
Revisiting Resveratrol as an Osteoprotective Agent: Molecular Evidence from In Vivo and In Vitro Studies

Abstract
:
1. Introduction
2. RSV Effects on Bone: In Vivo Evidence
2.1. The Effects of RSV on Osteoblast-Bone Forming Cells
2.2. The Effects of RSV on Osteoclast-Bone Resorbing Cells
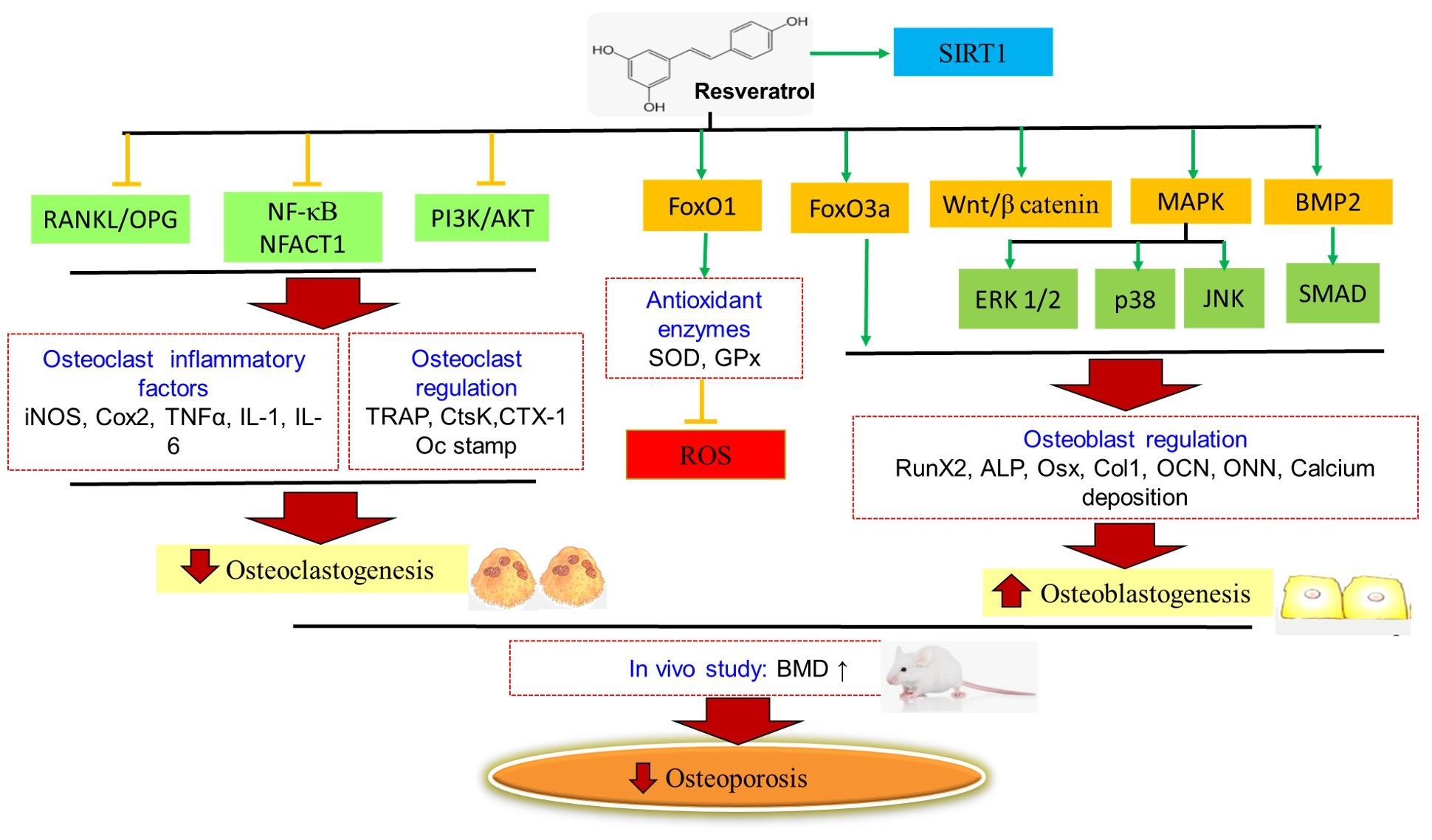
3. The Underlying Mechanisms of Action of Resveratrol as a Osteoprotective Age
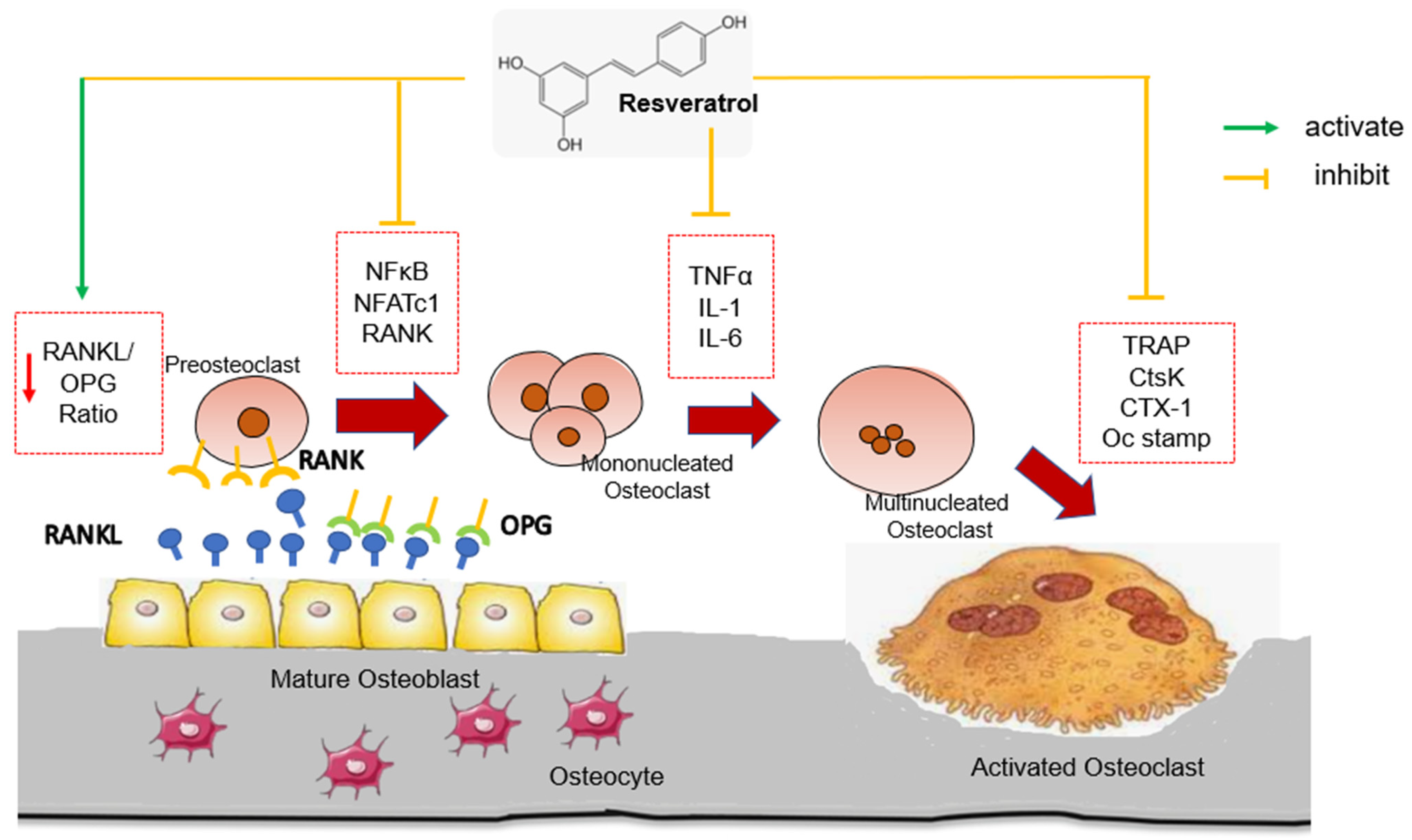
3.1. Regulating the RANK/RANKL/OPG System
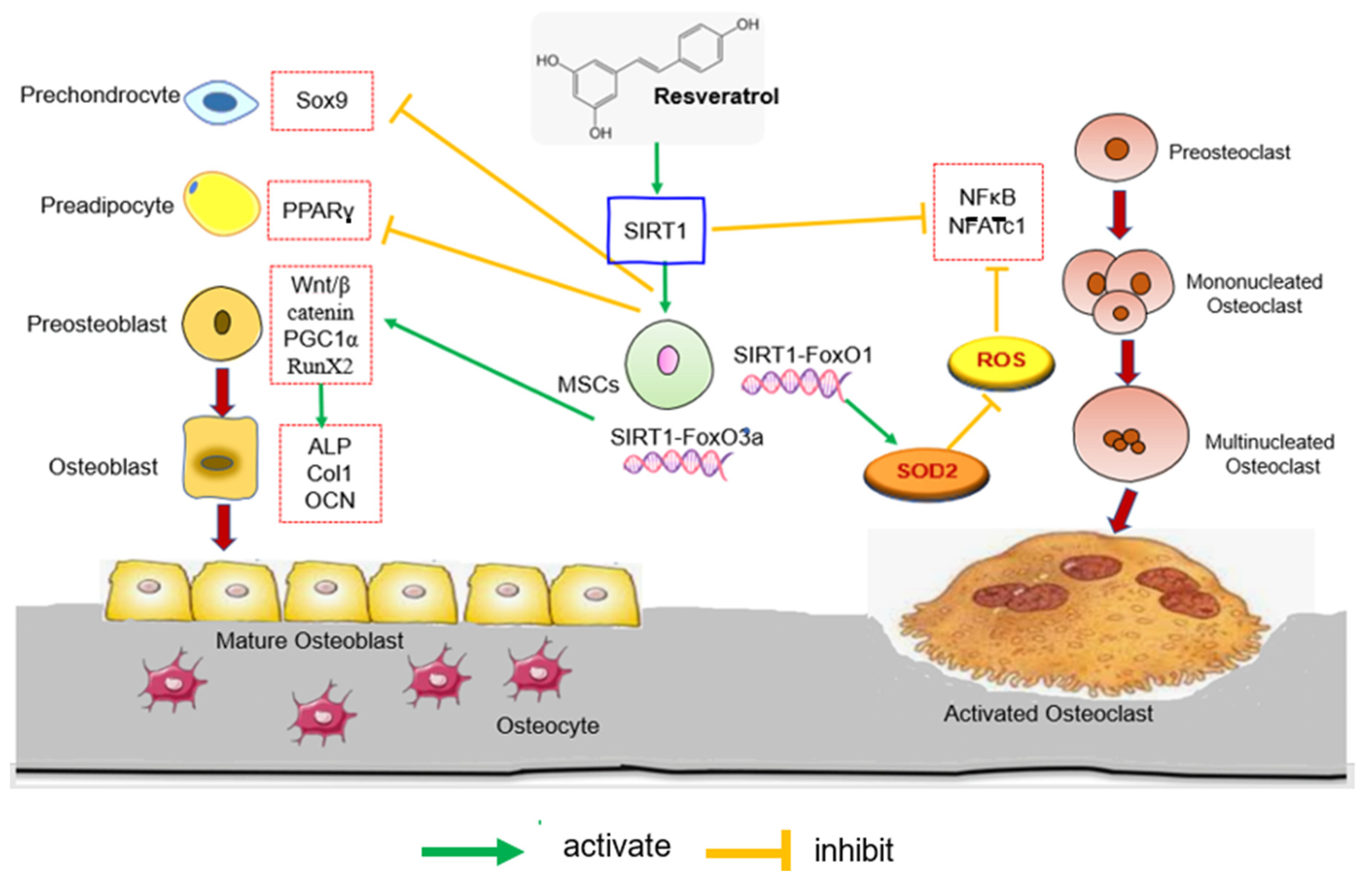
3.2. Regulating the SIRT1 and Wnt/β-Catenin Signalling
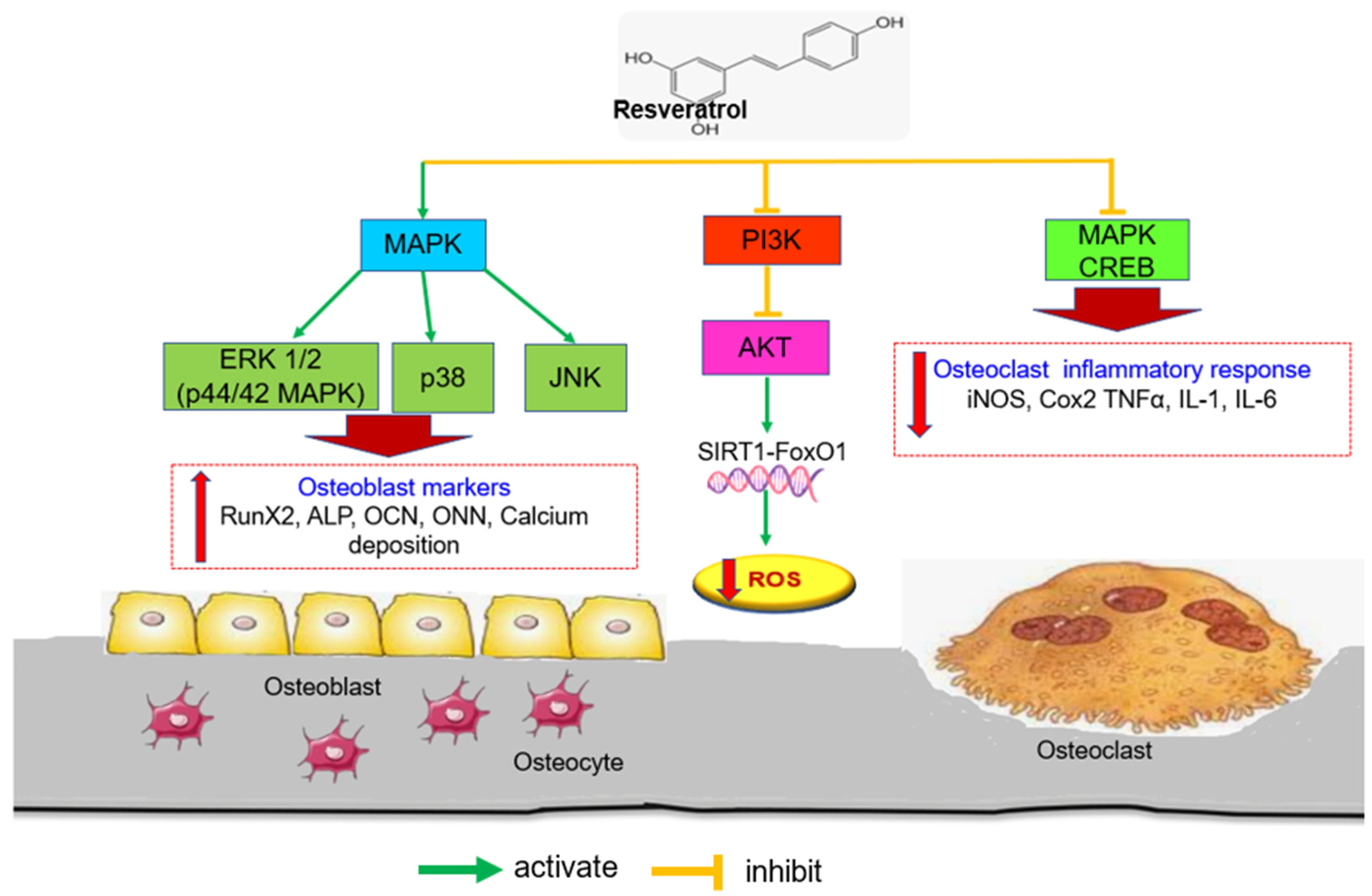
3.3. Regulating the MAPKs/JNK/ERK and PI3K/AKT Signaling
3.4. Regulating microRNA and BMP2 Signalling
3.5. Induction of Osteoclastogenesis by RANKL-RANK, NF-κB and NFATc1 Signaling
4. Bioavailability and Clinical Perspective of RSV
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BMD | Bone mineral density |
| BV/TV | bone volume/Total volume |
| Tb.Ar | Trabecular Area |
| Tb.Th | Trabecular thickness |
| Tb.N | Trabecular Number |
| Tb.Sp | Trabecular space |
| Ct. Vol | Cortical Volume |
| Ct. Th | Cortical Thickness |
| ALP | Alkaline phosphatase |
| OCN | Osteocalcin |
| SIRT1 | silent information regulator of transcription 1 |
| Akt | Ak strain transforming |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| TNFα | tumour necrosis factor alpha |
| ROS | Reactive Oxygen Species |
| MDA | Malondialdehyde |
| SOD | Superoxide dismutase |
| GSH | Glutathione |
| FoxO1 | Forkhead box O1 |
| PI3K/Akt | phosphatidylinositol-3-kinase |
| RANK | Receptor activator of nuclear factor κB |
| RANKL | receptor activator of nuclear factor-κB ligand |
| OPG | osteoprotegrin |
| TRAP | tartrate-resistant acid phosphatase |
| IL-6 | Interleukin-6 |
| IL-1β | Interleukin-1beta |
| CTX-1 | C-terminal telopeptides type I collagen |
| CtsK | Cathepsin K |
| NFATc1 | nuclear factor-activated T cells |
| Nrf2 | nuclear factor E2-related factor 2 |
| BMP2 | bone morphogenic protein 2 |
| Osx | Osterix |
| Col1a1 | alpha-1 type I collagen |
| P1NP | amino-terminal propeptide of type 1 procollagen |
| RunX2 | runt-related transcription factor 2 |
| BMSC | bone mesenchymal stem cells; |
| HBMSC | Human bone mesenchymal stem cells |
| PGE2 | prostaglandin E2 |
| MAP kinase | mitogen-activated protein kinase |
| SAPK/JNK | stress-activated protein kinase/c-Jun N-terminal kinase |
| AMPK | AMP-activated protein kinase |
| ONN | Osteonectin |
| OCN | Osteocalcin |
| ONN | Osteonectin |
| MMP | matrix metalloproteinase |
| GSK-3b | Glycogen synthase kinase-3 beta |
| ERK | extracellular signal-regulated kinase |
| CREB | cyclic AMP-responsive element-binding protein |
| LPS | Lipopolysaccharide |
| NO | nitric oxide |
| iNOS | inducible nitric oxide synthase |
| COX-2 | cyclooxygenase 2 |
| LDH | Lactate Dehydrogenase |
| PINP | Procollagen type I N-terminal propeptide |
References
- Liu, P.; Liang, X.; Li, Z.; Zhu, X.; Zhang, Z.; Cai, L. Decoupled Effects of Bone Mass, Microarchitecture and Tissue Property on the Mechanical Deterioration of Osteoporotic Bones. Compos. Part B Eng. 2019, 177, 107436. [Google Scholar] [CrossRef]
- Terkawi, M.A.; Matsumae, G.; Shimizu, T.; Takahashi, D.; Kadoya, K.; Norimasa Iwasaki. Interplay between Inflammation and Pathological Bone Resorption: Insights into Recent Mechanisms and Pathways in Related Diseases for Future Perspectives. Int. J. Mol. Sci. 2022, 23, 1786. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Chen, L.-R.; Chen, K.-H. Osteoporosis Due to Hormone Imbalance: An Overview of the Effects of Estrogen Deficiency and Glucocorticoid Overuse on Bone Turnover. Int. J. Mol. Sci. 2022, 23, 1376. [Google Scholar] [CrossRef]
- Vita, F.; Gangemi, S.; Pioggia, G.; Trimarchi, F.; Di Mauro, D. Physical Activity and Post-Transcriptional Regulation of Aging Decay: Modulation of Pathways in Postmenopausal Osteoporosis. Medicina 2022, 58, 767. [Google Scholar] [CrossRef]
- Soen, S.; Kaku, M.; Okubo, N.; Onishi, Y.; Saito, K.; Kobayashi, M. Fracture Risk Associated with Glucocorticoid-Induced Osteoporosis in Japan. J. Bone Miner. Metab. 2022, 40, 636–647. [Google Scholar] [CrossRef]
- Weng, W.; Li, H.; Zhu, S. An Overlooked Bone Metabolic Disorder: Cigarette Smoking-Induced Osteoporosis. Genes 2022, 13, 806. [Google Scholar] [CrossRef]
- Llorente, I.; García-Castañeda, N.; Valero, C.; González-Álvaro, I.; Castañeda, S. Osteoporosis in Rheumatoid Arthritis: Dangerous Liaisons. Front. Med. 2020, 7, 601618. [Google Scholar] [CrossRef]
- Rhee, S.Y.; Yon, D.K.; Kwon, M.J.; Kim, J.H.; Kim, J.-H.; Bang, W.J.; Lee, J.W.; Kwon, B.-C.; Choi, H.G.; Min, C. Association between Metabolic Syndrome and Osteoporosis among Adults Aged 50 Years and Older: Using the National Health Information Database in South Korea. Arch. Osteoporos. 2022, 17, 124. [Google Scholar] [CrossRef]
- Tsai, K.-S. Pharmaceutical Treatment for Spinal Osteoporosis: Bisphosphonates, Denosumab, Serms and Teriparatide. Osteoporos. Spine Asian Perspect. 2021, 157–169. [Google Scholar]
- Black, D.M.; Condra, K.; Adams, A.L.; Eastell, R. Bisphosphonates and the Risk of Atypical Femur Fractures. Bone 2022, 156, 116297. [Google Scholar] [CrossRef]
- Lu, L.; Lu, L.; Zhang, J.; Li, J. Potential risks of rare serious adverse effects related to long-term use of bisphosphonates: An overview of systematic reviews. J. Clin. Pharm. Ther. 2020, 45, 45–51. [Google Scholar] [CrossRef]
- Vannala, V.; Palaian, S.; Shankar, P.R. Therapeutic dimensions of bisphosphonates: A clinical update. Int. J. Prev. Med. 2020, 11, 166. [Google Scholar]
- Burns, J.; Yokota, T.; Ashihara, H.; Lean, M.E.J.; Crozier, A. Plant Foods and Herbal Sources of Resveratrol. J. Agric. Food Chem. 2002, 50, 3337–3340. [Google Scholar] [CrossRef]
- Cui, B.; Wang, Y.; Jin, J.; Yang, Z.; Guo, R.; Li, X.; Yang, L.; Li, Z. Resveratrol Treats Uvb-Induced Photoaging by Anti-Mmp Expression, through Anti-Inflammatory, Antioxidant, Antiapoptotic Properties, Treats Photoaging by Upregulating Vegf-B Expression. Oxidative Med. Cell. Longev. 2022, 2022, 6037303. [Google Scholar] [CrossRef]
- de Sousa, R.S.; Vasconcelos, U. Recent Progresses on Inhibition of Bacterial Biofilms by Resveratrol: A Systematic Review Covering a Ten-Year Period. Appl. Microbiol. Theory Technol. 2022, 3, 12–19. [Google Scholar] [CrossRef]
- Wu, S.-X.; Xiong, R.-G.; Huang, S.-Y.; Zhou, D.-D.; Saimaiti, A.; Zhao, C.-N.; Shang, A.; Zhang, Y.-J.; Gan, R.-Y.; Li, H.-B. Effects and Mechanisms of Resveratrol for Prevention and Management of Cancers: An Updated Review. Crit. Rev. Food Sci. Nutr. 2022, 1–19. [Google Scholar] [CrossRef]
- Kobylka, P.; Kucinska, M.; Kujawski, J.; Lazewski, D.; Wierzchowski, M.; Murias, M. Resveratrol Analogues as Selective Estrogen Signaling Pathway Modulators: Structure–Activity Relationship. Molecules 2022, 27, 6973. [Google Scholar] [CrossRef]
- Santos, M.G.D.; Schimith, L.E.; André-Miral, C.; Muccillo-Baisch, A.L.; Arbo, B.D.; Hort, M.A. Neuroprotective Effects of Resveratrol in in Vivo and in Vitro Experimental Models of Parkinson’s Disease: A Systematic Review. Neurotox. Res. 2022, 40, 319–345. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Y.; Sun, J.; Gao, F.; Resveratrol, G.S. Novel Application by Preconditioning to Attenuate Myocardial Ischemia/Reperfusion Injury in Mice through Regulate Ampk Pathway and Autophagy Level. J. Cell. Mol. Med. 2022, 26, 4216–4229. [Google Scholar] [CrossRef]
- Hecker, A.; Schellnegger, M.; Hofmann, E.; Luze, H.; Nischwitz, S.P.; Kamolz, L.-P.; Kotzbeck, P. The Impact of Resveratrol on Skin Wound Healing, Scarring, Aging. Int. Wound J. 2022, 19, 9–28. [Google Scholar] [CrossRef]
- Inchingolo, A.D.; Inchingolo, A.M.; Malcangi, G.; Avantario, P.; Azzollini, D.; Buongiorno, S.; Viapiano, F.; Campanelli, M.; Ciocia, A.M.; De Leonardis, N. Curcumin and Quercetin Supplementation on Bone Metabolism—A Systematic Review. Nutrients 2022, 14, 3519. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.; Bhuyan, P.; Ganguly, M.; Hazarika, J. Resveratrol: An Anfi-Androgen for the Treatment of Prostate Cancer. J Oncology. 2022, 2, 1046. [Google Scholar]
- Oğuz, O.; Adan, A. Involvement of Sphingolipid Metabolism Enzymes in Resveratrol-Mediated Cytotoxicity in Philadelphia-Positive Acute Lymphoblastic Leukemia. Nutr. Cancer 2022, 74, 2508–2521. [Google Scholar] [CrossRef] [PubMed]
- Gowd, V.; Jori, C.; Chaudhary, A.A.; Rudayni, H.A.; Khan, R. Resveratrol and Resveratrol Nano-Delivery Systems in the Treatment of Inflammatory Bowel Disease. J. Nutr. Biochem. 2022, 109, 109101. [Google Scholar] [CrossRef]
- Lee, I.-T.; Lin, H.-C.; Huang, T.-H.; Tseng, C.-N.; Cheng, H.-T.; Huang, W.-C.; Cheng, C.-Y. Anti-Inflammatory Effect of Resveratrol Derivatives Via the Downregulation of Oxidative-Stress-Dependent and C-Src Transactivation Egfr Pathways on Rat Mesangial Cells. Antioxidants 2022, 11, 835. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, J.; Li, W.; Zhang, D.; Wang, Z.; Zhai, Y.; Yu, H.; Li, Z. Integrative Investigation of the Tf–Mirna Coregulatory Network Involved in the Inhibition of Breast Cancer Cell Proliferation by Resveratrol. FEBS Open Bio 2022, 12, 379–393. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, J.; Xiang, Z.; Zou, H.; Shao, X. Amelioration of Renal Injury by Resveratrol in a Rat Renal Transplantation Model Via Activation of the Sirt1/Nf-Κb Signaling Pathway. BioMed Res. Int. 2022, 2022, 7140961. [Google Scholar] [CrossRef]
- Peng, X.; Wang, J.; Peng, J.; Jiang, H.; Le, K. Resveratrol Improves Synaptic Plasticity in Hypoxic-Ischemic Brain Injury in Neonatal Mice Via Alleviating Sirt1/Nf-Κb Signaling–Mediated Neuroinflammation. J. Mol. Neurosci. 2022, 72, 113–125. [Google Scholar] [CrossRef]
- Li, Y.; Tseng, W.-J.; de Bakker, C.M.J.; Zhao, H.; Chung, R.; Liu, X.S. Peak Trabecular Bone Microstructure Predicts Rate of Estrogen-Deficiency-Induced Bone Loss in Rats. Bone 2021, 145, 115862. [Google Scholar] [CrossRef]
- Pietschmann, P.; Skalicky, M.; Kneissel, M.; Rauner, M.; Hofbauer, G.; Stupphann, D.; Viidik, A. Bone Structure and Metabolism in a Rodent Model of Male Senile Osteoporosis. Exp. Gerontol. 2007, 42, 1099–1108. [Google Scholar] [CrossRef]
- David, V.; Martin, A.; Lafage-Proust, M.-H.; Malaval, L.; Peyroche, S.; Jones, D.B.; Vico, L.; Guignandon, A. Mechanical Loading Down-Regulates Peroxisome Proliferator-Activated Receptor Γ in Bone Marrow Stromal Cells and Favors Osteoblastogenesis at the Expense of Adipogenesis. Endocrinology 2007, 148, 2553–2562. [Google Scholar] [CrossRef] [PubMed]
- Bettis, T.; Kim, B.-J.; Hamrick, M.W. Impact of Muscle Atrophy on Bone Metabolism and Bone Strength: Implications for Muscle-Bone Crosstalk with Aging and Disuse. Osteoporos. Int. 2018, 29, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Deng, L.; Fan, J.; Zhang, Y. Effects of Resveratrol on Bone Metabolism and Bone Turnover Related Indexes in Ovariectomized Osteoporosis Rats. Cell. Mol. Biol. 2020, 66, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Flores, L.E.; Nelson, S.; Waltman, N.; Kupzyk, K.; Lappe, J.; Mack, L.; Bilek, L.D. Examining Effects of Habitual Physical Activity and Body Composition on Bone Structure in Early Post-Menopausal Women: A Pqct Analysis. Osteoporos. Int. 2021, 33, 425–433. [Google Scholar] [CrossRef]
- Zhao, H.; Li, X.; Li, N.; Liu, T.; Liu, J.; Li, Z.; Xiao, H.; Li, J. Long-Term Resveratrol Treatment Prevents Ovariectomy-Induced Osteopenia in Rats without Hyperplastic Effects on the Uterus. Br. J. Nutr. 2014, 111, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Khera, A.; Kanta, P.; Kalra, J.; Dumir, D. Resveratrol Restores the Level of Key Inflammatory Cytokines and Rankl/Opg Ratio in the Femur of Rat Osteoporosis Model. J. Women Aging 2019, 31, 540–552. [Google Scholar] [CrossRef]
- Feng, Y.-L.; Jiang, X.-T.; Ma, F.-F.; Han, J.; Tang, X.-L. Resveratrol Prevents Osteoporosis by Upregulating Foxo1 Transcriptional Activity. Int. J. Mol. Med. 2018, 41, 202–212. [Google Scholar] [CrossRef]
- Jiang, Y.; Luo, W.; Wang, B.; Wang, X.; Gong, P.; Xiong, Y. Resveratrol Promotes Osteogenesis Via Activating Sirt1/Foxo1 Pathway in Osteoporosis Mice. Life Sci. 2020, 246, 117422. [Google Scholar] [CrossRef]
- Feng, J.; Liu, S.; Ma, S.; Zhao, J.; Zhang, W.; Qi, W.; Cao, P.; Wang, Z.; Lei, W. Protective Effects of Resveratrol on Postmenopausal Osteoporosis: Regulation of Sirt1-Nf-Κb Signaling Pathway. Acta Biochim. Biophys. Sin. 2014, 46, 1024–1033. [Google Scholar] [CrossRef]
- Dana, P.M.; Sadoughi, F.; Mansournia, M.A.; Mirzaei, H.; Asemi, Z.; Yousefi, B. Targeting Wnt Signaling Pathway by Polyphenols: Implication for Aging and Age-Related Diseases. Biogerontology 2021, 22, 479–494. [Google Scholar] [CrossRef]
- Gao, Y.; Patil, S.; Qian, A. The Role of Micrornas in Bone Metabolism and Disease. Int. J. Mol. Sci. 2020, 21, 6081. [Google Scholar] [CrossRef] [PubMed]
- Elseweidy, M.M.; El-Swefy, S.E.; Shaheen, M.A.; Baraka, N.M.; Hammad, S.K. Effect of Resveratrol and Mesenchymal Stem Cell Monotherapy and Combined Treatment in Management of Osteoporosis in Ovariectomized Rats: Role of Sirt1/Foxo3a and Wnt/Β-Catenin Pathways. Arch. Biochem. Biophys. 2021, 703, 108856. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.-P.; Wang, X.-K.; Jiang, K.; Yin, C.; Xiang, C.; Wang, Y.; Pu, C.; Chen, L.; Li, Y.-L. Β-Ecdysterone Enhanced Bone Regeneration through the Bmp-2/Smad/Runx2/Osterix Signaling Pathway. Front. Cell Dev. Biol. 2022, 10, 883228. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-L.; Xiao, F.; Wang, C.-D.; Zhu, J.-F.; Shen, C.; Zuo, B.; Wang, H.; Li, D.; Wang, X.-Y.; Feng, W.-J. Gremlin2 Suppression Increases the Bmp-2-Induced Osteogenesis of Human Bone Marrow-Derived Mesenchymal Stem Cells Via the Bmp-2/Smad/Runx2 Signaling Pathway. J. Cell. Biochem. 2017, 118, 286–297. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, M.-W.; He, Y.; Deng, N.; Chen, Y.; Huang, J.; Xie, W. Protective Effect of Resveratrol on Estrogen Deficiency-Induced Osteoporosis Though Attenuating Nadph Oxidase 4/Nuclear Factor Kappa B Pathway by Increasing Mir-92b-3p Expression. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420941762. [Google Scholar] [CrossRef]
- Guo, W.; Han, X.; Cong, L.; Liang, D.; Tu, G.-J. Resveratrol Prevents Osteoporosis in Ovariectomized Rats by Regulating Microrna-338-3p. Mol. Med. Rep. 2015, 12, 2098–2106. [Google Scholar] [CrossRef]
- Li, W.; He, P.; Huang, Y.; Li, Y.-F.; Lu, J.; Li, M.; Kurihara, H.; Luo, Z.; Meng, T.; Onishi, M. Selective Autophagy of Intracellular Organelles: Recent Research Advances. Theranostics 2021, 11, 222. [Google Scholar] [CrossRef]
- Behera, J.; Ison, J.; Tyagi, A.; Mbalaviele, G.; Tyagi, N. Mechanisms of Autophagy and Mitophagy in Skeletal Development, Diseases and Therapeutics. Life Sci. 2022, 301, 120595. [Google Scholar] [CrossRef]
- Wang, S.; Deng, Z.; Ma, Y.; Jin, J.; Qi, F.; Li, S.; Liu, C.; Lyu, F.-J.; Zheng, Q. The Role of Autophagy and Mitophagy in Bone Metabolic Disorders. Int. J. Biol. Sci. 2020, 16, 2675. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, L.-M.; Guo, C.; Han, J.-F. Resveratrol Promotes Osteoblastic Differentiation in a Rat Model of Postmenopausal Osteoporosis by Regulating Autophagy. Nutr. Metab. 2020, 17, 29. [Google Scholar] [CrossRef]
- Hawkins, F.; Garla, V.; Allo, G.; Males, D.; Mola, L.; Corpas, E. Senile and Postmenopausal Osteoporosis: Pathophysiology, Diagnosis, Treatment. In Endocrinology of Aging; Elsevier: Amsterdam, The Netherlands, 2021; pp. 131–169. [Google Scholar]
- Föger-Samwald, U.; Kerschan-Schindl, K.; Butylina, M.; Pietschmann, P. Age Related Osteoporosis: Targeting Cellular Senescence. Int. J. Mol. Sci. 2022, 23, 2701. [Google Scholar] [CrossRef] [PubMed]
- Tresguerres, I.F.; Tamimi, F.; Eimar, H.; Barralet, J.; Torres, J.; Blanco, L.; Tresguerres, J.A.F. Resveratrol as Anti-Aging Therapy for Age-Related Bone Loss. Rejuvenation Res. 2014, 17, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Ameen, O.; Yassien, R.I.; Naguib, Y.M. Activation of Foxo1/Sirt1/Rankl/Opg Pathway May Underlie the Therapeutic Effects of Resveratrol on Aging-Dependent Male Osteoporosis. BMC Musculoskelet. Disord. 2020, 21, 375. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.M.C.; Shandala, T.; Nguyen, L.; Muhlhausler, B.S.; Chen, K.-M.; Howe, P.R.; Xian, C.J. Effects of Resveratrol Supplementation on Bone Growth in Young Rats and Microarchitecture and Remodeling in Ageing Rats. Nutrients 2014, 6, 5871–5887. [Google Scholar] [CrossRef] [PubMed]
- Yousefzadeh, N.; Kashfi, K.; Jeddi, S.; Ghasemi, A. Ovariectomized Rat Model of Osteoporosis: A Practical Guide. EXCLI J. 2020, 19, 89. [Google Scholar]
- Chowdhury, P.; Long, A.; Harris, G.; Soulsby, M.E.; Dobretsov, M. Animal Model of Simulated Microgravity: A Comparative Study of Hindlimb Unloading Via Tail Versus Pelvic Suspension. Physiol. Rep. 2013, 1, e00012. [Google Scholar] [CrossRef]
- Durbin, S.M.; Jackson, J.R.; Ryan, M.J.; Gigliotti, J.C.; Alway, S.E.; Tou, J.C. Resveratrol Supplementation Preserves Long Bone Mass, Microstructure, Strength in Hindlimb-Suspended Old Male Rats. J. Bone Miner. Metab. 2014, 32, 38–47. [Google Scholar] [CrossRef]
- Ebeling, P.R.; Nguyen, H.H.; Aleksova, J.; Vincent, A.J.; Wong, P. Secondary Osteoporosis. Endocr. Rev. 2022, 43, 240–313. [Google Scholar] [CrossRef]
- Yang, X.; Jiang, T.; Wang, Y.; Guo, L. The Role and Mechanism of Sirt1 in Resveratrol-Regulated Osteoblast Autophagy in Osteoporosis Rats. Sci. Rep. 2019, 9, 18424. [Google Scholar] [CrossRef]
- El Miedany, Y. New Horizons in Osteoporosis Management; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Lee, A.M.C.; Shandala, T.; Soo, P.P.; Su, Y.-W.; King, T.J.; Chen, K.-M.; Howe, P.R.; Xian, C.J. Effects of Resveratrol Supplementation on Methotrexate Chemotherapy-Induced Bone Loss. Nutrients 2017, 9, 255. [Google Scholar] [CrossRef]
- Gardner, O.F.W.; Alini, M.; Stoddart, M.J. Mesenchymal Stem Cells Derived from Human Bone Marrow. Cartil. Tissue Eng. Methods Protoc. 2015, 1340, 41–52. [Google Scholar]
- Dosier, C.R.; Erdman, C.P.; Park, J.H.; Schwartz, Z.; Boyan, B.D.; Guldberg, R.E. Resveratrol Effect on Osteogenic Differentiation of Rat and Human Adipose Derived Stem Cells in a 3-D Culture Environment. J. Mech. Behav. Biomed. Mater. 2012, 11, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Li, Y.; Quarles, L.D.; Song, T.; Pan, W.; Zhou, H.; Xiao, Z. Resveratrol Enhances Proliferation and Osteoblastic Differentiation in Human Mesenchymal Stem Cells Via Er-Dependent Erk1/2 Activation. Phytomedicine 2007, 14, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.-E.; Yang, Z.; Zhang, H.; Yao, G.; Liu, J.; Wei, Q.; Ma, B. Resveratrol Promotes Osteogenic Differentiation of Canine Bone Marrow Mesenchymal Stem Cells through Wnt/Beta-Catenin Signaling Pathway. Cell. Reprogramming 2018, 20, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Khademi-Shirvan, M.; Ghorbaninejad, M.; Hosseini, S.; Baghaban Eslaminejad, M. The Importance of Stem Cell Senescence in Regenerative Medicine. Adv Exp Med Biol. 2020, 1288, 87–102. [Google Scholar]
- 68. Davalli, P.; Tijana, M.; Andrea, C.; Angela, L.; Domenico, D.A. ROS, cell senescence, and novel molecular mechanisms in aging and age-related diseases. Oxidative Med. Cell. Longev. 2016, 2016, 3565127. [Google Scholar] [CrossRef]
- Atashi, F.; Modarressi, A.; Pepper, M.S. The Role of Reactive Oxygen Species in Mesenchymal Stem Cell Adipogenic and Osteogenic Differentiation: A Review. Stem Cells Dev. 2015, 24, 1150–1163. [Google Scholar] [CrossRef]
- Zhou, T.; Yan, Y.; Zhao, C.; Xu, Y.; Wang, Q.; Xu, N. Resveratrol Improves Osteogenic Differentiation of Senescent Bone Mesenchymal Stem Cells through Inhibiting Endogenous Reactive Oxygen Species Production Via Ampk Activation. Redox Rep. 2019, 24, 62–69. [Google Scholar] [CrossRef]
- Heher, P.; Ganassi, M.; Weidinger, A.; Engquist, E.N.; Pruller, J.; Nguyen, T.H.; Tassin, A.; Decleves, A.-E.; Mamchaoui, K.; Banerji, C.R.S. Interplay between Mitochondrial Reactive Oxygen Species, Oxidative Stress and Hypoxic Adaptation in Facioscapulohumeral Muscular Dystrophy: Metabolic Stress as Potential Therapeutic Target. Redox Biol. 2022, 51, 102251. [Google Scholar] [CrossRef]
- Yan, C.; Wang, Z.; Liu, W.; Pu, L.; Li, R.; Ai, C.; Xu, H.; Zhang, B.; Wang, T.; Zhang, X. Resveratrol Ameliorates High Altitude Hypoxia-Induced Osteoporosis by Suppressing the Ros/Hif Signaling Pathway. Molecules 2022, 27, 5538. [Google Scholar] [CrossRef]
- He, N.; Zhu, X.; He, W.; Zhao, S.; Zhao, W.; Zhu, C. Resveratrol Inhibits the Hydrogen Dioxide-Induced Apoptosis Via Sirt 1 Activation in Osteoblast Cells. Biosci. Biotechnol. Biochem. 2015, 79, 1779–1786. [Google Scholar] [CrossRef] [PubMed]
- Mei, W.; Song, D.; Wu, Z.; Yang, L.; Wang, P.; Zhang, R.; Zhu, X. Resveratrol Protects Mc3t3-E1 Cells against Cadmium-Induced Suppression of Osteogenic Differentiation by Modulating the Erk1/2 and Jnk Pathways. Ecotoxicol. Environ. Saf. 2021, 214, 112080. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.D.; Beyer, M.; Krause-Buchholz, U.; Wobus, M.; Bornhäuser, M.; Rödel, G. Oxphos Supercomplexes as a Hallmark of the Mitochondrial Phenotype of Adipogenic Differentiated Human Mscs. PLoS ONE 2012, 7, e35160. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Yang, Q.; Guo, D.; Xie, J.; Yang, Y.-S.; Chaugule, S.; DeSouza, N.; Oh, W.-T.; Li, R.; Chen, Z. Impaired Mitochondrial Oxidative Metabolism in Skeletal Progenitor Cells Leads to Musculoskeletal Disintegration. Nat. Commun. 2022, 13, 6869. [Google Scholar] [CrossRef]
- Babaei-Abraki, S.; Karamali, F.; Nasr-Esfahani, M.H. The Role of Endoplasmic Reticulum and Mitochondria in Maintaining Redox Status and Glycolytic Metabolism in Pluripotent Stem Cells. Stem Cell Rev. Rep. 2022, 18, 1789–1808. [Google Scholar] [CrossRef]
- Garone, C.; Pietra, A.; Nesci, S. From the Structural and (Dys) Function of Atp Synthase to Deficiency in Age-Related Diseases. Life 2022, 12, 401. [Google Scholar] [CrossRef]
- Moon, D.K.; Kim, B.G.; Lee, A.R.; Choe, Y.I.; Khan, I.; Moon, K.M.; Jeon, R.-H.; Byun, J.-H.; Hwang, S.-C.; Woo, D.K. Resveratrol Can Enhance Osteogenic Differentiation and Mitochondrial Biogenesis from Human Periosteum-Derived Mesenchymal Stem Cells. J. Orthop. Surg. Res. 2020, 15, 203. [Google Scholar] [CrossRef]
- Du, L.; Lei, X.; Wang, J.; Wang, L.; Zhong, Q.; Fang, X.; Li, P.; Du, B.; Wang, Y.; Liao, Z. Lipopolysaccharides Derived from Gram-Negative Bacterial Pool of Human Gut Microbiota Promote Inflammation and Obesity Development. Int. Rev. Immunol. 2022, 41, 45–56. [Google Scholar] [CrossRef]
- Al Zahrani, S.; Shinwari, Z.; Gaafar, A.; Alaiya, A.; Al-Kahtani, A. Anti-Inflammatory Effect of Specialized Proresolving Lipid Mediators on Mesenchymal Stem Cells: An In Vitro Study. Cells 2022, 12, 122. [Google Scholar] [CrossRef]
- Zhou, C.; Gao, J.; Qu, H.; Xu, L.; Zhang, B.; Guo, Q.; Jing, F. Anti-Inflammatory Mechanism of Action of Benzoylmesaconine in Lipopolysaccharide-Stimulated Raw264. 7 Cells. Evid.-Based Complement. Altern. Med. 2022, 2022, 7008907. [Google Scholar]
- Jayashree, B.; Bibin, Y.S.; Prabhu, D.; Shanthirani, C.S.; Gokulakrishnan, K.; Lakshmi, B.S.; Mohan, V.; Balasubramanyam, M. Increased Circulatory Levels of Lipopolysaccharide (Lps) and Zonulin Signify Novel Biomarkers of Proinflammation in Patients with Type 2 Diabetes. Mol. Cell. Biochem. 2014, 388, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Banyard, A.; Mullan, A.; Sriskantharajah, S.; Southworth, T.; Singh, D. Characterization of the Inflammatory Response to Inhaled Lipopolysaccharide in Mild to Moderate Chronic Obstructive Pulmonary Disease. Br. J. Clin. Pharmacol. 2015, 79, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Carla, R.A.B.; Giovanni, F.G.; Eduardo, C.-J.; Bernd, L.F.; Antonio, C.P.D.O. Lipopolysaccharide-Induced Neuroinflammation as a Bridge to Understand Neurodegeneration. Int. J. Mol. Sci. 2019, 20, 2293. [Google Scholar]
- Jin, M.; Nie, J.; Zhu, J.; Li, J.; Fang, T.; Xu, J.; Jiang, X.; Chen, Z.; Li, J.; Wu, F. Acacetin Inhibits Rankl-Induced Osteoclastogenesis and Lps-Induced Bone Loss by Modulating Nfatc1 Transcription. Biochem. Biophys. Res. Commun. 2021, 583, 146–153. [Google Scholar] [CrossRef]
- Ma, J.; Wang, Z.; Zhao, J.; Miao, W.; Ye, T.; Chen, A. Resveratrol Attenuates Lipopolysaccharides (Lps)-Induced Inhibition of Osteoblast Differentiation in Mc3t3-E1 Cells. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 2045. [Google Scholar] [CrossRef] [PubMed]
- Ornstrup, M.J.; Harsløf, T.; Sørensen, L.; Stenkjær, L.; Langdahl, B.L.; Pedersen, S.B. Resveratrol Increases Osteoblast Differentiation in Vitro Independently of Inflammation. Calcif. Tissue Int. 2016, 99, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Vergara-Hernez, F.B.; Nielsen, B.D.; Colbath, A.C. Is the Use of Bisphosphonates Putting Horses at Risk? An Osteoclast Perspective. Animals 2022, 12, 1722. [Google Scholar] [CrossRef] [PubMed]
- João, B.B.W.; Renata, S.C.; Monique, E.P. Efficacy of Laser Therapy in the Management of Bisphosphonate-Related Osteonecrosis of the Jaw (Bronj): A Systematic Review. Lasers Med. Sci. 2016, 31, 1261–1272. [Google Scholar]
- Borsani, E.; Bonazza, V.; Buffoli, B.; Nocini, P.F.; Albanese, M.; Zotti, F.; Inchingolo, F.; Rezzani, R.; Rodella, L.F. Beneficial Effects of Concentrated Growth Factors and Resveratrol on Human Osteoblasts in Vitro Treated with Bisphosphonates. BioMed Res. Int. 2018, 2018, 4597321. [Google Scholar] [CrossRef]
- Jee, H.-K.; Jeon, W.-Y.; Kwak, H.-W.; Seok, H. Long-Term Changes in Adipose Tissue in the Newly Formed Bone Induced by Recombinant Human Bmp-2 In Vivo. Biomimetics 2023, 8, 33. [Google Scholar] [CrossRef]
- Hikiji, H.; Takato, T.; Shimizu, T.; Ishii, S. The Roles of Prostanoids, Leukotrienes, Platelet-Activating Factor in Bone Metabolism and Disease. Prog. Lipid Res. 2008, 47, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Harvey, W. Prostaglandins and the Mechanism of Bone Resorption. In Prostaglandins in Bone Resorption; CRC Press: Boca Raton, FL, USA, 2020; pp. 43–56. [Google Scholar]
- Agas, D.; Marchetti, L.; Hurley, M.M.; Sabbieti, M.G. Prostaglandin F2α: A Bone Remodeling Mediator. J. Cell. Physiol. 2013, 228, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-H.; Zhu, D.; Yang, T.; Cheng, L.; Sun, J.; Tan, L. Crosstalk between the Cox2-Pge2-Ep4 Signaling Pathway and Primary Cilia in Osteoblasts after Mechanical Stimulation. J. Cell. Physiol. 2021, 236, 4764–4777. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Tokuda, H.; Tanabe, K.; Yamaguchi, S.; Hioki, T.; Tachi, J.; Matsushima-Nishiwaki, R.; Kozawa, O.; Iida, H. Acetaminophen Reduces Osteoprotegerin Synthesis Stimulated by Pge2 and Pgf2α in Osteoblasts: Attenuation of Sapk/Jnk but Not P38 Mapk or P44/P42 Mapk. Biomed. Res. 2021, 42, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Tokuda, H.; Kuroyanagi, G.; Mizutani, J.; Matsushima-Nishiwaki, R.; Kondo, A.; Kozawa, O.; Otsuka, T. Regulation by Resveratrol of Prostaglandin E2-Stimulated Osteoprotegerin Synthesis in Osteoblasts. Int. J. Mol. Med. 2014, 34, 1439–1445. [Google Scholar] [CrossRef]
- Kuroyanagi, G.; Tokuda, H.; Matsushima-Nishiwaki, R.; Kondo, A.; Mizutani, J.; Kozawa, O.; Otsuka, T. Resveratrol Suppresses Prostaglandin F2α-Induced Osteoprotegerin Synthesis in Osteoblasts: Inhibition of the Map Kinase Signaling. Arch. Biochem. Biophys. 2014, 542, 39–45. [Google Scholar] [CrossRef]
- Jing, Y.; Hu, T.; Lin, C.; Xiong, Q.; Liu, F.; Yuan, J.; Zhao, X.; Wang, R. Resveratrol Downregulates Pcsk9 Expression and Attenuates Steatosis through Estrogen Receptor A-Mediated Pathway in L02 Cells. Eur. J. Pharmacol. 2019, 855, 216–226. [Google Scholar] [CrossRef]
- Solak, K.A.; Wijnolts, F.M.J.; Nijmeijer, S.M.; Blaauboer, B.J.; van den Berg, M.; van Duursen, M.B.M. Excessive Levels of Diverse Phytoestrogens Can Modulate Steroidogenesis and Cell Migration of Kgn Human Granulosa-Derived Tumor Cells. Toxicol. Rep. 2014, 1, 360–372. [Google Scholar] [CrossRef]
- Shah, A.A.; Shah, A.; Kumar, A.; Lakra, A.; Singh, D.; Nayak, Y. Phytoestrogenic Potential of Resveratrol by Selective Activation of Estrogen Receptor-A in Osteoblast Cells. Rev. Bras. Farmacogn. 2022, 32, 248–256. [Google Scholar] [CrossRef]
- Tseng, P.-C.; Hou, S.-M.; Chen, R.-J.; Peng, H.-W.; Hsieh, C.-F.; Kuo, M.-L.; Yen, M.-L. Resveratrol Promotes Osteogenesis of Human Mesenchymal Stem Cells by Upregulating Runx2 Gene Expression Via the Sirt1/Foxo3a Axis. J. Bone Miner. Res. 2011, 26, 2552–2563. [Google Scholar] [CrossRef]
- Constanze, B.; Popper, B.; Aggarwal, B.B.; Shakibaei, M. Evidence That Tnf-Β Suppresses Osteoblast Differentiation of Mesenchymal Stem Cells and Resveratrol Reverses It through Modulation of Nf-Κb, Sirt1 and Runx. Cell Tissue Res. 2020, 381, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Sun, L.; Liu, B.; Deng, Y.-S.; Zhan, D.; Chen, Y.-L.; He, Y.; Liu, J.; Zhang, Z.-J.; Sun, J. Resveratrol Inhibits Lps-Induced Mapks Activation Via Activation of the Phosphatidylinositol 3-Kinase Pathway in Murine Raw 264.7 Macrophage Cells. PLoS ONE 2012, 7, e44107. [Google Scholar] [CrossRef] [PubMed]
- Poudel, S.; Martins, G.; Cancela, M.L.; Gavaia, P.J. Resveratrol-Mediated Reversal of Doxorubicin-Induced Osteoclast Differentiation. Int. J. Mol. Sci. 2022, 23, 15160. [Google Scholar] [CrossRef]
- Hooshiar, S.H.; Tobeiha, M.; Jafarnejad, S. Soy Isoflavones and Bone Health: Focus on the Rankl/Rank/Opg Pathway. BioMed Res. Int. 2022, 2022, 8862278. [Google Scholar] [CrossRef] [PubMed]
- Elson, A.; Anuj, A.; Barnea-Zohar, M.; Reuven, N. The Origins and Formation of Bone-Resorbing Osteoclasts. Bone 2022, 164, 116538. [Google Scholar] [CrossRef] [PubMed]
- Szwarc, M.M.; Hai, L.; Maurya, V.K.; Rajapakshe, K.; Perera, D.; Ittmann, M.M.; Mo, Q.; Lin, Y.; Bettini, M.L.; Coarfa, C. Histopathologic and Transcriptomic Phenotypes of a Conditional Rankl Transgenic Mouse Thymus. Cytokine 2022, 160, 156022. [Google Scholar] [CrossRef]
- Li, B.; Wang, P.; Jiao, J.; Wei, H.; Xu, W.; Zhou, P. Roles of the Rankl–Rank Axis in Immunity—Implications for Pathogenesis and Treatment of Bone Metastasis. Front. Immunol. 2022, 13, 824117. [Google Scholar] [CrossRef]
- Wang, C.; Ning, H.; Gao, J.; Xue, T.; Zhao, M.; Jiang, X.; Zhu, X.; Guo, X.; Li, H.; Wang, X. Disruption of Hematopoiesis Attenuates the Osteogenic Differentiation Capacity of Bone Marrow Stromal Cells. Stem Cell Res. Ther. 2022, 13, 27. [Google Scholar] [CrossRef]
- He, J.; Zheng, L.; Li, X.; Huang, F.; Hu, S.; Chen, L.; Jiang, M.; Lin, X.; Jiang, H.; Zeng, Y. Obacunone Targets Macrophage Migration Inhibitory Factor (Mif) to Impede Osteoclastogenesis and Alleviate Ovariectomy-Induced Bone Loss. J. Adv. Res. 2023, in press. [Google Scholar] [CrossRef]
- Casado-Díaz, A.; Rodríguez-Ramos, Á.; Torrecillas-Baena, B.; Dorado, G.; Quesada-Gómez, J.M.; Gálvez-Moreno, M.Á. Flavonoid Phloretin Inhibits Adipogenesis and Increases Opg Expression in Adipocytes Derived from Human Bone-Marrow Mesenchymal Stromal-Cells. Nutrients 2021, 13, 4185. [Google Scholar] [CrossRef]
- Wang, T.; He, C. Tnf-A and Il-6: The Link between Immune and Bone System. Curr. Drug Targets 2020, 21, 213–227. [Google Scholar] [PubMed]
- Chakraborty, D.; Sarkar, A.; Mann, S.; Agnihotri, P.; Saquib, M.; Malik, S.; Kumavat, R.; Mathur, A.; Biswas, S. Estrogen-Mediated Differential Protein Regulation and Signal Transduction in Rheumatoid Arthritis. J. Mol. Endocrinol. 2022, 69, R25–R43. [Google Scholar] [CrossRef] [PubMed]
- Adhyatmika, A.; Putri, K.S.S.; Gore, E.; Mangnus, K.A.; Reker-Smit, C.; Schuppan, D.; Beljaars, L.; Olinga, P.; Melgert, B.N. Osteoprotegerin Expression in Liver Is Induced by Il-13 through Tgf-Β. bioRxiv 2020. [Google Scholar] [CrossRef]
- Wang, X.; Li, Z.; Wang, Z.; Liu, H.; Cui, Y.; Liu, Y.; Ren, M.; Zhan, H.; Li, Z.; Wu, M. Incorporation of Bone Morphogenetic Protein-2 and Osteoprotegerin in 3d-Printed Ti6al4v Scaffolds Enhances Osseointegration under Osteoporotic Conditions. Front. Bioeng. Biotechnol. 2021, 9, 754205. [Google Scholar] [CrossRef]
- Carrillo-López; Natalia; Martínez-Arias, L.; Fernández-Villabrille, S.; Ruiz-Torres, M.P.; Dusso, A.; Cannata-Andía, J.B.; Naves-Díaz, M.; Panizo, S.; European Renal Osteodystrophy Workgroup. Role of the Rank/Rankl/Opg and Wnt/Β-Catenin Systems in Ckd Bone and Cardiovascular Disorders. Calcif. Tissue Int. 2021, 108, 439–451. [Google Scholar] [CrossRef]
- Jong, Y.; Sohn, H.M.; Jang, Y.; Park, M.; Kim, B.; Kim, B.; Park, J.-I.; Hyun, H.; Jeong, B.; Hong, C. A Novel Modified Rankl Variant Can Prevent Osteoporosis by Acting as a Vaccine and an Inhibitor. Clin. Transl. Med. 2021, 11, e368. [Google Scholar]
- Qu, H.; Zhang, Y.; He, R.; Lin, N.; Wang, C. Anethole Inhibits Rankl-Induced Osteoclastogenesis by Downregulating Erk/Akt Signaling and Prevents Ovariectomy-Induced Bone Loss in Vivo. Int. Immunopharmacol. 2021, 100, 108113. [Google Scholar] [CrossRef]
- Huang, X.-L.; Liu, C.; Shi, X.-M.; Cheng, Y.-T.; Zhou, Q.; Li, J.-P.; Liao, J. Zoledronic Acid Inhibits Osteoclastogenesis and Bone Resorptive Function by Suppressing Rankl-Mediated Nf-Κb and Jnk and Their Downstream Signalling Pathways. Mol. Med. Rep. 2022, 25, 59. [Google Scholar] [CrossRef]
- Min, H.; Morony, S.; Sarosi, I.; Dunstan, C.R.; Capparelli, C.; Scully, S.; Van, G.; Kaufman, S.; Kostenuik, P.J.; Lacey, D.L. Osteoprotegerin Reverses Osteoporosis by Inhibiting Endosteal Osteoclasts and Prevents Vascular Calcification by Blocking a Process Resembling Osteoclastogenesis. J. Exp. Med. 2000, 192, 463–474. [Google Scholar] [CrossRef]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Yasuda, H.; Fukuda, C. Osteoclast Differentiation by Rankl and Opg Signaling Pathways. J. Bone Miner. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef]
- Zhao, J.; Huang, M.; Zhang, X.; Xu, J.; Hu, G.; Zhao, X.; Cui, P.; Zhang, X. Mir-146a Deletion Protects from Bone Loss in Ovx Mice by Suppressing Rankl/Opg and M-Csf in Bone Microenvironment. J. Bone Miner. Res. 2019, 34, 2149–2161. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.D.; Sood, P.P. Osteoporosis: A Gender Based Analysis. J. Cell Tissue Res. 2022, 22, 7183–7186. [Google Scholar]
- Zhu, S.; He, H.; Gao, C.; Luo, G.; Xie, Y.; Wang, H.; Tian, L.; Chen, X.; Yu, X.; He, C. Ovariectomy-Induced Bone Loss in Tnfα and Il6 Gene Knockout Mice Is Regulated by Different Mechanisms. J. Mol. Endocrinol. 2018, 60, 185–198. [Google Scholar] [CrossRef]
- Jiang, N.; An, J.; Yang, K.; Liu, J.; Guan, C.; Ma, C.; Tang, X. Nlrp3 Inflammasome: A New Target for Prevention and Control of Osteoporosis? Front. Endocrinol. 2021, 12, 752546. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Getting, S.J.; Locke, I.C. Regulation of Tnf-Induced Osteoclast Differentiation. Cells 2021, 11, 132. [Google Scholar] [CrossRef] [PubMed]
- Garg, G.; Singh, S. Antiaging Strategies Based on Sirtuin Activation. In Emerging Anti-Aging Strategies; Springer: Berlin/Heidelberg, Germany, 2023; pp. 257–268. [Google Scholar]
- Chen, Y.E.; Zhou, F.; Liu, H.; Li, J.; Che, H.; Shen, J.; Luo, E. SIRT 1 a Promising Regulator of Bone Homeostasis. Life Sci. 2021, 269, 119041. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, M.; Ge, Y.; Wang, X. Sirt1 and Aging Related Signaling Pathways. Mech. Ageing Dev. 2020, 187, 111215. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Matsushita, T.; Takayama, K.; Matsumoto, T.; Nishida, K.; Kuroda, R.; Kurosaka, M. Disruption of Sirt1 in Chondrocytes Causes Accelerated Progression of Osteoarthritis under Mechanical Stress and During Ageing in Mice. Ann. Rheum. Dis. 2014, 73, 1397–1404. [Google Scholar] [CrossRef]
- Zainabadi, K.; Liu, C.J.; Caldwell, A.L.M.; Guarente, L. Sirt1 Is a Positive Regulator of in Vivo Bone Mass and a Therapeutic Target for Osteoporosis. PLoS ONE 2017, 12, e0185236. [Google Scholar] [CrossRef]
- El-Haj, M.; Gurt, I.; Cohen-Kfir, E.; Dixit, V.; Artsi, H.; Kandel, L.; Yakubovsky, O.; Safran, O.; Dresner-Pollak, R. Reduced Sirtuin1 Expression at the Femoral Neck in Women Who Sustained an Osteoporotic Hip Fracture. Osteoporos. Int. 2016, 27, 2373–2378. [Google Scholar] [CrossRef]
- Godfrin-Valnet, M.; Khan, K.A.; Guillot, X.; Prati, C.; Baud, L.; Abbas, W.; Toussirot, E.; Wendling, D.; Herbein, G. Sirtuin 1 Activity in Peripheral Blood Mononuclear Cells of Patients with Osteoporosis. Med. Sci. Monit. Basic Res. 2014, 20, 142. [Google Scholar] [PubMed]
- Ma, C.; Pi, C.; Yang, Y.; Lin, L.; Shi, Y.; Li, Y.; Li, Y.; He, X. Nampt Expression Decreases Age-Related Senescence in Rat Bone Marrow Mesenchymal Stem Cells by Targeting Sirt. PLoS ONE 2017, 12, e0170930. [Google Scholar]
- Kayvan, Z. The Variable Role of Sirt1 in the Maintenance and Differentiation of Mesenchymal Stem Cells. Regen. Med. 2018, 13, 343–356. [Google Scholar]
- Sun, W.; Qiao, W.; Zhou, B.; Hu, Z.; Yan, Q.; Wu, J.; Wang, R.; Zhang, Q.; Miao, D. Overexpression of Sirt1 in Mesenchymal Stem Cells Protects against Bone Loss in Mice by Foxo3a Deacetylation and Oxidative Stress Inhibition. Metabolism 2018, 88, 61–71. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhang, Y.; Zhou, F.; Liu, Y.; Yi, Z.; Gong, P.; Wu, Y. Foxo1 Differentially Regulates Bone Formation in Young and Aged Mice. Cell. Signal. 2022, 99, 110438. [Google Scholar] [CrossRef]
- Higuchi, M.; Dusting, G.J.; Peshavariya, H.; Jiang, F.; Hsiao, S.T.-F.; Chan, E.C.; Liu, G.-S. Differentiation of Human Adipose-Derived Stem Cells into Fat Involves Reactive Oxygen Species and Forkhead Box O1 Mediated Upregulation of Antioxidant Enzymes. Stem Cells Dev. 2013, 22, 878–888. [Google Scholar] [CrossRef]
- Iyer, S.; Han, L.; Ambrogini, E.; Yavropoulou, M.; Fowlkes, J.; Manolagas, S.C.; Almeida, M. Deletion of FoxO1, 3, and 4 in Osteoblast Progenitors Attenuates the Loss of Cancellous Bone Mass in a Mouse Model of Type 1 Diabetes. J. Bone Miner. Res. 2017, 32, 60–69. [Google Scholar] [CrossRef]
- Jalgaonkar, M.P.; Parmar, U.M.; Kulkarni, Y.A.; Oza, M.J. Sirt1-Foxos Activity Regulates Diabetic Complications. Pharmacol. Res. 2022, 175, 106014. [Google Scholar] [CrossRef]
- Li, Y.; Jin, D.; Xie, W.; Wen, L.; Chen, W.; Xu, J.; Ding, J.; Ren, D. Ppar-Γ and Wnt Regulate the Differentiation of Mscs into Adipocytes and Osteoblasts Respectively. Curr. Stem Cell Res. Ther. 2018, 13, 185–192. [Google Scholar] [CrossRef]
- Peter VN Bodine. Wnt Signaling Control of Bone Cell Apoptosis. Cell Res. 2008, 18, 248–253. [Google Scholar] [CrossRef]
- Baksh, D.; Tuan, R.S. Canonical and Non-Canonical Wnts Differentially Affect the Development Potential of Primary Isolate of Human Bone Marrow Mesenchymal Stem Cells. J. Cell. Physiol. 2007, 212, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Li, Y.; Feng, R.; He, P.; Zhang, Y. Γ-Tocotrienol Induced the Proliferation and Differentiation of Mc3t3-E1 Cells through the Stimulation of the Wnt/Β-Catenin Signaling Pathway. Food Funct. 2022, 13, 398–410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhou, X.; Hou, W.; Chen, E.; Ye, C.; Chen, M.; Lu, Q.; Yu, X.; Li, W. Reversing the Imbalance in Bone Homeostasis Via Sustained Release of Sirt-1 Agonist to Promote Bone Healing under Osteoporotic Condition. Bioact. Mater. 2023, 19, 429–443. [Google Scholar] [CrossRef]
- Kong, S.; Cai, B.; Nie, Q. Pgc-1α Affects Skeletal Muscle and Adipose Tissue Development by Regulating Mitochondrial Biogenesis. Mol. Genet. Genom. 2022, 297, 621–633. [Google Scholar] [CrossRef]
- Yao, H.; Yao, Z.; Zhang, S.; Zhang, W.; Zhou, W. Upregulation of Sirt1 Inhibits H2o2-Induced Osteoblast Apoptosis Via Foxo1/Β-Catenin Pathway. Mol. Med. Rep. 2018, 17, 6681–6690. [Google Scholar] [CrossRef]
- Liu, T.; Ma, X.; Ouyang, T.; Chen, H.; Lin, J.; Liu, J.; Xiao, Y.; Yu, J.; Huang, Y. Sirt1 Reverses Senescence Via Enhancing Autophagy and Attenuates Oxidative Stress-Induced Apoptosis through Promoting P53 Degradation. Int. J. Biol. Macromol. 2018, 117, 225–234. [Google Scholar] [CrossRef]
- Tao, H.; Ge, G.; Liang, X.; Zhang, W.; Sun, H.; Li, M.; Geng, D. Ros Signaling Cascades: Dual Regulations for Osteoclast and Osteoblast. Acta Biochim. Biophys. Sin. 2020, 52, 1055–1062. [Google Scholar] [CrossRef]
- Manzoor, Z.; Koh, Y.-S. Mitogen-Activated Protein Kinases in Inflammation. J. Bacteriol. Virol. 2012, 42, 189–195. [Google Scholar] [CrossRef]
- Yuan, J.; Dong, X.; Yap, J.; Hu, J. The Mapk and Ampk Signalings: Interplay and Implication in Targeted Cancer Therapy. J. Hematol. Oncol. 2020, 13, 59. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; López, J.M. Understanding Mapk Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef]
- Ba, P.; Duan, X.; Fu, G.; Lv, S.; Yang, P.; Sun, Q. Differential Effects of P38 and Erk1/2 on the Chondrogenic and Osteogenic Differentiation of Dental Pulp Stem Cells. Mol. Med. Rep. 2017, 16, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Semba, T.; Sammons, R.; Wang, X.; Xie, X.; Dalby, K.N.; Ueno, N.T. Jnk Signaling in Stem Cell Self-Renewal and Differentiation. Int. J. Mol. Sci. 2020, 21, 2613. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Huang, Z.; Yin, X.; Zhang, J.; Gong, L.; Chen, J.; Rong, K.; Xu, J.; Lu, L.; Cui, L. Role of C-Jun N-Terminal Kinase in the Osteogenic and Adipogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells. Exp. Cell Res. 2015, 339, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Galvão, I.; Sousa, L.P.; Teixeira, M.M.; Pinho, V. Pi3k Isoforms in Cell Signalling and Innate Immune Cell Responses. In Pi3k and Akt Isoforms in Immunity: Mechanisms and Therapeutic Opportunities; Springer: Berlin/Heidelberg, Germany, 2022; pp. 147–164. [Google Scholar]
- Gupta, S.; Kumar, M.; Chaudhuri, S.; Kumar, A. The Non-Canonical Nuclear Functions of Key Players of the Pi3k-Akt-Mtor Pathway. J. Cell. Physiol. 2022, 237, 3181–3204. [Google Scholar] [CrossRef]
- He, M.; Li, D.; Fang, C.; Xu, Q. Ythdf1 Regulates Endoplasmic Reticulum Stress, Nf-Κb, Mapk and Pi3k-Akt Signaling Pathways in Inflammatory Osteoclastogenesis. Arch. Biochem. Biophys. 2022, 732, 109464. [Google Scholar] [CrossRef]
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The Role of the Transcription Factor Creb in Immune Function. J. Immunol. 2010, 185, 6413–6419. [Google Scholar] [CrossRef]
- Lee, K.; Chung, Y.H.; Ahn, H.; Kim, H.; Rho, J.; Jeong, D. Selective Regulation of Mapk Signaling Mediates Rankl-Dependent Osteoclast Differentiation. Int. J. Biol. Sci. 2016, 12, 235. [Google Scholar] [CrossRef]
- Subramanian, S.; Steer, C.J. Microrna Regulation in Health and Disease. Genes 2019, 10, 457. [Google Scholar] [CrossRef]
- Iaquinta, M.R.; Lanzillotti, C.; Mazziotta, C.; Bononi, I.; Frontini, F.; Mazzoni, E.; Oton-Gonzalez, L.; Rotondo, J.C.; Torreggiani, E.; Tognon, M. The Role of Micrornas in the Osteogenic and Chondrogenic Differentiation of Mesenchymal Stem Cells and Bone Pathologies. Theranostics 2021, 11, 6573. [Google Scholar] [CrossRef]
- Luis, A.B.V.; Mariana, Y.M.B.; Erick, O.M.H.; Gabriela, G.d.L.C.; María, E.A.P.; Samik, C.; Anindya, B.; Sujay, P. The Emerging Role of Micrornas in Bone Diseases and Their Therapeutic Potential. Molecules 2021, 27, 211. [Google Scholar]
- Yu, L.; Li, W.; Yang, P.; Zhang, W.; Tao, H.; Ge, G.; Yang, H.; Bai, J.; Wang, H.; Geng, D. Osteoblastic Micrornas in Skeletal Diseases: Biological Functions and Therapeutic Implications. Eng. Regen. 2022, 3, 241–257. [Google Scholar] [CrossRef]
- Fakhry, M.; Hamade, E.; Badran, B.; Buchet, R.; Magne, D. Molecular Mechanisms of Mesenchymal Stem Cell Differentiation Towards Osteoblasts. World J. Stem Cells 2013, 5, 136. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Wang, C.; Wang, C.; Gao, Y.; Zhang, X.; Chen, X. Bmper Enhances Bone Formation by Promoting the Osteogenesis-Angiogenesis Coupling Process in Mesenchymal Stem Cells. Cell. Physiol. Biochem. 2018, 45, 1927–1939. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Jiao, G.; Liu, H.; Wu, W.; Li, S.; Wang, Q.; Xu, D.; Li, X.; Liu, H.; Chen, Y. Biological Silicon Stimulates Collagen Type 1 and Osteocalcin Synthesis in Human Osteoblast-Like Cells through the Bmp-2/Smad/Runx2 Signaling Pathway. Biol. Trace Elem. Res. 2016, 173, 306–315. [Google Scholar] [CrossRef]
- Halloran, D.; Durbano, H.W.; Nohe, A. Bone Morphogenetic Protein-2 in Development and Bone Homeostasis. J. Dev. Biol. 2020, 8, 19. [Google Scholar] [CrossRef]
- Chen, J.-R.; Lazarenko, O.P.; Blackburn, M.L.; Chen, J.F.; Randolph, C.E.; Zabaleta, J.; Schroder, K.; Pedersen, K.B.; Ronis, M.J.J. Nox4 Expression in Osteo-Progenitors Controls Bone Development in Mice During Early Life. Commun. Biol. 2022, 5, 583. [Google Scholar] [CrossRef]
- Najjar, R.S.; Knapp, D.; Wanders, D.; Feresin, R.G. Raspberry and Blackberry Act in a Synergistic Manner to Improve Cardiac Redox Proteins and Reduce Nf-Κb and Sapk/Jnk in Mice Fed a High-Fat, High-Sucrose Diet. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 1784–1796. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, N.K.; Lee, S.Y. Current understanding of RANK signaling in osteoclast differentiation and maturation. Mol. Cells 2017, 40, 706. [Google Scholar]
- Oh, J.H.; Lee, N.K. Up-regulation of RANK expression via ERK1/2 by insulin contributes to the enhancement of osteoclast differentiation. Moleucles Cells 2017, 40, 371–377. [Google Scholar]
- Kim, J.H.; Kim, N. Regulation of NFATc1 in osteoclast differentiation. J. Bone Metab. 2014, 21, 233–241. [Google Scholar] [CrossRef]
- Su, X.-D.; Yang, S.Y.; Shrestha, S.K.; Soh, Y. Aster saponin A2 inhibits osteoclastogenesis through mitogen-activated protein kinase-c-Fos-NFATc1 signaling pathway. J. Vet. Sci. 2022, 23, e47. [Google Scholar] [CrossRef]
- Chen, D.; Wang, Q.; Li, Y.; Sun, P.; Kuek, V.; Yuan, J.; Yang, J.; Wen, L.; Wang, H.; Xu, J.; et al. Notopterol attenuates estrogen deficiency-induced osteoporosis via repressing RANKL signaling and reactive oxygen species. Front. Pharmacol. 2021, 12, 664836. [Google Scholar] [CrossRef]
- Chen, D.; Ye, Z.; Wang, C.; Wang, Q.; Wang, H.; Kuek, V.; Wang, Z.; Qiu, H.; Yuan, J.; Kenny, J.; et al. Arctiin abrogates osteoclastogenesis and bone resorption via suppressing RANKL-induced ROS and NFATc1 activation. Pharmacol. Res. 2020, 159, 104944. [Google Scholar] [CrossRef] [PubMed]
- Hong, G.; Chen, Z.; Han, X.; Zhou, L.; Pang, F.; Wu, R.; Shen, Y.; He, X.; Hong, Z.; Li, Z.; et al. A novel RANKL-targeted flavonoid glycoside prevents osteoporosis through inhibiting NFATc1 and reactive oxygen species. Clin. Transl. Med. 2021, 11, e392. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Xie, N.; Hou, Y.; Chen, X.; Hu, Y.; Zhang, Y.; Meng, X.; Wang, X.; Tang, C. The enhanced mitochondrial dysfunction by cantleyoside confines inflammatory response and promotes apoptosis of human HFLS-RA cell line via AMPK/Sirt 1/NF-κB pathway activation. Biomed. Pharmacother. 2022, 149, 112847. [Google Scholar] [CrossRef] [PubMed]
- Thimmuri, D.; Shantanu, P.A.; Syamprasad, N.P.; Khan, A.; Gawali, B.; Rajdev, B.; Adhikari, C.; Ravichandiran, V.; Sharma, P.; Naidu, V.G. Hispolon inhibits RANKL induced osteoclast differentiation in vitro. Immunol. Lett. 2021, 231, 35–42. [Google Scholar] [CrossRef]
- Shan, S.; Su, J.; Zhang, Y.; Huang, X.W.; Wang, X.P.; Huang, M.C.; Li, B.; Shou, D. Benefits and mechanisms of polysaccharides from Chinese medicinal herbs for anti-osteoporosis therapy: A review. Int. J. Biol. Macromol. 2021, 193, 1996–2005. [Google Scholar]
- Springer, M.; Moco, S. Resveratrol and Its Human Metabolites—Effects on Metabolic Health and Obesity. Nutrients 2019, 11, 143. [Google Scholar] [CrossRef]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E.; Walle, U.K. High Absorption but Very Low Bioavailability of Oral Resveratrol in Humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef]
- Williams, L.D.; Burdock, G.A.; Edwards, J.A.; Beck, M.; Bausch, J. Safety Studies Conducted on High-Purity Trans-Resveratrol in Experimental Animals. Food Chem. Toxicol. 2009, 47, 2170–2182. [Google Scholar] [CrossRef]
- Asis, M.; Hemmati, N.; Moradi, S.; Venkata, K.C.N.; Mohammadi, E.; Farzaei, M.H.; Bishayee, A. Effects of Resveratrol Supplementation on Bone Biomarkers: A Systematic Review and Meta-Analysis. Ann. N. Y. Acad. Sci. 2019, 1457, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Bo, S.; Gambino, R.; Ponzo, V.; Cioffi, I.; Goitre, I.; Evangelista, A.; Ciccone, G.; Cassader, M.; Procopio, M. Effects of Resveratrol on Bone Health in Type 2 Diabetic Patients. A Double-Blind Randomized-Controlled Trial. Nutr. Diabetes 2018, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.H.X.; Zaw, J.J.T.; Xian, C.J.; Howe, P.R.C. Regular Supplementation with Resveratrol Improves Bone Mineral Density in Postmenopausal Women: A Randomized, Placebo-Controlled Trial. J. Bone Miner. Res. 2020, 35, 2121–2131. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
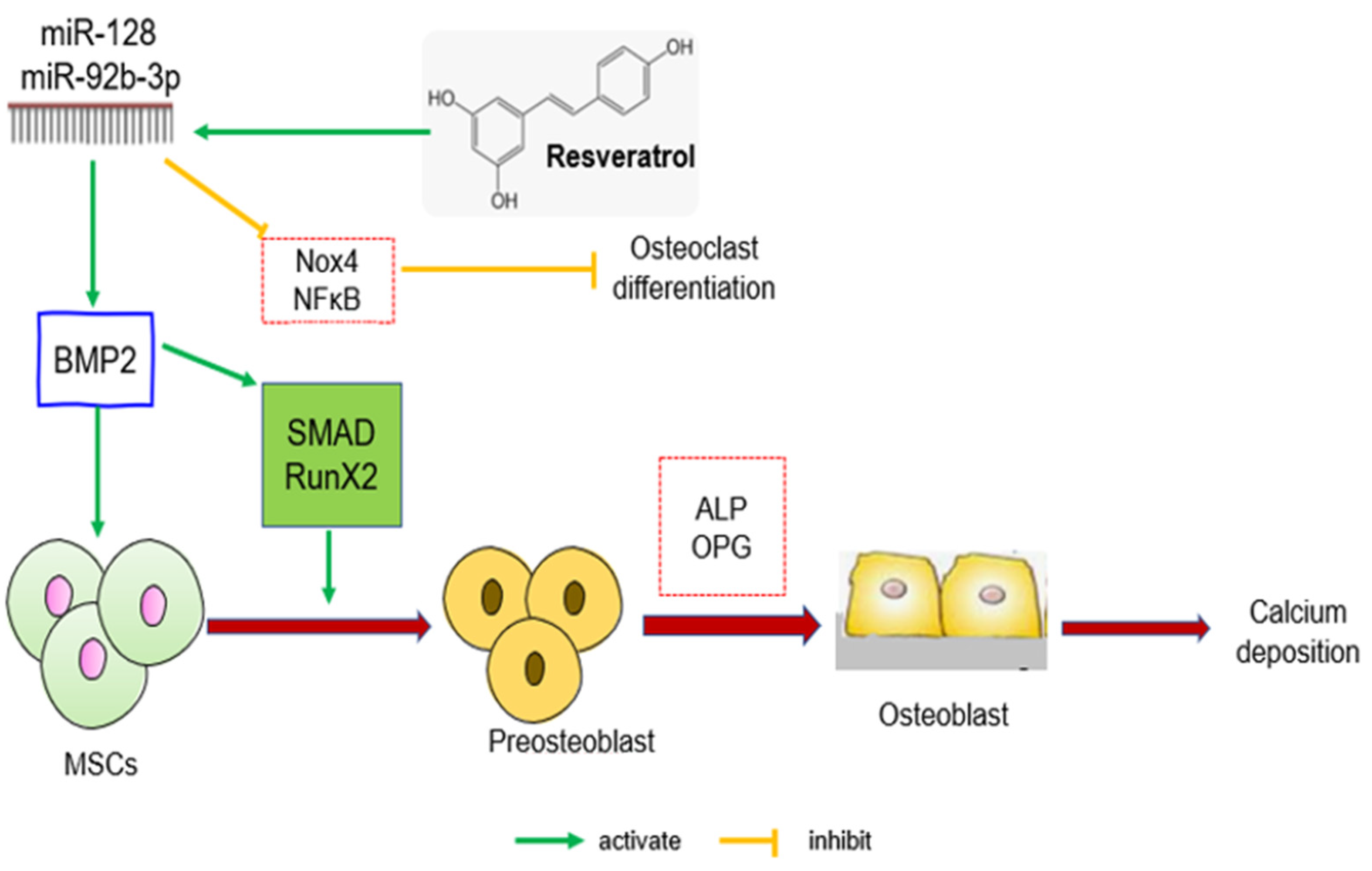{kind=link}
| Type of Animal and Induction | Intervention (Dose, Route, and Duration) | Research Findings | Mode of Action | References |
|---|---|---|---|---|
| Post menopausal osteoporosis | ||||
| Adult unmated female Sprague Dawley (SD) rats, 3-month-old, and bilateral ovariectomy | RSV (5, 15, and 45 mg/kg/day), oral, and 12 weeks | Serum estrogen ↑, serum Ca ↔, Serum P ↔, TRAP ↓, PINP ↑, ALP ↔, CTX-1 ↓, DPD ↔, and BMD ↑ | RSV increased bone mineral density by downregulating osteoclastogenesis | Zhang et al., 2020 [33] |
| Female Wistar Rats 3 to 4-month-old, bilateral ovariectomy | RSV (20, 40, 80 mg/kg/day), oral, 8 weeks | BMD ↑, Tb Ar ↑, Tb Th ↑, Tb sep ↓, Tb N ↑, Serum Ca ↔, Serum P ↔, ALP ↑, TRAP ↓, IL-6 ↓, TNFα ↓, OPG ↑, RANKL ↓ | RSV promoted osteogenesis by downregulating pro-inflammatory pathway and upregulating OPG/RANKL ratio | Zhao et al., 2014 [35] |
| Female Sprague Dawley (SD) rats, 3-month-old, and bilateral ovariectomy | RSV (625 µg/kg/day), feeding diet pellets, 4 weeks | BMD ↑, RANKL/OPG ↓, IL-23 ↓, IL-17a ↓, IL1β ↓, TNFα↓ | RSV promoted osteogenesis by downregulating pro-inflammatory pathway and RANKL/OPG ratio | Khera et al., 2019 [36] |
| Female Sprague Dawley (SD) rats, 3-month-old, bilateral ovariectomy | RSV (40 mg/kg/day), subcutaneous injection, and 10 weeks | BMD ↑ (right femur), BV/TV ↑, Tb.Th ↑, Tb.N ↑, Ct.Vol↑, Ct.Th ↑, Conn D ↑, TRAP↓, RANKL/OPG ↓, MDA ↓, ROS ↓, GSH ↑, and SOD ↑ | RSV attenuated oxidative stress damage and RSV-suppressed osteoclastogenesis | Feng et al., 2018 [37] |
| Female mice, 8-week-old, and bilateral ovariectomy | RSV (40 mg/kg/day), intraperitoneal injection, and 8 weeks | BV/TV ↑, Tb Th ↑, Tb sep ↓, ALP ↑, RunX2 ↑, Ob Nu ↑, Oc Nu ↓, TRAP ↓ Osx ↑, P1NP ↑, Sirt1 ↑, and SOD ↑ | RSV induced osteogenesis via activation of the SIRT1/FoxO1 signalling pathway. | Jiang et al. 2020 [38] |
| Female Sprague Dawley (SD) rats, 3-month-old, and bilateral ovariectomy | RSV (5, 25, 45 mg/kg/day), oral, and 8 weeks | BMD ↑, BV/TV ↑, Tb Th ↑, Tb sep ↓, ALP ↑, OPN ↑, Col1a1↑, Sirt1 ↑, and NF-κB ↓ | RSV increased osteoblast differentiation by activating SIRT1 and inhibiting the NF-kB signalling pathway. | Feng et al., 2014 [39] |
| Female albino rats, 3-month-old, and bilateral ovariectomy | RSV (80 mg/kg/day), oral, and 8 weeks | BMC ↑, BMD ↑, PINP ↓, ALP ↓, CTX-1 ↓, RANKL/OPG ↓, Sirt1 ↑, FOXO3a ↑, Wnt ↑, β catenin ↑, and RunX2 ↑ | RSV-promoted osteogenesis through Sirt1 and Wnt signalling pathways. | Elseweidy et al., 2021 [42] |
| Sprague Dawley (SD) rats, 6-month-old, and bilateral ovariectomy | RSV (50, 100, 200 mg/kg/day), oral, and 12 weeks | BMD ↑, Tb Ar ↑, Tb Th ↑, Tb sep ↓, Tb N ↑, miR-92b-3p ↑, NF-κB ↓, Nox4 ↓, CtsK ↓, Smad7 ↑, BMP2 ↑, RUNX-2 ↑, ALP ↑, OPG ↑, TRAP ↓, CTX-1 ↓, Ca ↑, and P ↑ | RSV increased miR-92b-3p transcriptional activity to enhance the BMP-2/Smad/Runx2 signalling and reduced NF-κB to promote osteogenesis. | Zhang et al., 2020 [45] |
| Female Wistar rats, 8 –week old, and bilateral ovariectomy | RSV (500 mg/kg/day), tail intravenous injection, and 1 day | BMD ↑, ALP ↑, Ca ↑, miR3383p ↓, RunX2 ↑ | RSV decreased miR 338 3p expression, which was followed by a rise in RunX2 expression. | Guo et al., 2015 [46] |
| Female Sprague Dawley (SD) rats, 10 to 12-week-old, bilateral ovariectomy | RSV (10, 20, and 40 mg/kg/day), oral, 8 weeks | BMD ↑ (lumbar L3, femur-tibia), Ca content ↑, VEGF ↑, Col1a1 ↑, RANKL ↓, autophagy related genes, atg 5, atg 7, and atg 12 ↑ | RSV promoted osteoblastic differentiation and suppressed osteoclastic differentiation by regulating autophagy | Wang et al., 2020 [50] |
| Senile osteoporosis | ||||
| Aged male Wistar rats and 22 –month old | RSV (10 mg/kg/day), oral, and 10 weeks | BV/TV ↑, Tb Th ↑, Tb sep ↓, Cr Th ↑, CTX ↔, and OCN ↔ | Tresguerres et al., 2014 [53] | |
| Aged male Wistar albino rats and 18 to 20 –week old | RSV (20 mg/kg/day), oral, and 6 weeks | BMD ↔, OPG ↑, RANKL ↓, FoxO1 ↑, Sirt1 ↑, GSH ↑, MDA ↓, hsCRP ↓, TNFα↓, IL6 ↓, and IL1β ↓ | RSV showed anti-osteoporotic effects with involvement of FoxO1/SIRT/RANK/OPG pathways | Ameen et al., 2020 [54] |
| Aged male Wistar Rats and 6 –month old | RSV (20 mg/kg/day), oral, and 3 months | BV/TV ↔, cortical bone volume ↔, Sirt1 ↔, Osx ↔, OCN ↑, CTX-1 ↓, and ALP ↔, | Lee et al., 2014 [55] | |
| Senile and diuse osteoporosis | ||||
| Aged male Fischer 344 9 Brown Norway, 33-month-old, and hindlimbs suspended for 14 days | RSV (12.5 mg/kg/day), oral, and 3 weeks | BV/TV ↔, Tb Th ↑, Tb sep ↓, ALP ↑, OCN ↑, C-reactive protein ↓, TNFα↓, Ca ↔, P ↔, TRAP ↔, and CTX1 ↔ | RSV increased osteoblast bone formation and possibly due to reduced inflammation. | Durbin et al., 2014 [58] |
| Secondary osteoporosis | ||||
| Male Sprague Dawley (SD) rats, 3-month-old, intramuscular injection of dexamethasone 5 mg/kg, and twice a week 6 weeks | RSV (5, 45 mg/kg/day), oral, and 8 weeks | BMD ↑ (right femur), femoral porosity ↓, ALP ↓, OCN ↓, Sirt1 ↑, LC3 ↑, Beclin-1 ↑, phospho-mTOR ↓, and phospho-Akt ↓ | RSVenhanced SIRT1 expression and protect dexamethasone-treated osteoblasts by autophagy via Akt/mTOR pathway | Yang 2019 [60] |
| Male Sprague Dawley (SD) rats, 6 week old, and subcutaneous injection of methotrexate for five days (once daily) 0.75 mg/kg, | RSV (1 mg/kg/day), oral, and 12 days (7 days pretreatment, and 5 days during methotrexate injection) | growth plate thickness ↑, primary spongiosa bone, BV/TV ↑, Tb Th ↑, adipose density ↓, RunX2 ↔, Osx ↓, Ocn ↔, TRAP ↓, TNF-α ↓, IL-1 ↓, and IL-6 ↓ | RSV improved bone microstructure by inhibiting osteoclastogenesis and inflammatory processes. | Lee 2017 [62] |
| Type of Cell and Induction | Intervention | Research Findings | Mode of Action | References |
|---|---|---|---|---|
| Bone forming cells | ||||
| 3-D osteogenic differentiation on collagen scaffolds of rat adipose stem cells | 25 µM RSV | ALP ↑, OCN ↑, OPG ↑, and Mineral density ↑ | Dosier et al., 2012 [64] | |
| HBMSC cells | 10−6 M RSV | Cell proliferation ↑, Calcium deposition ↑, ALP ↑, Runx2 ↑, Osterix ↑, OCN ↑, ERK 1/2↑, ER ↑, and MAPK ↓ | RSV induced HBMSC proliferation and differentiation through an ER-dependent pathway linked to ERK1/2 activation. | Dai et al., 2007 [65] |
| BMSC cells | 20 µM RSV | Cell proliferation ↑, Calcium deposition ↑, MMP ↔, ALP ↑, Runx2 ↑, ONN ↑, OCN ↑, GSK-3b ↑, β catenin ↑, ERK ↑, and MAPK ↑ | RSV promoted osteoblastic differentiation of BMSCs by activating the Wnt/β-catenin and ERK/MAPK signalling pathways. | Zhao et al., 2018 [66] |
| Senescent BMSC cells | 5,10, 15, 20, and 25 µM RSV | Cell viability ↑, ALP ↑, Col-I ↑, OCN ↑, OPN ↑, RunX2 ↑, ROS ↓, p16 ↓, p21 ↓, p53 ↓, and AMPK ↑ | RSV promoted osteogenic differentiation of senescent BMSCs by slowing cell ageing and decreasing ROS production via AMPK activation. | Zhou et al., 2019 [70] |
| HBMSC cells, hypoxia | 0.1 and 1 µM RSV | Cell viability ↑, OCN ↑, OPN ↑, ALP ↑, RunX2 ↑, and ROS ↓ | RSV reduced high-altitude hypoxia-induced osteoporosis by increasing osteoblastogenesis and inhibiting the ROS/HIF-1 signalling pathway. | Yan et al., 2022 [72] |
| MC3T3-E1 cells, induced with 120 µM H2O2 | 15 µM RSV | p53 ↓, Bax ↓, Bcl-2 ↓, and caspase 9 ↓ | RSV suppressed oxidative stress-induced apoptosis in osteoblasts. | He et al., 2015 [73] |
| MC3T3-E1 cells, induced with 5 µM cadmium | 10 µM RSV | Cell viability ↑, ALP ↑, Col 1 ↑, BMP2 ↑, RunX2 ↑, ERK ↑, and JNK ↑ | RSV protected MC3T3-E1 cells from cadmium-induced osteogenic differentiation suppression by modulating the ERK1/2 and JNK pathways. | Mei et al., 2021 [74] |
| human periosteum derived MSCs (PO-MScs) | 5 µM RSV | Cell proliferation ↑, Calcium deposition ↑, ALP ↑, mitochondrial mass ↑, and mitochondrial DNA copy number ↑ | RSV stimulated mitochondrial biogenesis in the process of PO-MSC osteogenic differentiation. | Moon et al., 2020 [79] |
| MC3T3-E1 cells,0.5, 1, and 2 μg/mL Lipopolysaccharides(LPS)-Induced Inhibition of Osteoblast | 25 µM RSV | Cell viability ↑, ALP ↑, OCN ↑, OPN ↑, RUNX2 ↑, Sirt1 ↑, and PGC-1α ↑ | RSV reduced the inhibition of LPS on osteoblast development via enhancing mitochondrial activity and SIRT1 pathway | Ma et al., 2022 [87] |
| HBMSC cells, 1 μg/mL Lipopolysaccharides | 25 µM RSV | Cell proliferation ↑, ALP ↑, PINP ↑, OPG ↑, LDH ↔, IL-6 ↔, and IL-8 ↔ | RSV promoted osteoblast differentiation independently of inflammation. | Ornstrup et al., 2015 [88] |
| HOB cells, 5 µM alendronate and zoledronate | 10 µM RSV | Cell viability ↑, BMP2↑, OPG ↑, Col1 ↑, Sirt1 ↑, and Calcium deposition (Alizarin Red) ↑ | RSV improved the osteoblast proliferation, differentiation and mineralization that treated with alendronate and zoledronate, through BMP2, and SIRT1 signalling pathway | Borsani et al., 2018 [91] |
| MC3T3-E1 cells, induced with 10 µM PGE2 | 50 µM RSV | OPG ↓, p44/p42 MAP kinase ↓, p38 MAP kinase ↓, SAPK/JNK ↓, and Sirt1 ↔ | Resveratrol inhibited PGE2-stimulated OPG synthesis in osteoblasts by inhibiting p44/p42 MAP kinase, p38 MAP kinase, and SAPK/JNK, and these suppressive effects are independent of SIRT1 activation. | Yamamoto et al., 2014 [98] |
| MC3T3-E1 cells, induced with 10 µM PGF2α | 10 µM RSV | OPG ↓, p44/p42 MAP kinase ↓, p38 MAP kinase, SAPK/JNK↓, and SIRT1 ↑ | RSV suppressed PGF2-stimulated OPG synthesis in osteoblasts via the MAP kinase pathways and these suppressions are mediated by SIRT1 | Kuroyanagi et al., 2014 [99] |
| Primary rat calvarial osteoblasts | 0.05, 0.1 μM RSV | Cell viability ↑, Calcium deposition ↑, ALP ↑, ERα ↑, and ERβ ↔ | RSV stimulated osteoblast differentiation via an oestrogen-dependent pathway | Shah et al., 2022 [102] |
| human embryonic MSCs | 5 µM RSV | RunX2 ↑, ALP ↑, Ocn ↑, SIRT1 ↑, FoxO3a ↑, and SIRT1-FoxO3a binds to FoxO response element (FRE) site | RSV promoted osteogenesis of human mesenchymal stem cells by upregulating RUNX2 gene expression via the SIRT1/FoxO3a axis. | Tseng et al., 2011 [103] |
| Canin MSCs | 5 µM RSV | Col1 ↑, Ocn ↑, β1- Integrin ↑, RunX2 ↑, SIRT1 ↑, TNFβ ↓, and NF-κB ↓ | RSV suppressed TNFβ expression by activation of SIRT1 and inhibition of NFκB signalling pathway. | Costanze et al., 2020 [104] |
| Bone resorbing cells | ||||
| RAW 264.7 cells, induced with 5 µg/mL LPS | 1, 5, 10 µM RSV | Cytotoxic effect ↔, Nitric oxide ↓, PGE2 ↓, iNOS ↓, COX-2 ↓, TNF-α ↓, IL-1β ↓, CREB ↓, MAPK ↓, PI3K/AKT ↓, and Sirt1 ↑ | RSV inhibited proinflammatory mediators and cytokines production in response to LPS by activation SIRT1 expression, and inhibition of PI3K/AKT, and CREB and MAPK signalling pathway | Zong et al., 2012 [105] |
| RAW 264.7 cells, induced with 10-4M H2O2 | 10-5M RSV | Cytotoxic effect ↔, MMP-9 ↓, TRAP ↓, CtsK ↓, MDA ↓, ROS ↓, SOD ↑ GSH-PX ↑, FoxO1 ↑, and PI3K/AKT ↓ | RSV increased FoxO1 transcriptional activity by inhibiting the PI3K/AKT signalling pathway, promoting oxidative stress resistance, and inhibiting osteoclastogenesis. | Feng et al., 2018 [37] |
| RAW 264.7 cells, 0.1 µM doxorubicin-Induced Osteoclast Differentiation | 10 µM RSV | Cytotoxic effect ↔, Oc-Stamp ↓, RANK ↓, TRAP ↓, CtsK ↓ NFATc1 ↓, FoxO1 ↑, SOD 1 ↑, and Nrf 2 ↑ | RSV prevented doxorubicin-induced osteoclast fusion and activation and increased FoxO1 transcriptional activity by inhibiting the NFATC1 signalling pathway | Poudel et al., 2022 [106] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad Hairi, H.; Jayusman, P.A.; Shuid, A.N. Revisiting Resveratrol as an Osteoprotective Agent: Molecular Evidence from In Vivo and In Vitro Studies. Biomedicines 2023, 11, 1453. https://doi.org/10.3390/biomedicines11051453
Ahmad Hairi H, Jayusman PA, Shuid AN. Revisiting Resveratrol as an Osteoprotective Agent: Molecular Evidence from In Vivo and In Vitro Studies. Biomedicines. 2023; 11(5):1453. https://doi.org/10.3390/biomedicines11051453
Chicago/Turabian StyleAhmad Hairi, Haryati, Putri Ayu Jayusman, and Ahmad Nazrun Shuid. 2023. "Revisiting Resveratrol as an Osteoprotective Agent: Molecular Evidence from In Vivo and In Vitro Studies" Biomedicines 11, no. 5: 1453. https://doi.org/10.3390/biomedicines11051453