- Hollenbeck, S.T.; Senghaas, A.; Komatsu, I.; Zhang, Y.; Erdmann, D.; Klitzman, B. Tissue engraftment of hypoxic-preconditioned adipose-derived stem cells improves flap viability. Wound Repair Regen. 2012, 20, 872–878. [Google Scholar] [CrossRef]
- Lee, D.W.; Jeon, Y.R.; Cho, E.J.; Kang, J.H.; Lew, D.H. Optimal administration routes for adipose-derived stem cells therapy in ischaemic flaps. J. Tissue Eng. Regen. Med. 2014, 8, 596–603. [Google Scholar] [CrossRef]
- Laschke, M.W.; Karschnia, P.; Scheuer, C.; Heß, A.; Metzger, W.; Menger, M.D. Effects of cryopreservation on adipose tissue-derived microvascular fragments. J. Tissue Eng. Regen. Med. 2018, 12, 1020–1030. [Google Scholar] [CrossRef]
- Kryger, Z.; Zhang, F.; Dogan, T.; Cheng, C.; Lineaweaver, W.C.; Buncke, H.J. The effects of VEGF on survival of a random flap in the rat: Examination of various routes of administration. Br. J. Plast. Surg. 2000, 53, 234–239. [Google Scholar] [CrossRef]
- Zhang, F.; Oswald, T.; Lin, S.; Cai, Z.; Lei, M.; Jones, M.; Angel, M.; Lineaweaver, W. Vascular endothelial growth factor (VEGF) expression and the effect of exogenous VEGF on survival of a random flap in the rat. Br. J. Plast. Surg. 2003, 56, 653–659. [Google Scholar] [CrossRef]
- Vourtsis, S.A.; Spyriounis, P.K.; Agrogiannis, G.D.; Ionac, M.; Papalois, A.E. VEGF Application on Rat Skin Flap Survival. J. Investig. Surg. 2012, 25, 14–19. [Google Scholar] [CrossRef]
- Vourtsis, S.A.; Papalois, A.E.; Agrogiannis, G.D.; Spyriounis, P.K.; Patsouris, E.; Ionac, M. Improvement of a long random skin flap survival by application of vascular endothelial growth factor in various ways of local administration in a rat model. Indian J. Plast. Surg. 2012, 45, 102–108. [Google Scholar] [CrossRef]
- He, H.; Venema, V.J.; Gu, X.; Venema, R.C.; Marrero, M.B.; Caldwell, R.B. Vascular Endothelial Growth Factor Signals Endothelial Cell Production of Nitric Oxide and Prostacyclin through Flk-1/KDR Activation of c-Src. J. Biol. Chem. 1999, 274, 25130–25135. [Google Scholar] [CrossRef]
- Weinzierl, A.; Harder, Y.; Schmauss, D.; Menger, M.D.; Laschke, M.W. Bromelain Protects Critically Perfused Musculocutaneous Flap Tissue from Necrosis. Biomedicines 2022, 10, 1449. [Google Scholar] [CrossRef]
- Huang, C.; Qing, L.; Pang, X.; Fu, J.; Xiao, Y.; Tang, J.; Wu, P. Melatonin Improved the Survival of Multi-Territory Perforator Flaps by Promoting Angiogenesis and Inhibiting Apoptosis via the NRF2/FUNDC1 Axis. Front. Pharmacol. 2022, 13, 921189. [Google Scholar] [CrossRef]
- Xin, D.; Quan, R.; Zeng, L.; Xu, C.; Tang, Y. Lipoxin A4 protects rat skin flaps against ischemia-reperfusion injury through inhibiting cell apoptosis and inflammatory response induced by endoplasmic reticulum stress. Ann. Transl. Med. 2020, 8, 1086. [Google Scholar] [CrossRef]
- Später, T.; Menger, M.M.; Nickels, R.M.; Menger, M.D.; Laschke, M.W. Macrophages promote network formation and maturation of transplanted adipose tissue-derived microvascular fragments. J. Tissue Eng. 2020, 11, 2041731420911816. [Google Scholar] [CrossRef]
- Qian, Q.; He, W.; Liu, D.; Yin, J.; Ye, L.; Chen, P.; Xu, D.; Liu, J.; Li, Y.; Zeng, G.; et al. M2a macrophage can rescue proliferation and gene expression of benign prostate hyperplasia epithelial and stroma cells from insulin-like growth factor 1 knockdown. Prostate 2021, 81, 530–542. [Google Scholar] [CrossRef]
- Gehmert, S.; Wenzel, C.; Loibl, M.; Brockhoff, G.; Huber, M.; Krutsch, W.; Nerlich, M.; Gosau, M.; Klein, S.; Schreml, S.; et al. Adipose Tissue-Derived Stem Cell Secreted IGF-1 Protects Myoblasts from the Negative Effect of Myostatin. BioMed Res. Int. 2014, 2014, 129048. [Google Scholar] [CrossRef]
- Lee, S.H. Human Adipose-Derived Stem Cells’ Paracrine Factors in Conditioned Medium Can Enhance Porcine Oocyte Maturation and Subsequent Embryo Development. Int. J. Mol. Sci. 2021, 22, 579. [Google Scholar] [CrossRef] [PubMed]
- Assmann, A.; Heke, M.; Kröpil, P.; Ptok, L.; Hafner, D.; Ohmann, C.; Martens, A.; Karluß, A.; Emmert, M.Y.; Kutschka, I.; et al. Laser-Supported CD133+ Cell Therapy in Patients with Ischemic Cardiomyopathy: Initial Results from a Prospective Phase I Multicenter Trial. PLoS ONE 2014, 9, e101449. [Google Scholar] [CrossRef] [PubMed]
- Trevisson-Redondo, B.; Becerro-De-Bengoa-Vallejo, R.; Sevillano, D.; González, N.; Losa-Iglesias, M.E.; López-López, D.; Alou, L. Commercial blood cell separation systems versus tube centrifugation methods for the preparation of platelet-rich plasma: A preliminary cross-sectional study. Rev. Assoc. Médica Bras. 2021, 67, 536–541. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volumetric Blood Flow [pL/s] | d1 | d3 | d5 | d7 | d10 | |
|---|---|---|---|---|---|---|
| Arterioles | ||||||
| prox. | Ctrl | 654 ± 108 | 871 ± 153 | 1081 ± 212 | 1150 ± 223 | 1503 ± 286 |
| MVF | 773 ± 192 | 1273 ± 216 | 1371 ± 272 | 1741 ± 232 | 1962 ± 429 | |
| med. | Ctrl | 367 ± 85 | 718 ± 168 | 736 ± 70 | 980 ± 160 | 1314 ± 247 |
| MVF | 612 ± 131 | 983 ± 166 | 1108 ± 169 | 1530 ± 167 * | 1885 ± 297 | |
| dist. | Ctrl | 16 ± 3 | - | - | - | - |
| MVF | 340 ± 88 * | 765 ± 164 | 924 ± 228 | 1282 ± 312 | 1543 ± 299 | |
| Capillaries | ||||||
| prox. | Ctrl | 3 ± 0 | 4 ± 0 | 6 ± 0 | 5 ± 0 | 7 ± 0 |
| MVF | 3 ± 0 | 4 ± 0 | 5 ± 0 | 5 ± 0 | 6 ± 1 | |
| med. | Ctrl | 3 ± 0 | 4 ± 0 | 5 ± 0 | 5 ± 0 | 6 ± 0 |
| MVF | 3 ± 0 | 5 ± 0 | 5 ± 0 | 6 ± 0 | 6 ± 1 | |
| dist. | Ctrl | 0 ± 0 | - | - | - | - |
| MVF | 2 ± 0 * | 8 ± 5 | 4 ± 0 | 5 ± 0 | 6 ± 1 | |
| Venules | ||||||
| prox. | Ctrl | 518 ± 99 | 758 ± 160 | 1300 ± 279 | 1738 ± 468 | 1929 ± 782 |
| MVF | 572 ± 76 | 1027 ± 197 | 1093 ± 213 | 2081 ± 640 | 1500 ± 368 | |
| med. | Ctrl | 272 ± 88 | 711 ± 182 | 908 ± 234 | 1348 ± 251 | 1504 ± 291 |
| MVF | 458 ± 102 | 1034 ± 218 | 1549 ± 299 | 2190 ± 443 | 2232 ± 580 | |
| dist. | Ctrl | 65 ± 38 | - | - | - | - |
| MVF | 239 ± 31 * | 877 ± 272 | 1123 ± 182 | 2053 ± 528 | 2205 ± 515 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weinzierl, A.; Harder, Y.; Schmauss, D.; Menger, M.D.; Laschke, M.W. Microvascular Fragments Protect Ischemic Musculocutaneous Flap Tissue from Necrosis by Improving Nutritive Tissue Perfusion and Suppressing Apoptosis. Biomedicines 2023, 11, 1454. https://doi.org/10.3390/biomedicines11051454
Weinzierl A, Harder Y, Schmauss D, Menger MD, Laschke MW. Microvascular Fragments Protect Ischemic Musculocutaneous Flap Tissue from Necrosis by Improving Nutritive Tissue Perfusion and Suppressing Apoptosis. Biomedicines. 2023; 11(5):1454. https://doi.org/10.3390/biomedicines11051454
Chicago/Turabian StyleWeinzierl, Andrea, Yves Harder, Daniel Schmauss, Michael D. Menger, and Matthias W. Laschke. 2023. "Microvascular Fragments Protect Ischemic Musculocutaneous Flap Tissue from Necrosis by Improving Nutritive Tissue Perfusion and Suppressing Apoptosis" Biomedicines 11, no. 5: 1454. https://doi.org/10.3390/biomedicines11051454