
The Imitation of the Ovarian Fatty Acid Profile of Superfertile Dummerstorf Mouse Lines during IVM of Control Line Oocytes Could Influence Their Maturation Rates
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. IVM (In Vitro Maturation)
1.2. Superfertile Mouse Lines
2. Results
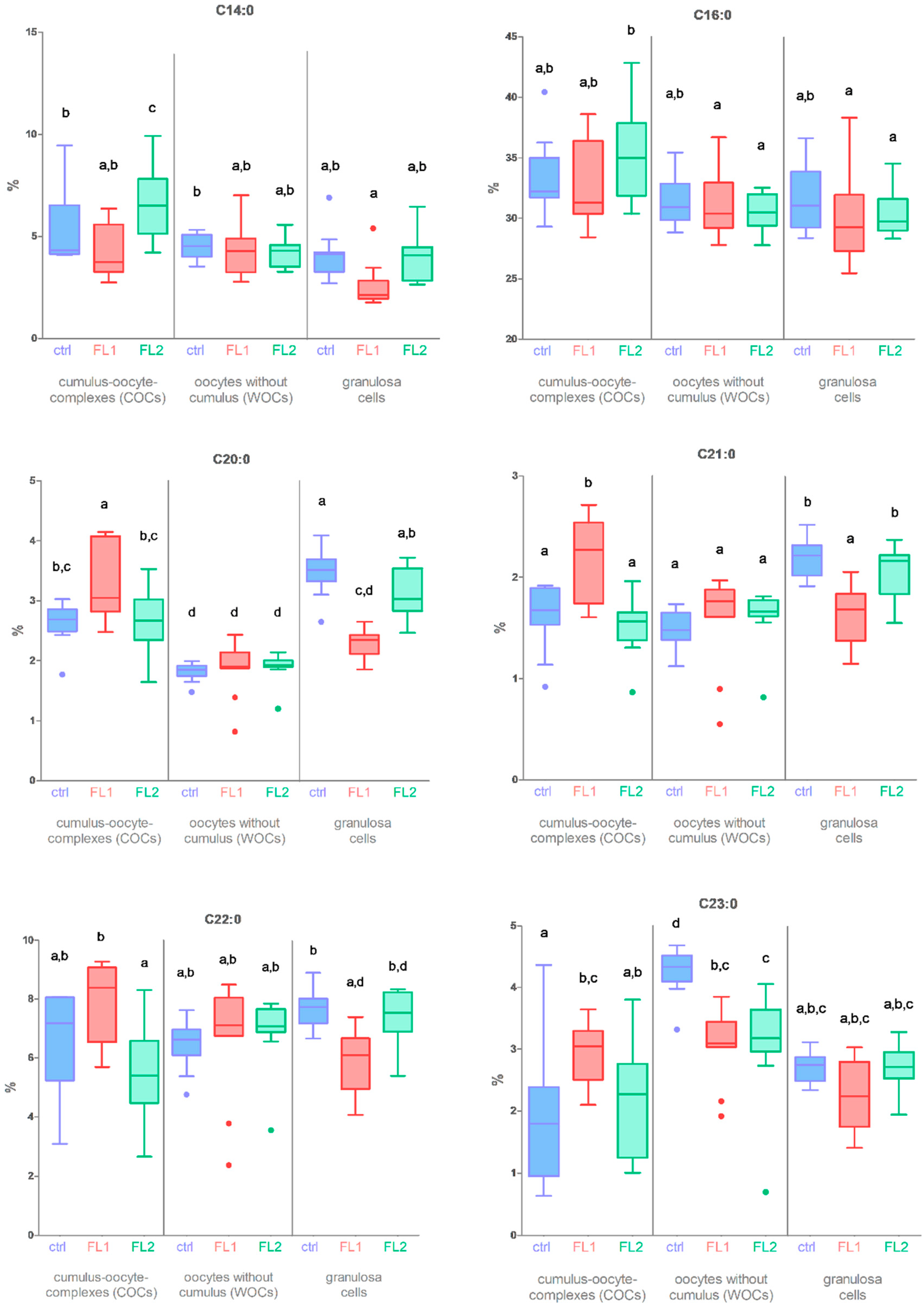
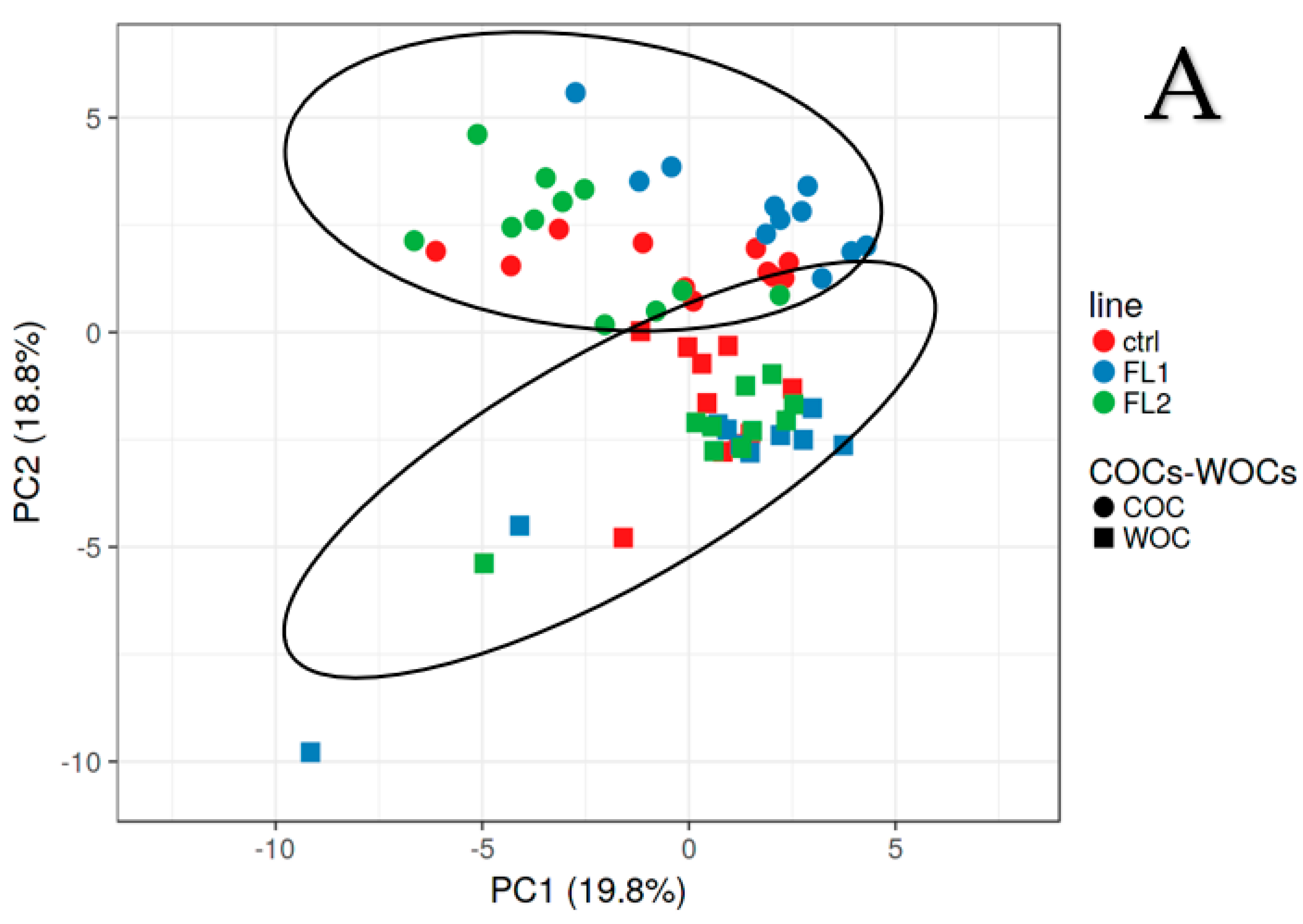
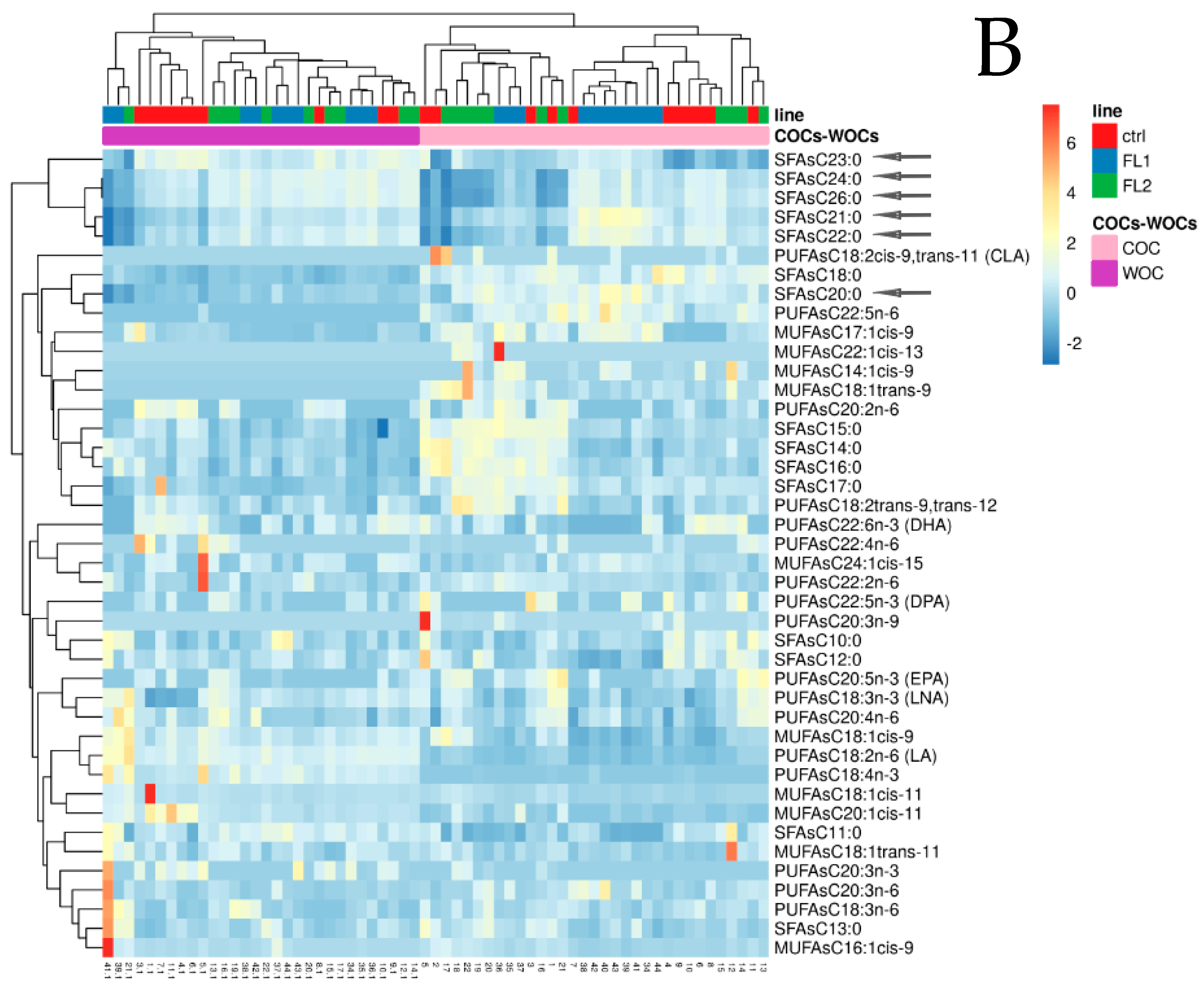
2.1. Fatty Acid Concentration in COCs-WOCs-Granulosa Cells
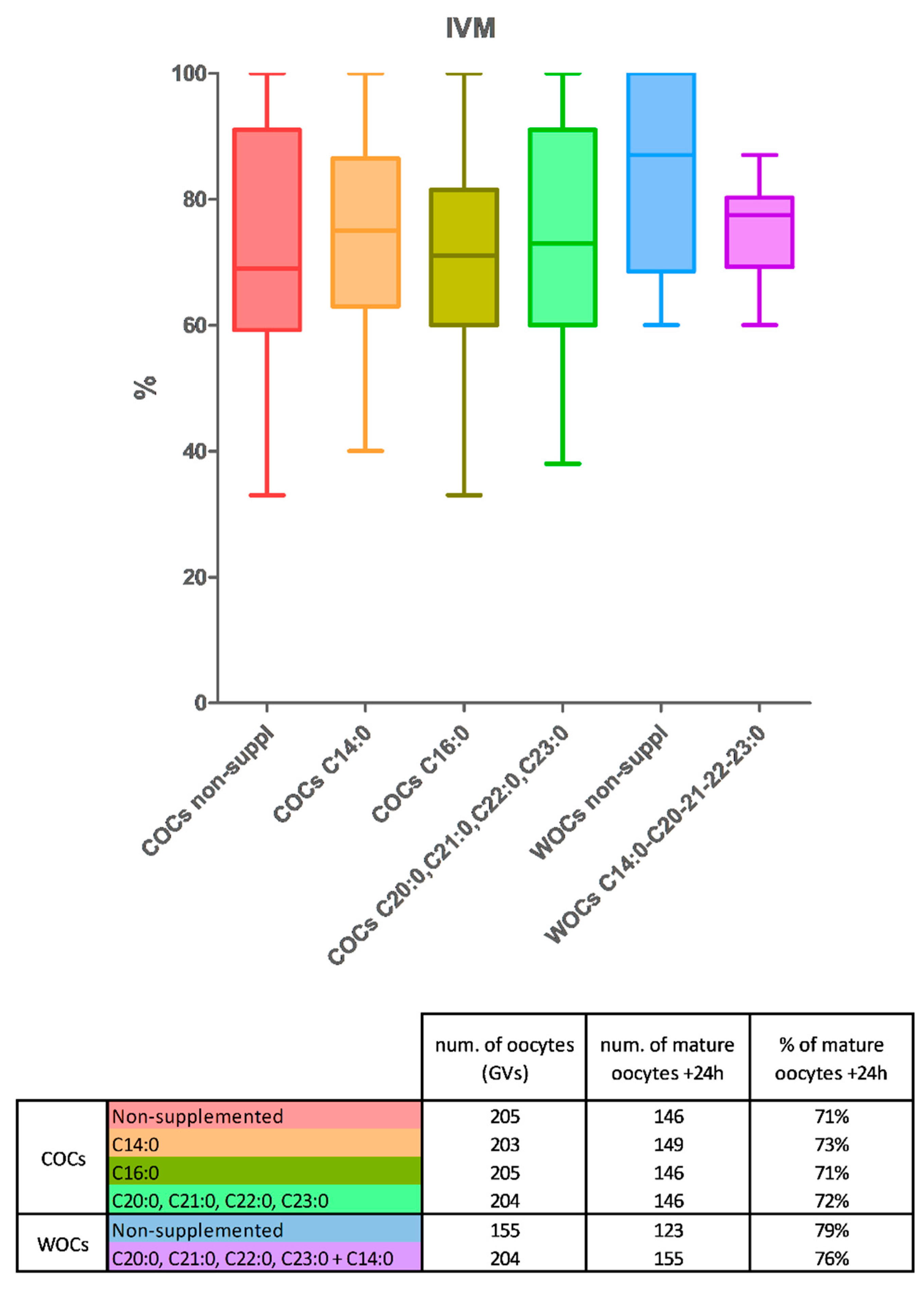
2.2. Maturation Rates
3. Discussion
4. Materials and Methods
4.1. Fatty Acids and Media Preparation
4.2. Mice
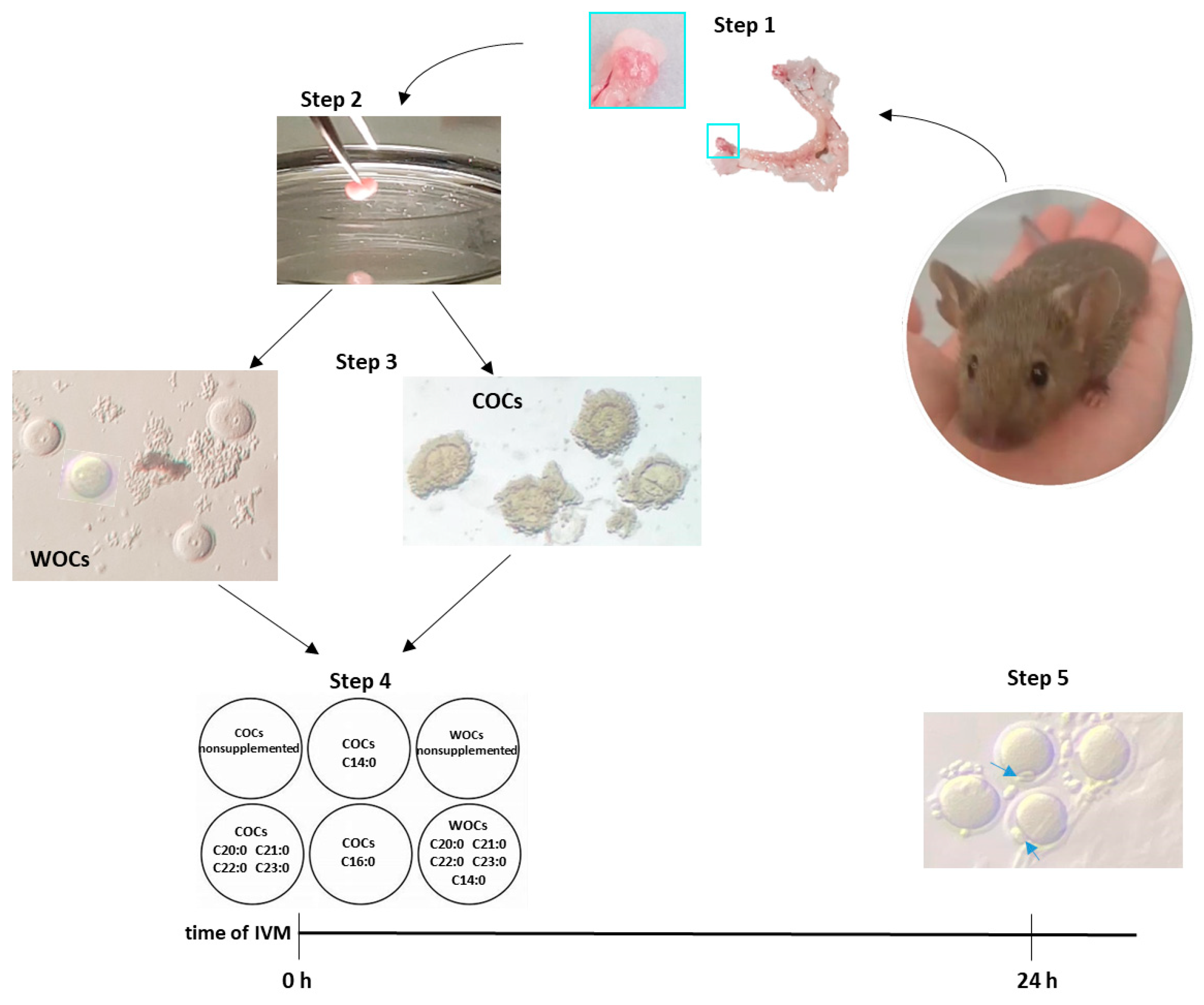
4.3. Sample Collection and In Vitro Maturation (IVM)
- n. 205 COCs—nonsupplemented, ctrl
- n. 203 COCs—with tetradecanoic acid (C 14:0)
- n. 205 COCs—with palmitic acid (C 16:0)
- n. 204 COCs—with a mixture of eicosanoic acid (C 20:0), henicosanoic acid (C 21:0), docosanoic acid (C 22:0), and tricosanoic acid (C23:0) in equal quantities (12.5 µM each).
- n. 155 WOCs—nonsupplemented
- n. 204 WOCs—with a mixture of tetradecanoic acid (C 14:0, 25 µM), eicosanoic acid (C 20:0, 6.25 µM), henicosanoic acid (C 21:0, 6.25 µM), docosanoic acid (C 22:0, 6.25 µM), and tricosanoic acid (C23:0, 6.25 µM).
4.4. Assessment of Nuclear Maturation
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aitken, R.J. The changing tide of human fertility. Hum. Reprod. 2022, 37, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Telfer, E.E.; Andersen, C.Y. In vitro growth and maturation of primordial follicles and immature oocytes. Fertil. Steril. 2021, 115, 1116–1125. [Google Scholar] [CrossRef]
- Xiao, S.; Duncan, F.E.; Bai, L.; Nguyen, C.T.; Shea, L.D.; Woodruff, T.K. Size-specific follicle selection improves mouse oocyte reproductive outcomes. Reproduction 2015, 150, 183–192. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.J.; Pendola, J.K.; Eppig, J.J. A revised protocol for in vitro development of mouse oocytes from primordial follicles dramatically improves their developmental competence. Biol. Reprod. 2003, 68, 1682–1686. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhong, Z.; Latham, K.E. Strain-specific spontaneous activation during mouse oocyte maturation. Fertil. Steril. 2012, 98, 200–206. [Google Scholar] [CrossRef]
- Hardy, K.; Wright, C.S.; Franks, S.; Winston, R.M.L. In Vitro maturation of oocytes. Br. Med. Bull. 2000, 56, 588–602. [Google Scholar] [CrossRef] [PubMed]
- Schüler, L.; Bünger, L. The Reproductive Lifetime Performance of Laboratory Mouse Lines Selected for Fertility. Arch. Anim. Breed. 1982, 25, 275–281. [Google Scholar]
- Langhammer, M.; Wytrwat, E.; Michaelis, M.; Schön, J.; Tuchscherer, A.; Reinsch, N.; Weitzel, J.M. Two mouse lines selected for large litter size display different lifetime fecundities. Reproduction 2021, 161, 721–730. [Google Scholar] [CrossRef]
- Edson, M.A.; Lin, Y.N.; Matzuk, M.M. Deletion of the novel oocyte-enriched gene, Gpr149, leads to increased fertility in mice. Endocrinology 2010, 151, 358–368. [Google Scholar] [CrossRef]
- Hsu, S.Y.; Lai, R.J.; Finegold, M.; Hsueh, A.J. Targeted overexpression of Bcl-2 in ovaries of transgenic mice leads to decreased follicle apoptosis, enhanced folliculogenesis. Endocrinology 1996, 137, 4837–4843. [Google Scholar] [CrossRef]
- Calanni-Pileri, M.; Weitzel, J.M.; Langhammer, M.; Michaelis, M. Higher quality rather than superior quantity of oocytes determine the amount of fertilizable oocytes in two outbred Dummerstorf high-fertility mouse lines. Reprod. Domest. Anim. 2022, 57, 1198–1207. [Google Scholar] [CrossRef] [PubMed]
- Calanni-Pileri, M.; Weitzel, J.M.; Dannenberger, D.; Langhammer, M.; Michaelis, M. Lower Plasmatic Levels of Saturated Fatty Acids and a Characteristic Fatty Acid Composition in the Ovary Could Contribute to the High-Fertility Phenotype in Dummerstorf Superfertile Mice. Int. J. Mol. Sci. 2022, 23, 10245. [Google Scholar] [CrossRef] [PubMed]
- Lolicato, F.; Brouwers, J.F.; de Lest, C.H.V.; Wubbolts, R.; Aardema, H.; Priore, P.; Gadella, B.M. The cumulus cell layer protects the bovine maturing oocyte against fatty acid-induced lipotoxicity. Biol. Reprod. 2015, 92, 16. [Google Scholar] [CrossRef] [PubMed]
- Aardema, H.; Lolicato, F.; van de Lest, C.H.; Brouwers, J.F.; Vaandrager, A.B.; van Tol, H.T.; Gadella, B.M. Bovine Cumulus Cells Protect Maturing Oocytes from Increased Fatty Acid Levels by Massive Intracellular Lipid Storage. Biol. Reprod. 2013, 88, 164. [Google Scholar] [CrossRef] [PubMed]
- Aardema, H.; Vos, P.L.; Lolicato, F.; Roelen, B.A.; Knijn, H.M.; Vaandrager, A.B.; Gadella, B.M. Oleic Acid Prevents Detrimental Effects of Saturated Fatty Acids on Bovine Oocyte Developmental Competence1. Biol. Reprod. 2011, 85, 62–69. [Google Scholar] [CrossRef]
- Baddela, V.S.; Sharma, A.; Plinski, C.; Vanselow, J. Palmitic acid protects granulosa cells from oleic acid induced steatosis and rescues progesterone production via cAMP dependent mechanism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2022, 1867, 159159. [Google Scholar] [CrossRef]
- Calanni-Pileri, M.; Weitzel, J.M.; Langhammer, M.; Wytrwat, E.; Michaelis, M. Altered insulin; leptin, and ghrelin hormone levels and atypical estrous cycle lengths in two highly-fertile mouse lines. Reprod. Domest. Anim. 2022, 57, 577–586. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Anderson, R.A.; Mitchell, R.T.; Kelsey, T.W.; Spears, N.; Telfer, E.E.; Wallace, W.H. Cancer treatment and gonadal function: Experimental and established strategies for fertility preservation in children and young adults. Lancet Diabetes Endocrinol. 2015, 3, 556–567. [Google Scholar] [CrossRef]
- Anderson, R.A.; Telfer, E.E. Being a good egg in the 21st century. Br. Med. Bull. 2018, 127, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Telfer, E.E. Progress and prospects for developing human immature oocytes in vitro. Reproduction 2019, 158, F45–F54. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.H.; Aitken, R.J.; Bromfield, E.G.; Nixon, B. DNA damage and repair in the female germline: Contributions to ART. Hum. Reprod. Update 2019, 25, 180–201. [Google Scholar] [CrossRef] [PubMed]
- Hirao, Y.; Nagai, T.; Kubo, M.; Miyano, T.; Miyake, M.; Kato, S. In vitro growth and maturation of pig oocytes. J. Reprod. Fertil. 1994, 100, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Crozet, N.; Dahirel, M.; Gall, L. Meiotic competence of in vitro grown goat oocytes. J. Reprod. Fertil. 2000, 118, 367–373. [Google Scholar] [CrossRef]
- Hirao, Y.; Naruse, K.; Kaneda, M.; Somfai, T.; Iga, K.; Shimizu, M.; Takenouchi, N. Production of fertile offspring from oocytes grown in vitro by nuclear transfer in cattle. Biol. Reprod. 2013, 89, 57. [Google Scholar] [CrossRef]
- Dieci, C.; Lodde, V.; Labreque, R.; Dufort, I.; Tessaro, I.; Sirard, M.A.; Luciano, A.M. Differences in cumulus cell gene expression indicate the benefit of a pre-maturation step to improve in-vitro bovine embryo production. Mol. Hum. Reprod. 2016, 22, 882–897. [Google Scholar] [CrossRef]
- Chian, R.C.; Uzelac, P.S.; Nargund, G. In vitro maturation of human immature oocytes for fertility preservation. Fertil. Steril. 2013, 99, 1173–1181. [Google Scholar] [CrossRef]
- Assareh, N.; Shahemabadi, M.; Varnosfaderani, S.R.; Jafarpour, F.; Hajian, M.; Nasr-Esfahani, M.H. Sequential IVM by CNP preincubation and cooperating of PGE2 with AREG enhances developmental competence of SCNT reconstructs in goat. Sci. Rep. 2022, 12, 4243. [Google Scholar] [CrossRef]
- Langhammer, M.; Michaelis, M.; Hoeflich, A.; Sobczak, A.; Schoen, J.; Weitzel, J.M. High-fertility phenotypes: Two outbred mouse models exhibit substantially different molecular and physiological strategies warranting improved fertility. Reproduction 2014, 147, 427–433. [Google Scholar] [CrossRef]
- Langhammer, M.; Michaelis, M.; Hartmann, M.F.; Wudy, S.A.; Sobczak, A.; Nurnberg, G.; Weitzel, J.M. Reproductive performance primarily depends on the female genotype in a two-factorial breeding experiment using high-fertility mouse lines. Reproduction 2017, 153, 361–368. [Google Scholar] [CrossRef]
- Aizawa, R.; Ibayashi, M.; Tatsumi, T.; Yamamoto, A.; Kokubo, T.; Miyasaka, N.; Tsukamoto, S. Synthesis and maintenance of lipid droplets are essential for mouse preimplantation embryonic development. Development 2019, 146, 1767–1776. [Google Scholar] [CrossRef] [PubMed]
- Leroy, J.L.; Vanholder, T.; Mateusen, B.; Christophe, A.; Opsomer, G.; de Kruif, A.; Van Soom, A. Non-esterified fatty acids in follicular fluid of dairy cows and their effect on developmental capacity of bovine oocytes in vitro. Reproduction 2005, 130, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.-M.; Yanase, T.; Nishi, Y.; Tanaka, A.; Saito, M.; Jin, C.H.; Nawata, H. Saturated FFAs, Palmitic Acid and Stearic Acid, Induce Apoptosis in Human Granulosa Cells. Endocrinology 2001, 142, 3590–3597. [Google Scholar] [CrossRef] [PubMed]
- Kalita, K.; Deka, B.C.; Biswas, R.K.; Barua, P.M.; Borah, P.; Dutta, D.J.; Das, S.K. Effect of Different Types of In Vitro Maturation Medium (IVM) on Cumulus Cell Expansion and Nuclear Maturation Rate of Non-vitrified and Post Vitrified-Thawed Porcine Follicular Oocytes. J. Fertil. In Vitro IVF Worldw. Reprod. Med. Genet. Stem Cell Biol. 2019, 6, 211. [Google Scholar] [CrossRef]
- Singh, S.R.; Byers, S.L.; Wiles, M.V.; Dunn, S.L.; Taft, R.A. Mouse Estrous Cycle Identification Tool and Images. PLoS ONE 2012, 7, e35538. [Google Scholar] [CrossRef]
- Behringer, M.G.R.; Nagy, K.V.; Nagy, A. Manipulating the Mouse Embryo: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2014. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calanni-Pileri, M.; Michaelis, M.; Langhammer, M.; Tognetti, P.R.; Weitzel, J.M. The Imitation of the Ovarian Fatty Acid Profile of Superfertile Dummerstorf Mouse Lines during IVM of Control Line Oocytes Could Influence Their Maturation Rates. Biomedicines 2023, 11, 1439. https://doi.org/10.3390/biomedicines11051439
Calanni-Pileri M, Michaelis M, Langhammer M, Tognetti PR, Weitzel JM. The Imitation of the Ovarian Fatty Acid Profile of Superfertile Dummerstorf Mouse Lines during IVM of Control Line Oocytes Could Influence Their Maturation Rates. Biomedicines. 2023; 11(5):1439. https://doi.org/10.3390/biomedicines11051439
Chicago/Turabian StyleCalanni-Pileri, Michela, Marten Michaelis, Martina Langhammer, Paolo Rosellini Tognetti, and Joachim M. Weitzel. 2023. "The Imitation of the Ovarian Fatty Acid Profile of Superfertile Dummerstorf Mouse Lines during IVM of Control Line Oocytes Could Influence Their Maturation Rates" Biomedicines 11, no. 5: 1439. https://doi.org/10.3390/biomedicines11051439