
Umbilical-Cord-Derived Mesenchymal Stromal Cells Modulate 26 Out of 41 T Cell Subsets from Systemic Sclerosis Patients

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. GMP Manufacturing of SLCTmsc02–Umbilical Cord Tissue MSCs
2.3. Peripheral Blood Mononuclear Cell Isolation
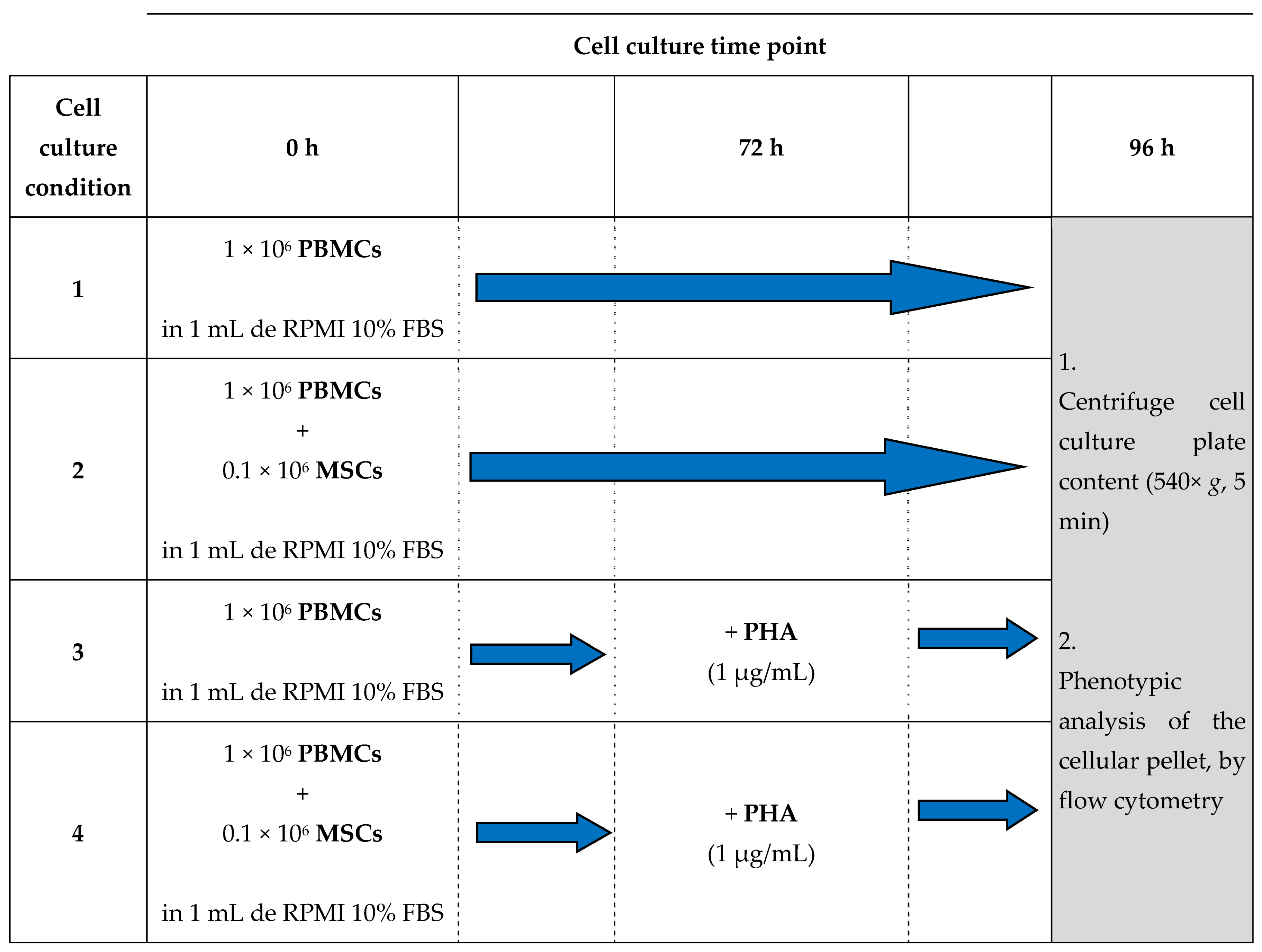
2.4. Cell Culture
2.5. Flow Cytometry Analyses
2.5.1. Staining Protocol and Sample Acquisition
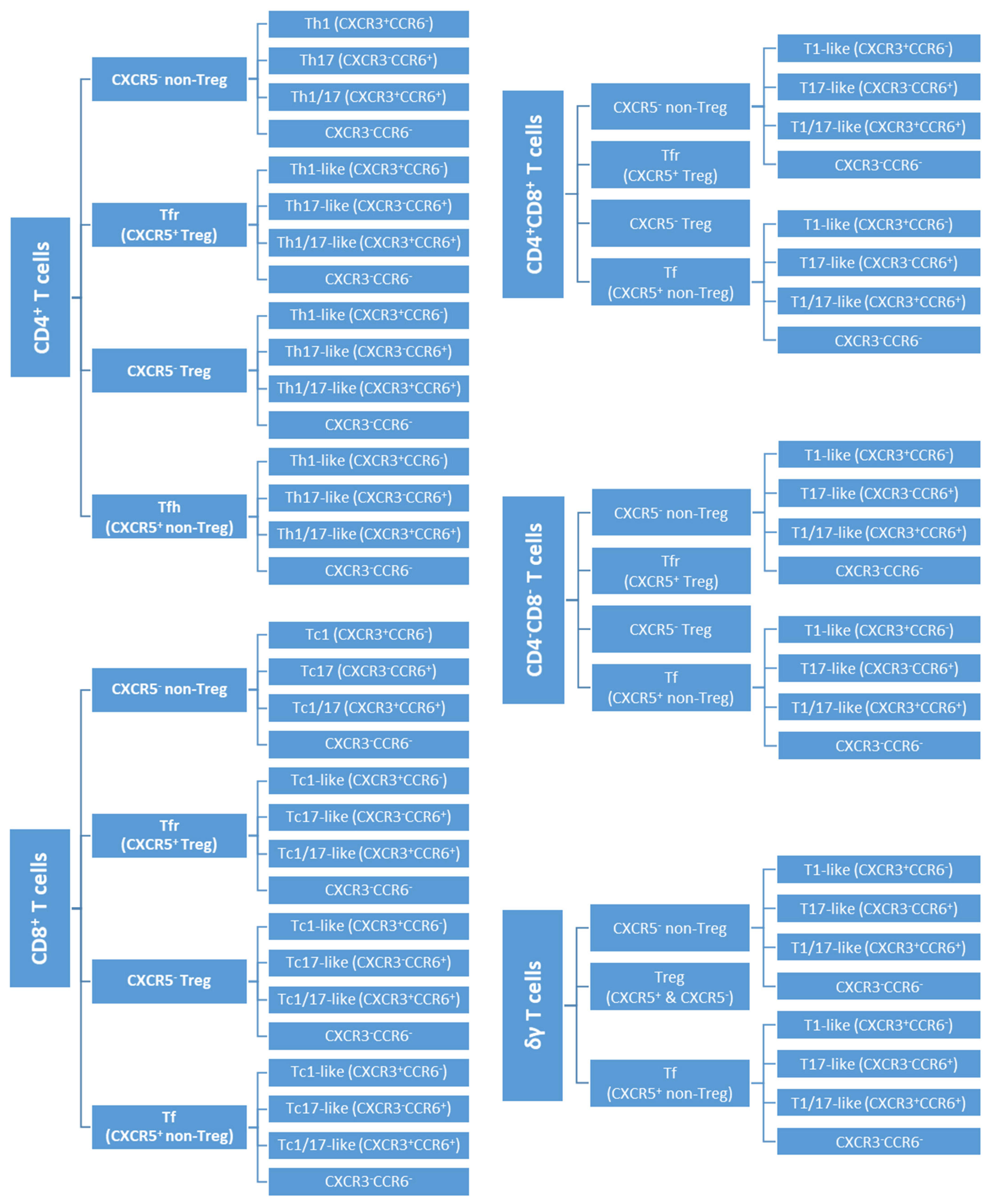
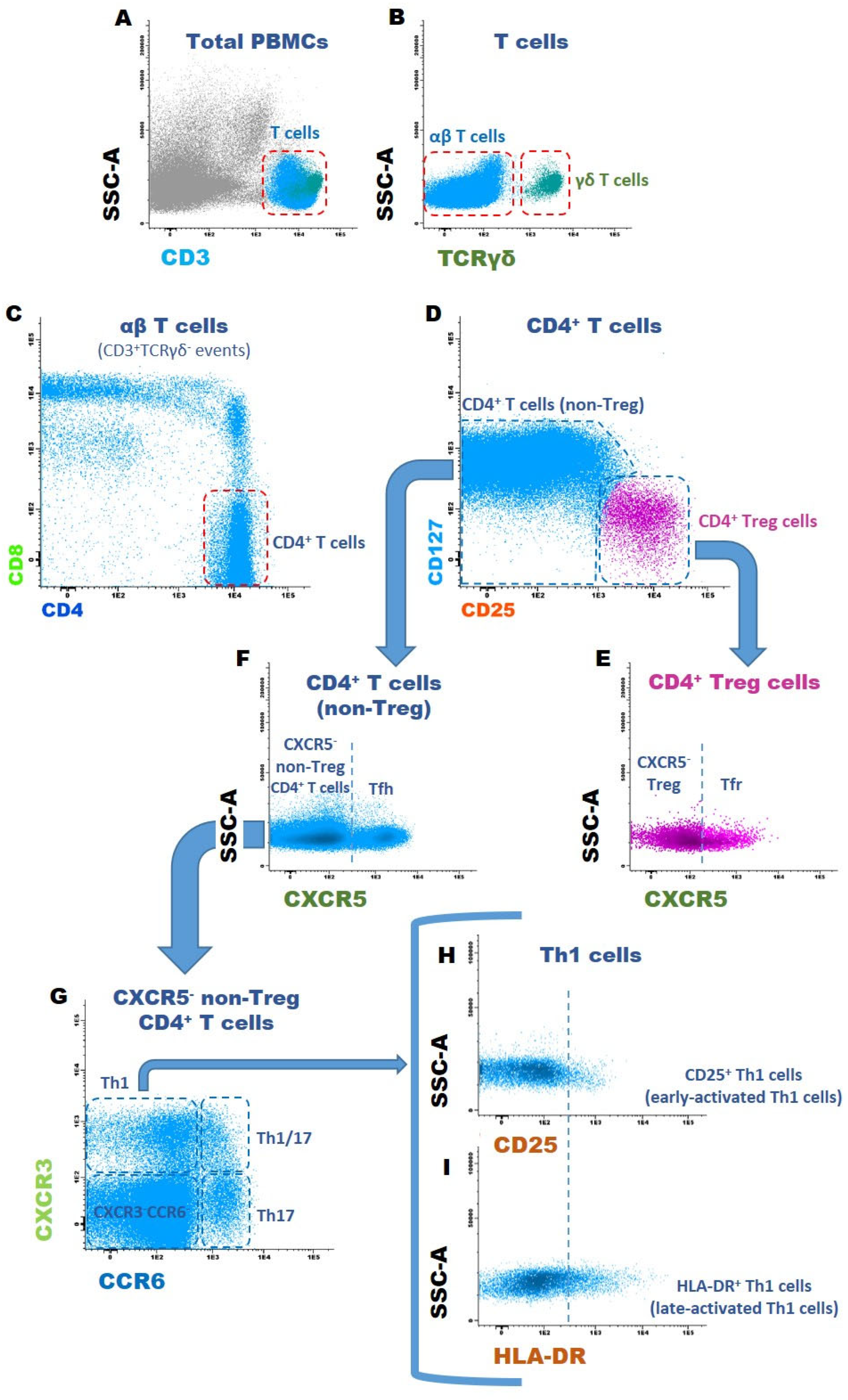
2.5.2. Gating Strategy to Identify T Cell Subsets
2.6. Statistical Analysis
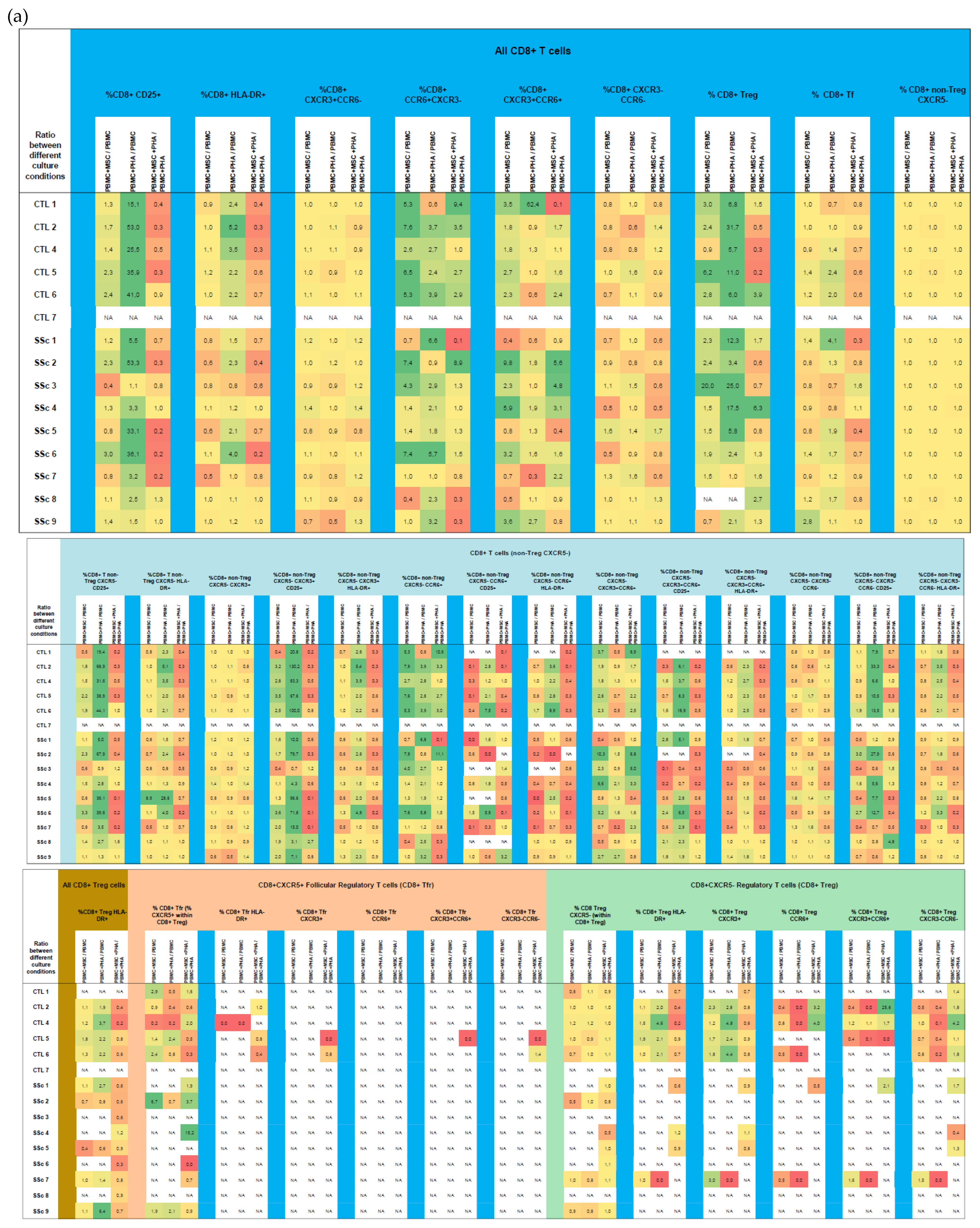
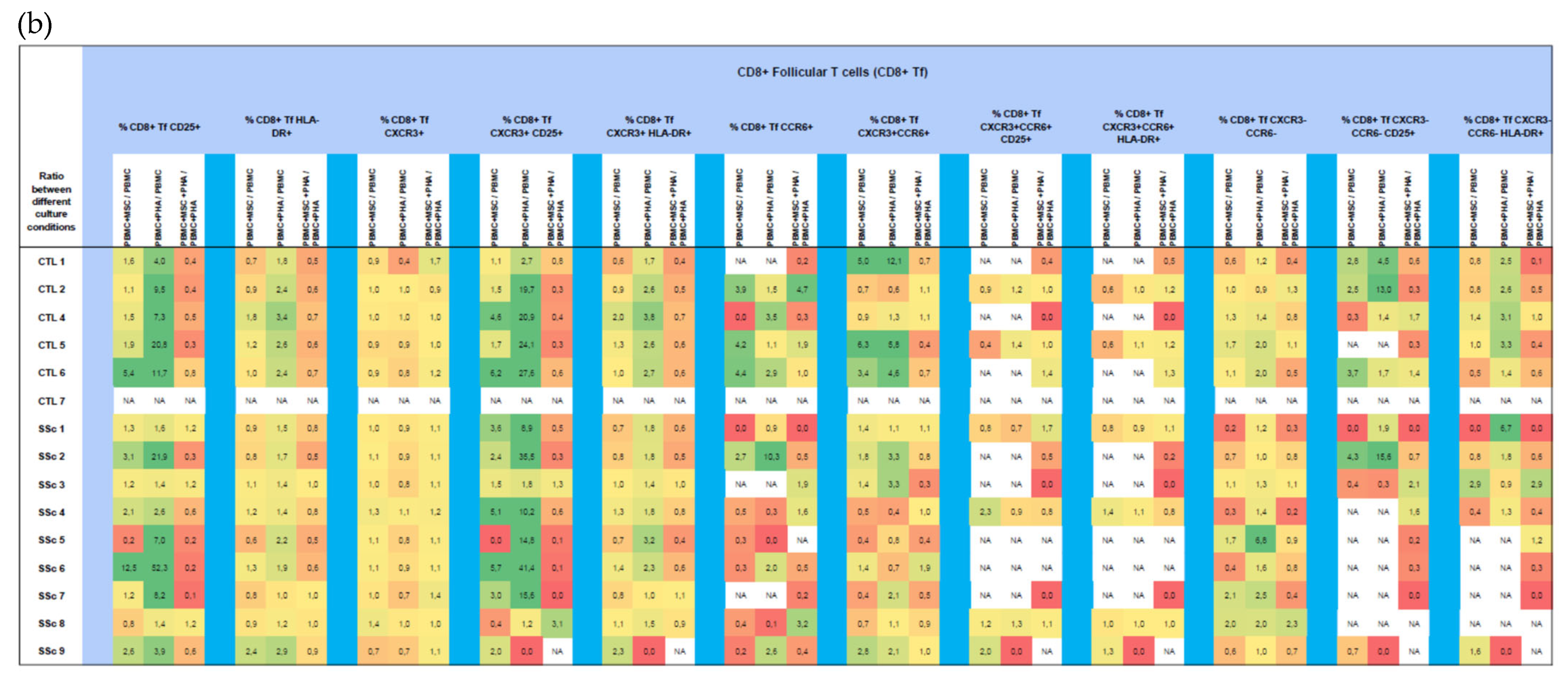
3. Results
3.1. CD4+ T Cells
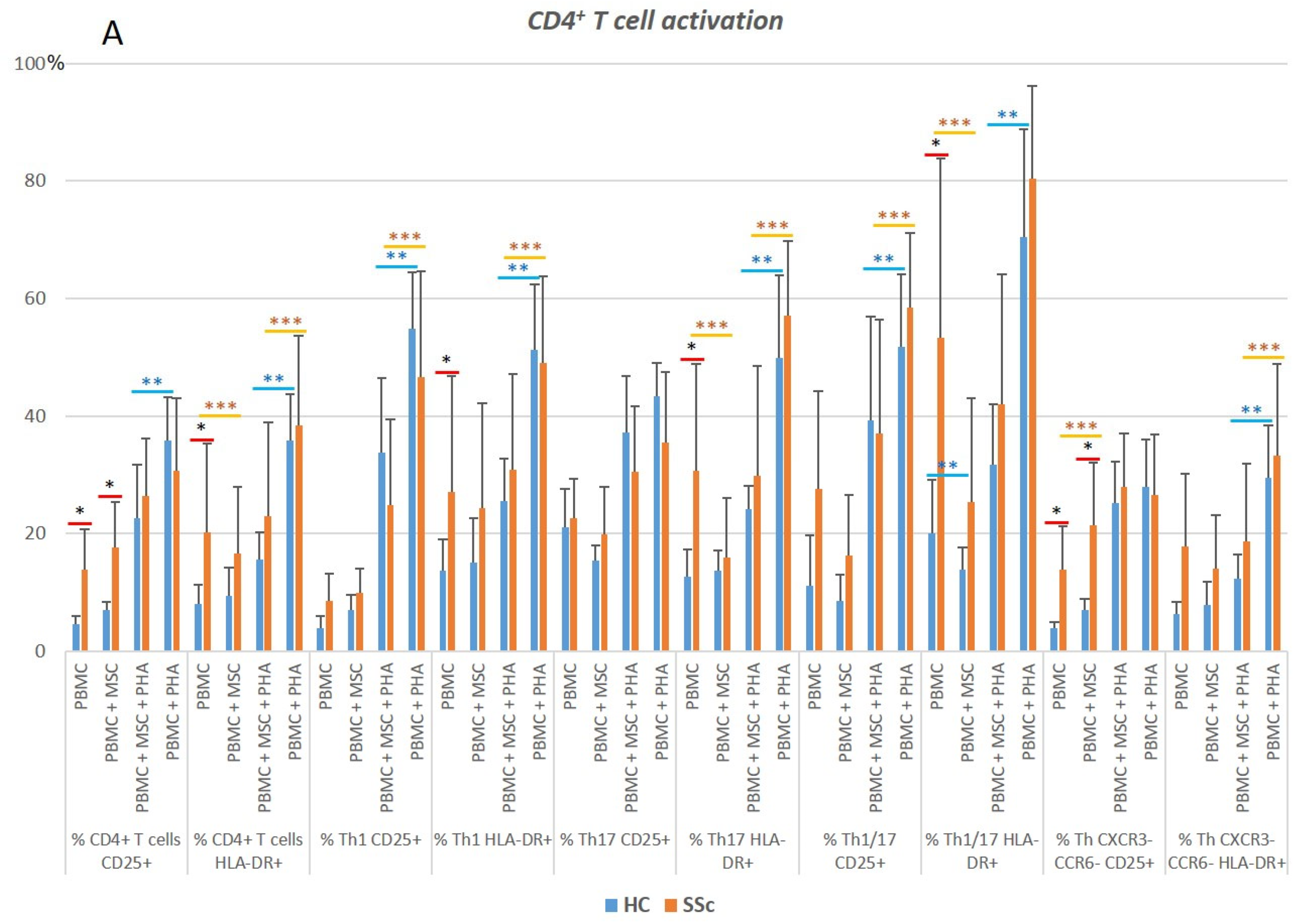
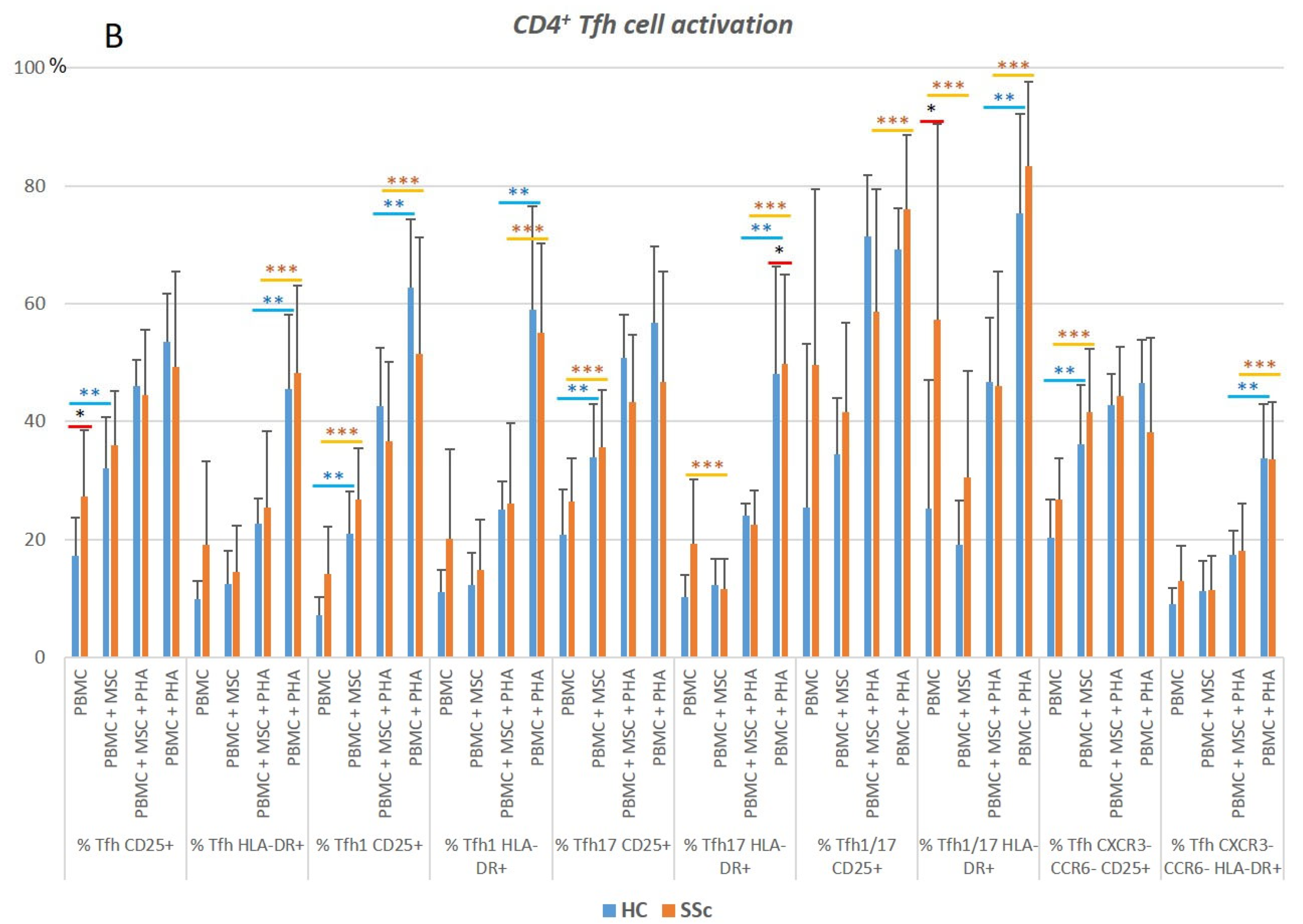
3.1.1. CD4+ T Cells from SSc Patients Display an Increased Activation Status Compared to HC
3.1.2. PHA Stimulation Induces CD4+ T Cells’ Activation and Polarization towards Th1, in Both HC and SSc
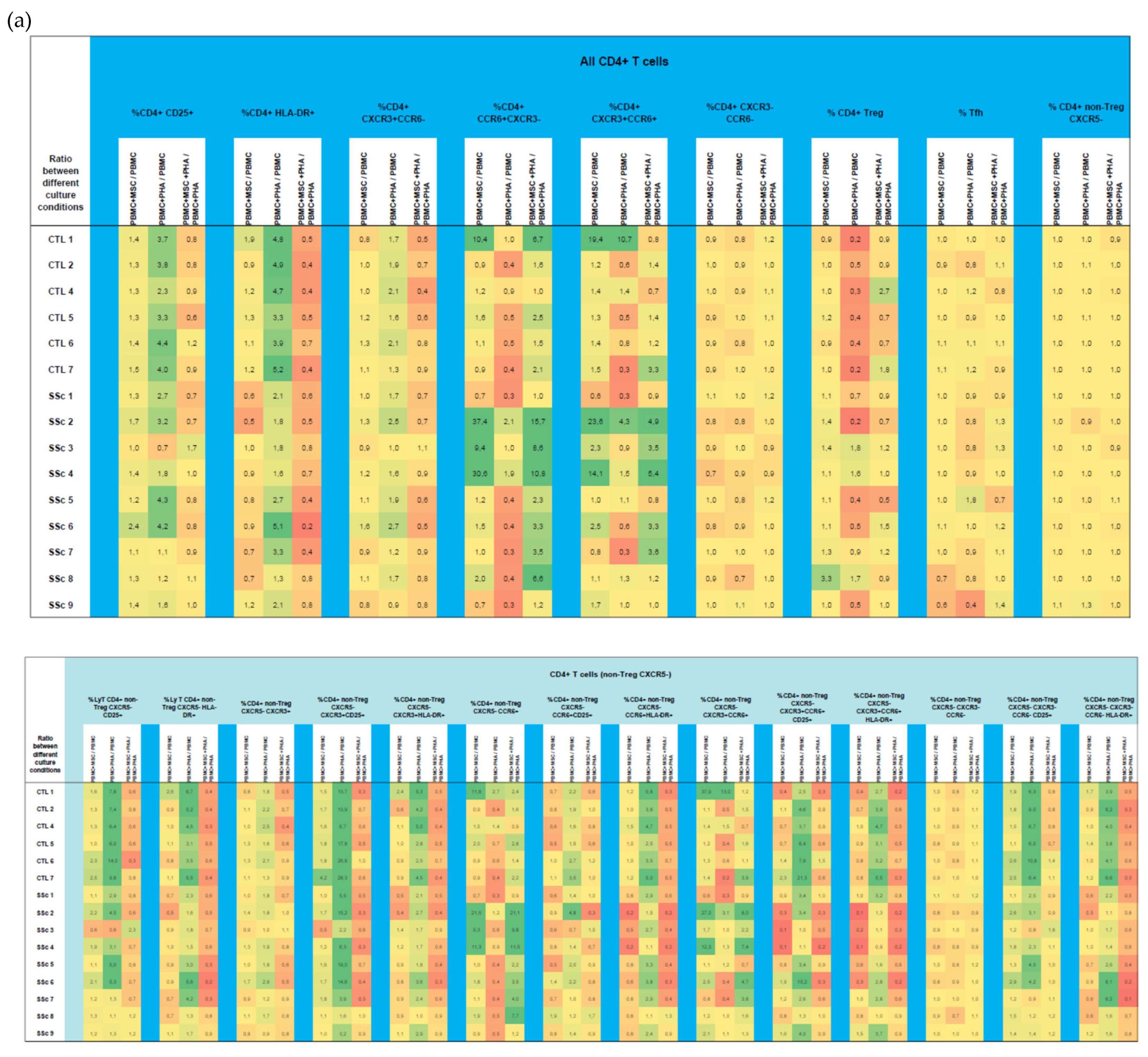
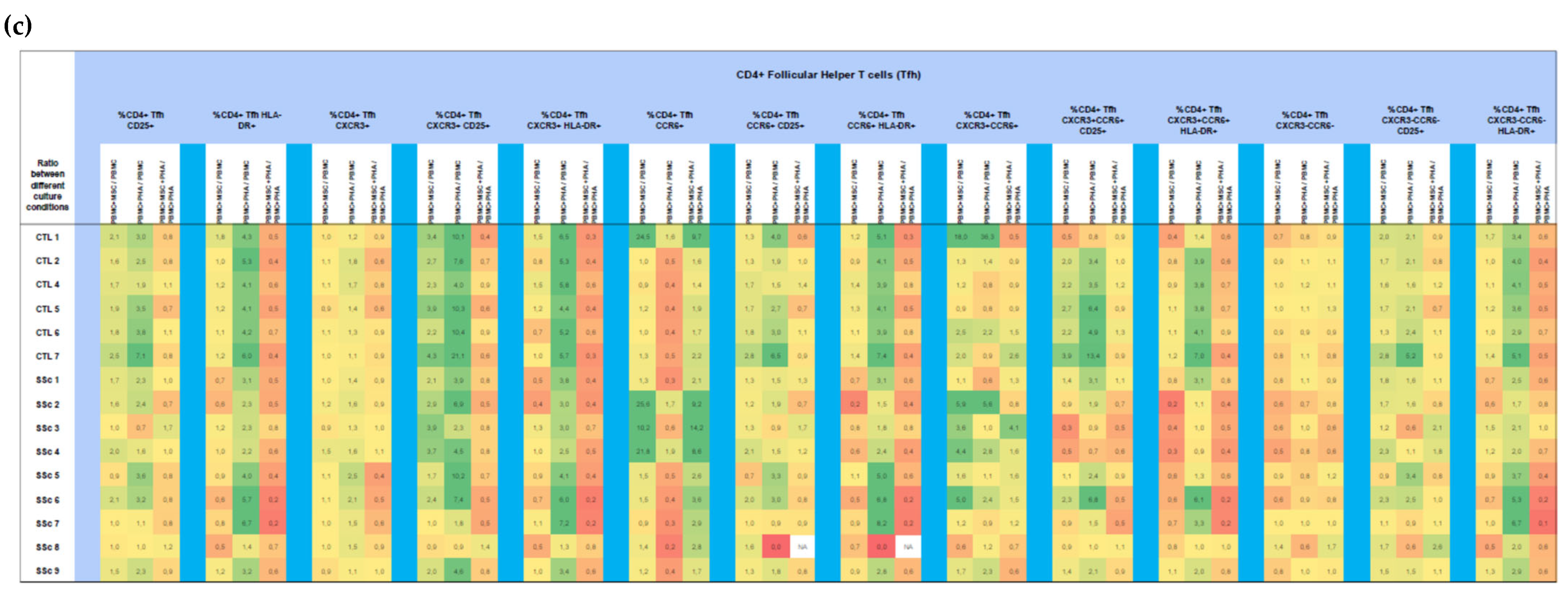
3.1.3. MSCs Control the Excessive Activation Detected in Unstimulated Th17 and Th1/17 Cells from Systemic Sclerosis Patients and Regulate PHA-Induced Cell Activation in HC and SSc
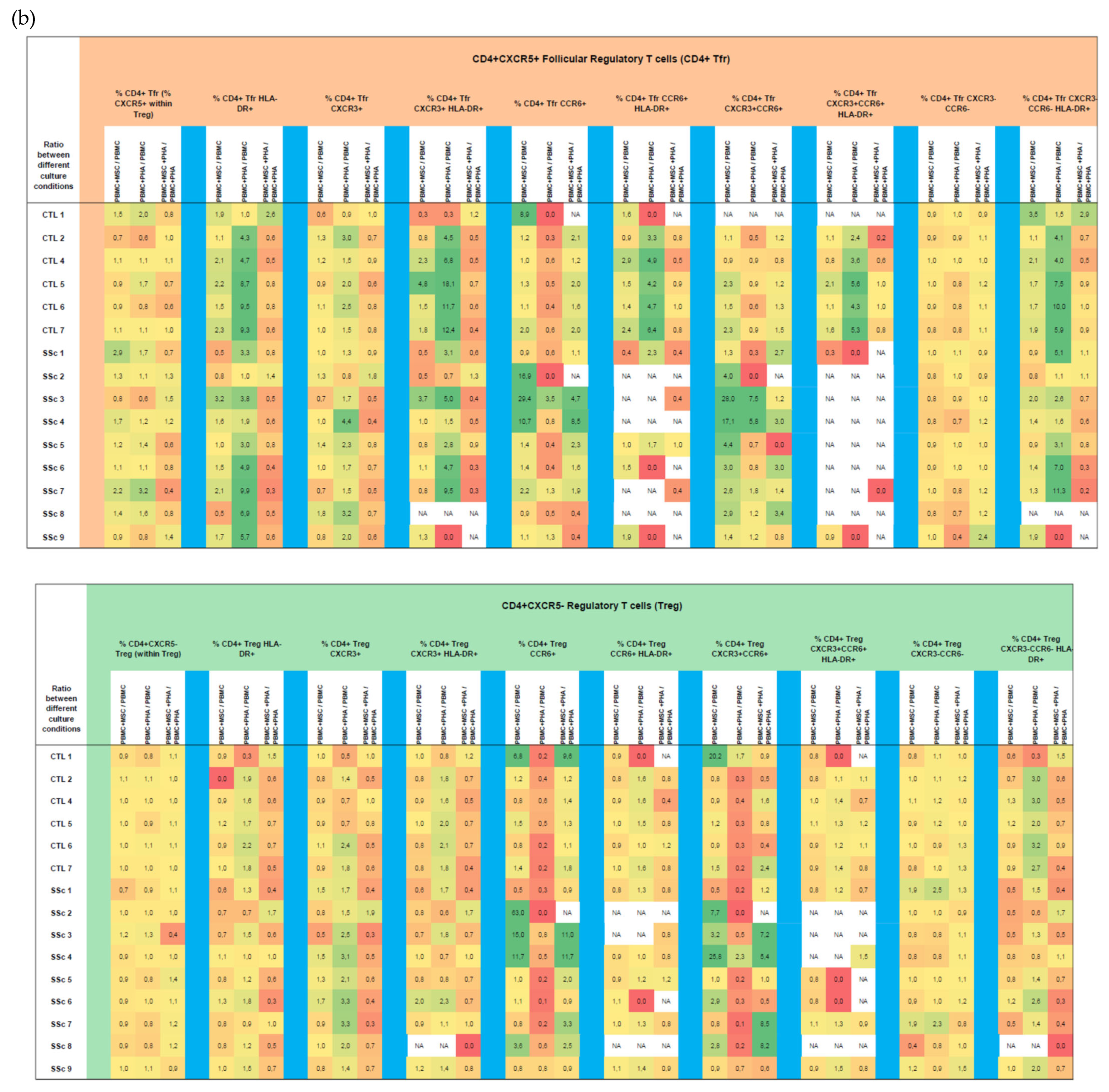
3.1.4. In HC and SSc PBMCs Stimulated with PHA, MSCs Induce Treg, Th17, and Th1/17 Polarization and Suppress PHA-Induced Th1 Differentiation
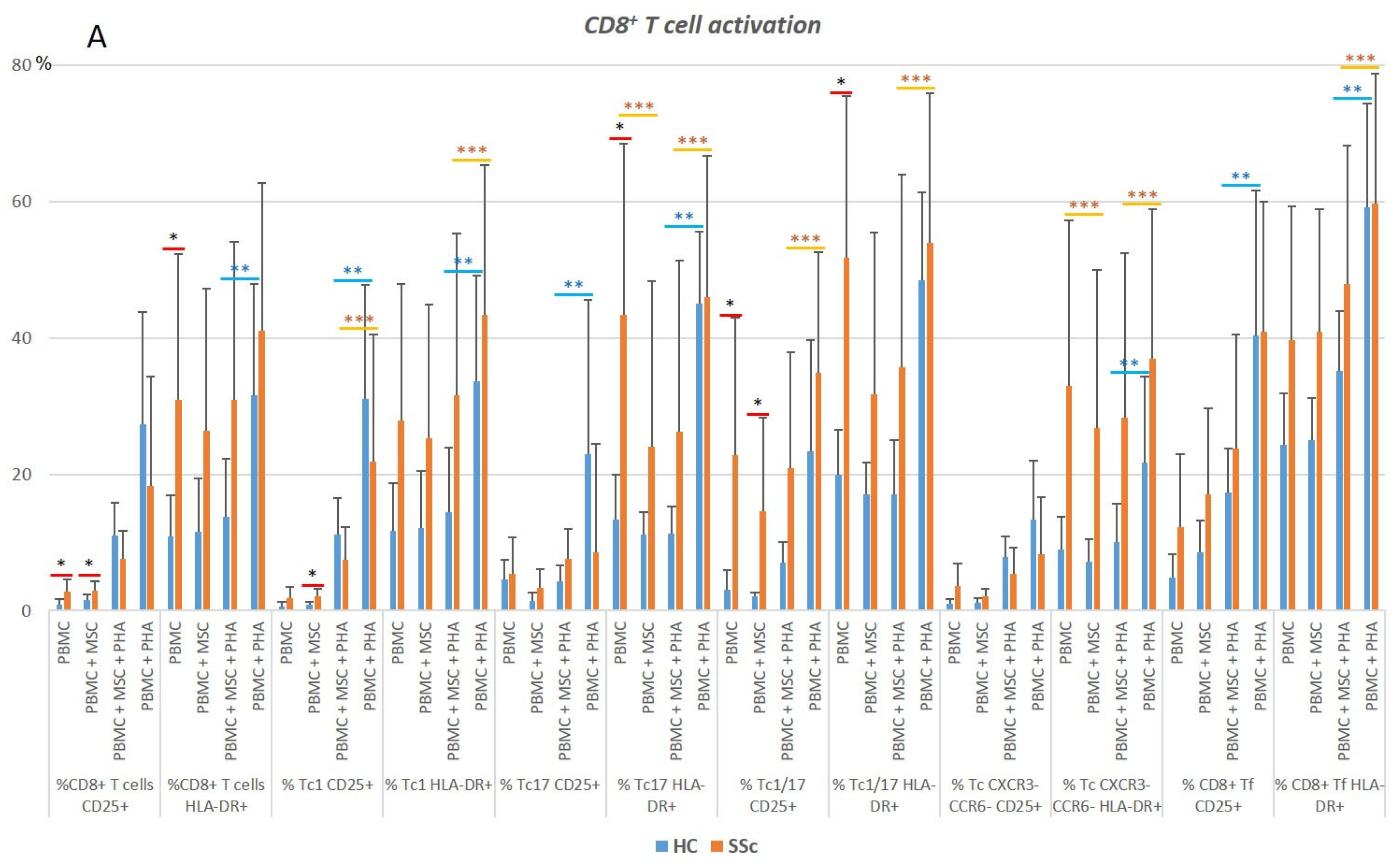
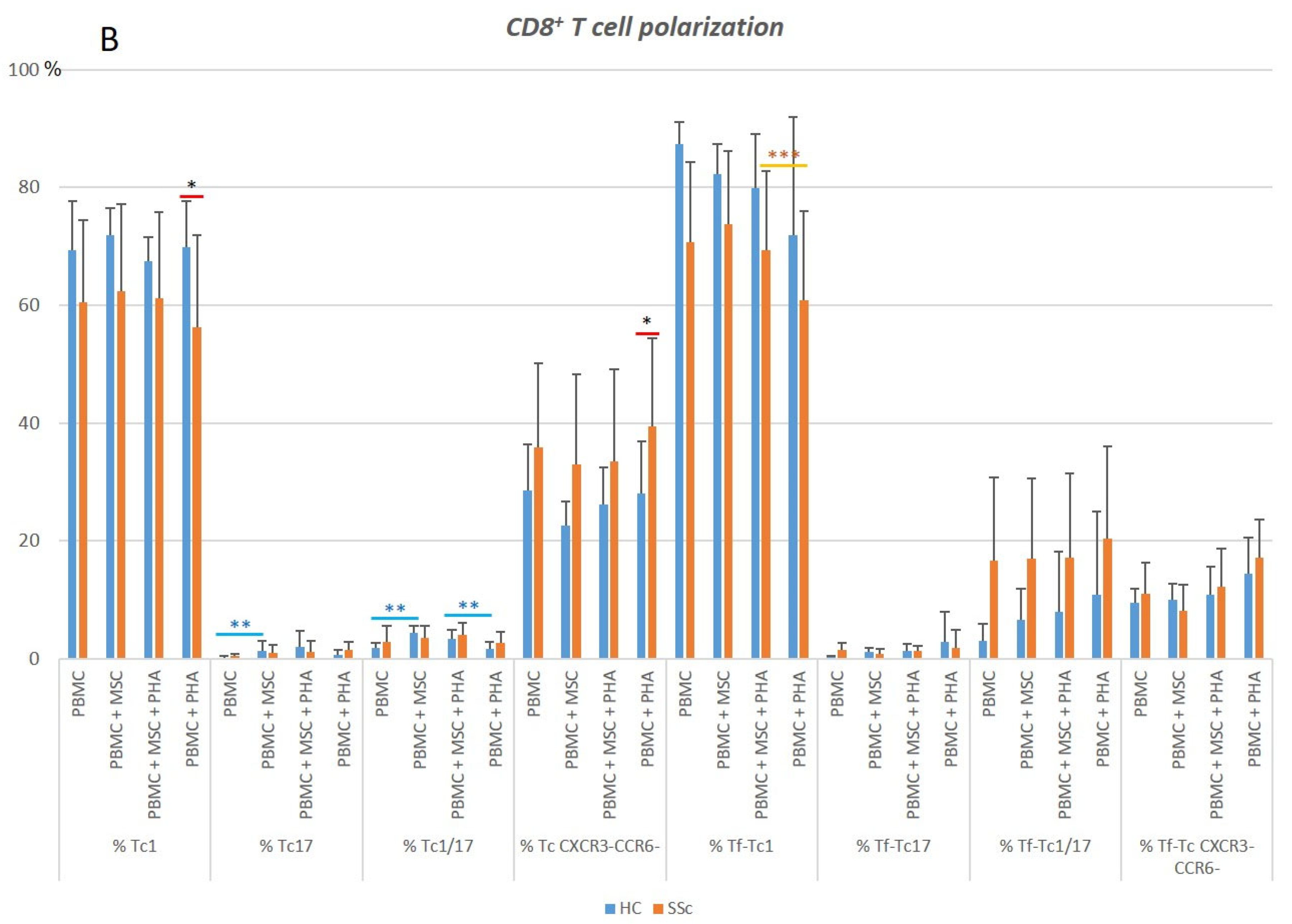
3.2. CD8+ T Cells
3.2.1. In Systemic Sclerosis, There Is an Increased Percentage of Activated CD8+ T Cells, and CD8+ Tf Cells Are Preferentially Polarized into Tf-Tc1/Tc17
3.2.2. PHA Induces CD8+ T Cell Activation but Has Little Effect on CD8+ T Cell Polarization
3.2.3. MSCs Suppress Activation of Tc17 Cells from SSc Patients in Both Non-Stimulated and PHA-Stimulated PBMC Cultures
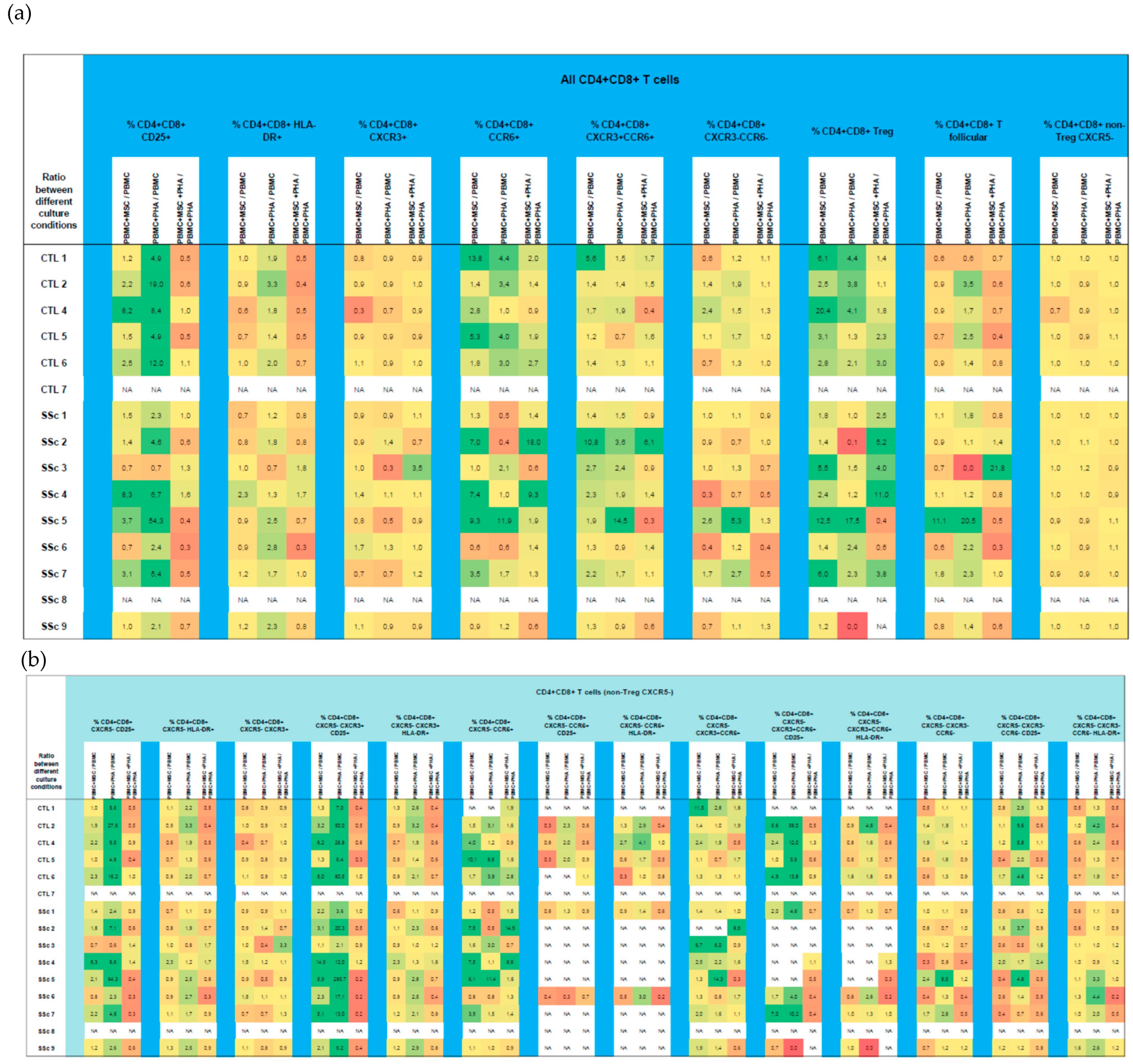
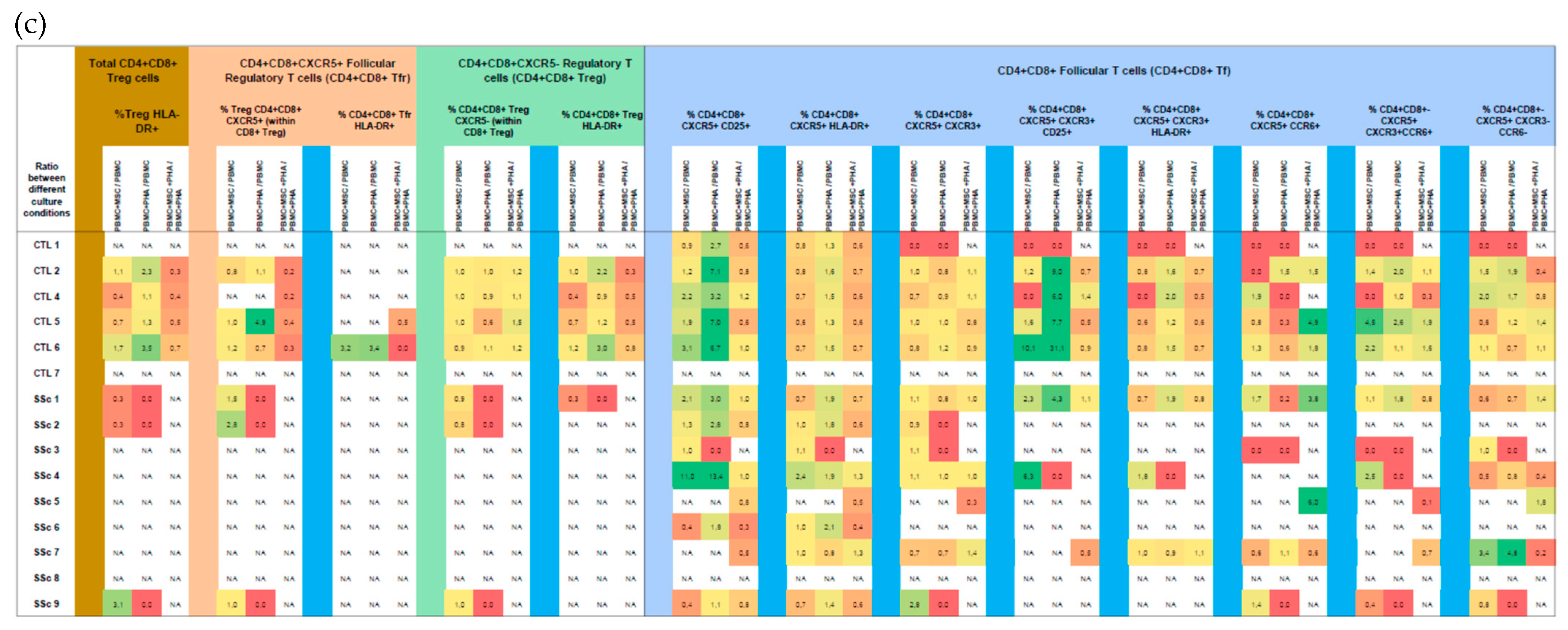
3.3. CD4+CD8+ T Cells
3.3.1. Like CD8+ T Cells, Systemic Sclerosis CD4+CD8+ T Cells Tend to Be More Activated and Polarized into Tf-Tc1/Tc17 Compared to HC
3.3.2. PHA Induces CD4+CD8+ T Cells’ Activation, with No Effect on Their Polarization
3.3.3. MSCs Induce CD4+CD8+ Treg Cells and Suppress PHA-Induced Activation
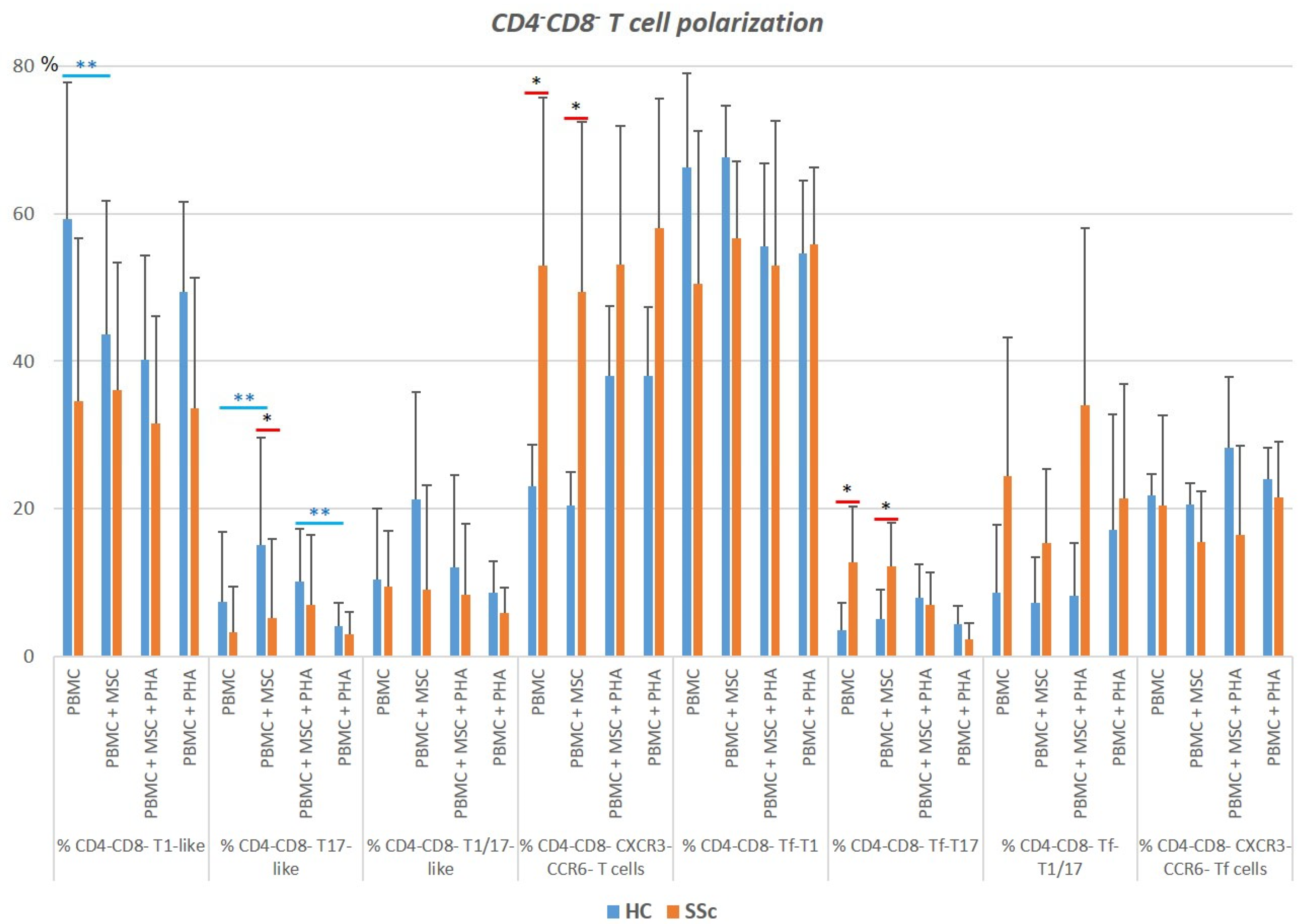
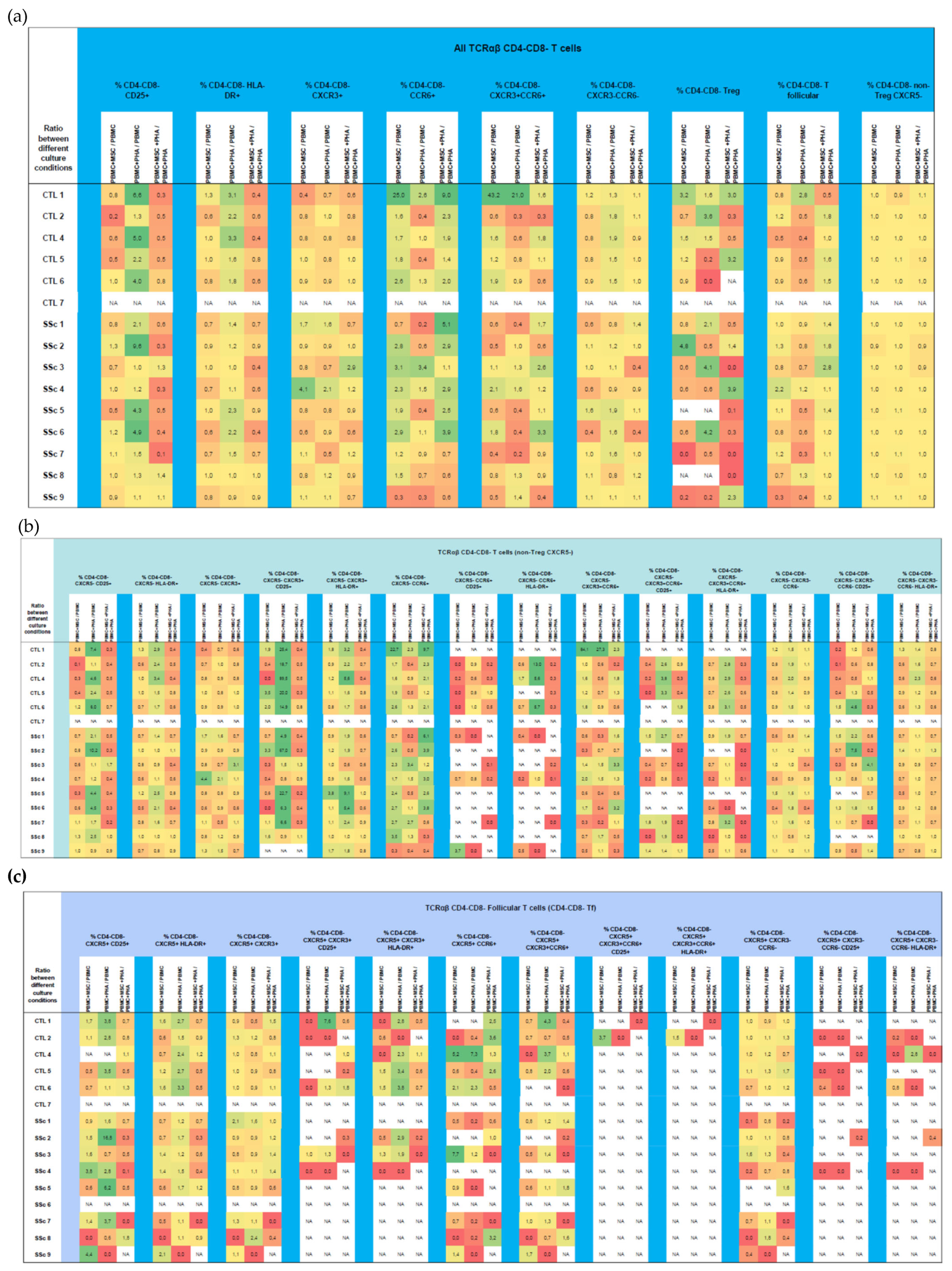
3.4. CD4−CD8−TCRαβ T Cells
3.4.1. Systemic Sclerosis Patients Exhibit a Significant Increase in CXCR3−CCR6−CD4−CD8−TCRαβ T Cells, and Tf Cells Are Preferentially Polarized towards T1/17
3.4.2. PHA Activates CD4−CD8− TCRαβ T Cells Isolated from HC and SSC Patients and Affects Their Polarization in HC
3.4.3. MSCs Regulate Both the Activation and Polarization of CD4−CD8−TCRαβ T Cells
3.5. γδ. T Cells
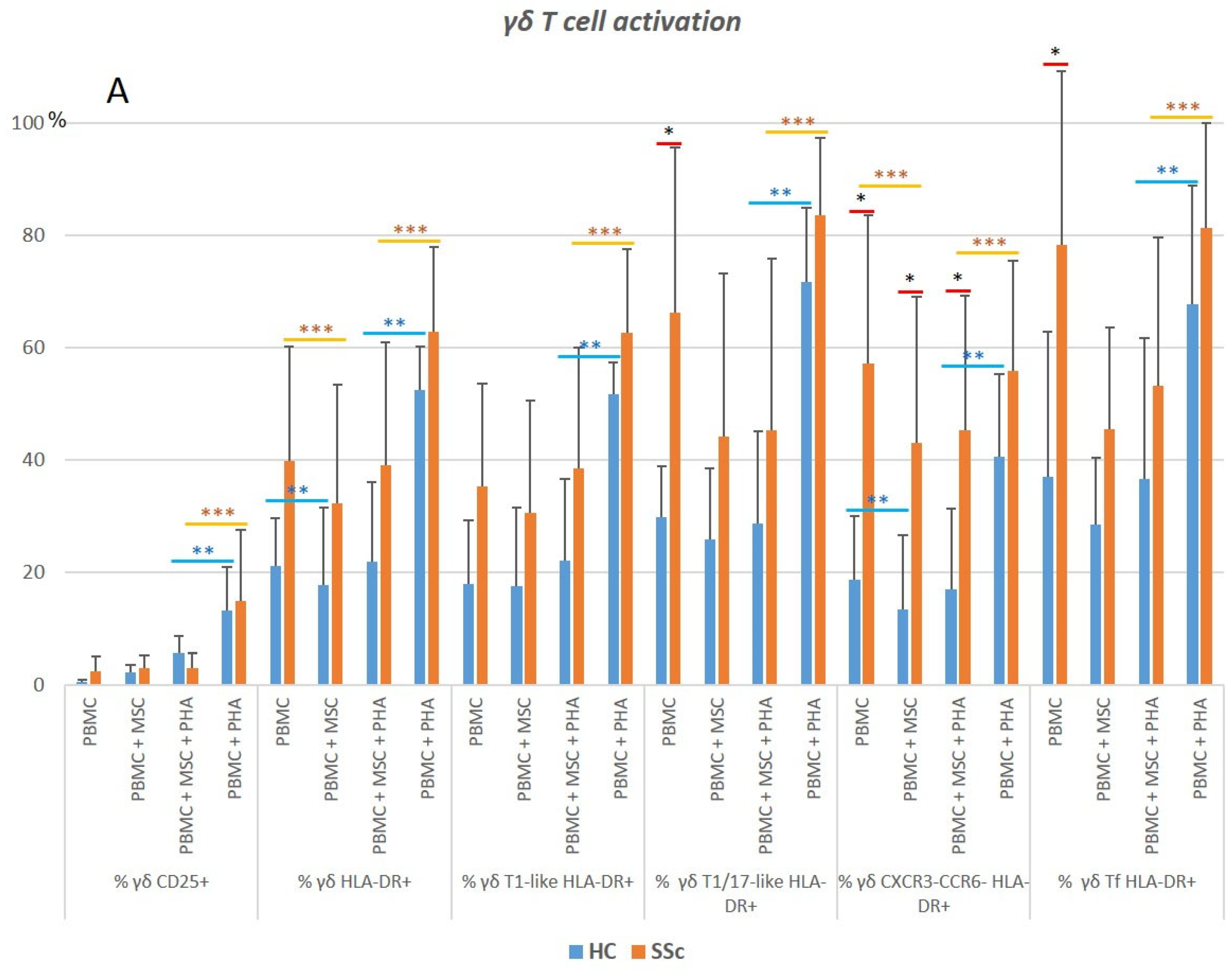
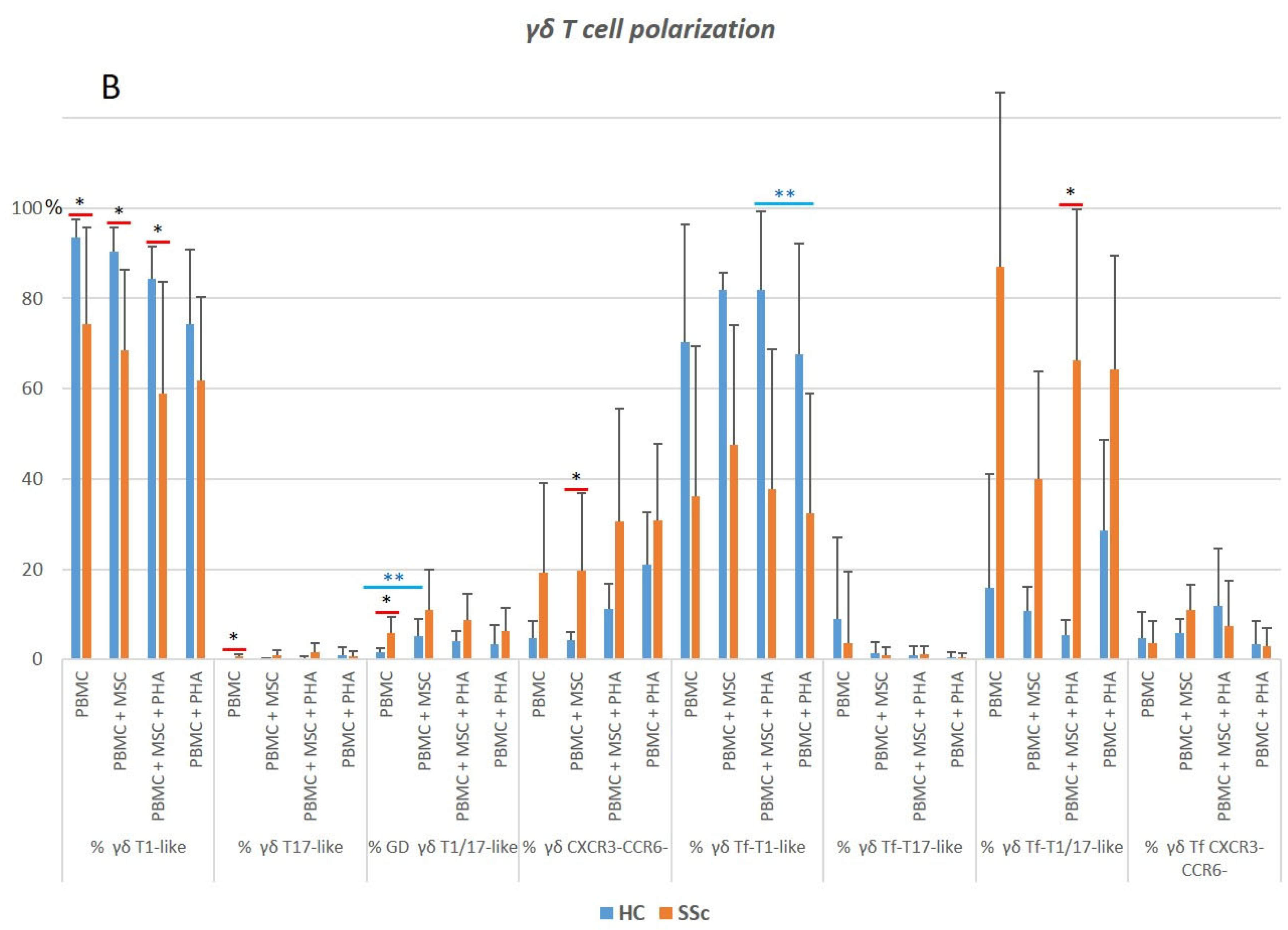
3.5.1. γδ. T Cells from Systemic Sclerosis Patients Display an Increased Activation Status and a Polarization Bias toward T1/17 and CXCR3−CCR6−Cell Compartments
3.5.2. PHA Induces Activation and Modulates Polarization of γδ T Cells in HC and SSc
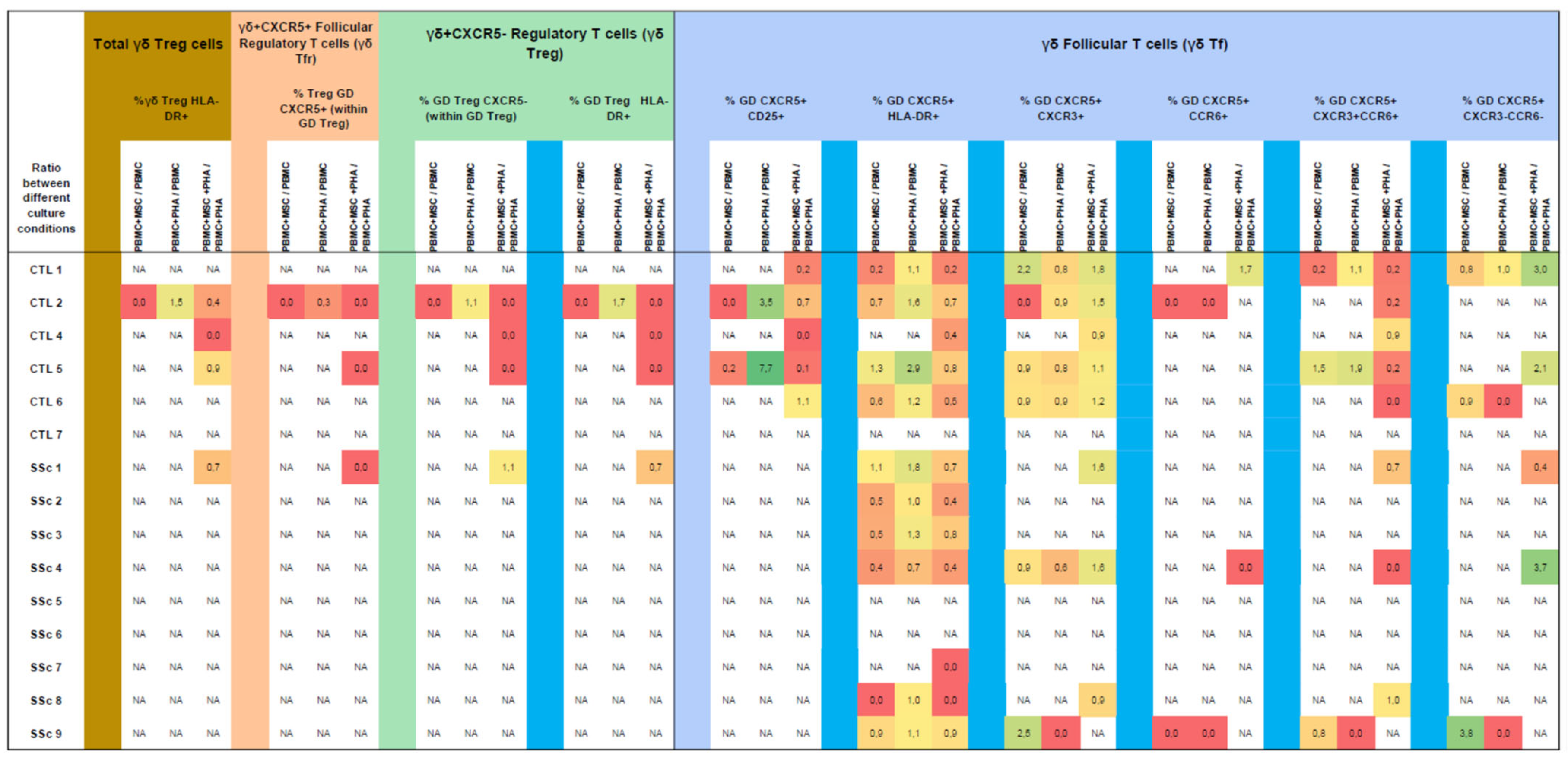
3.5.3. MSCs Inhibit the Strongly Activated Tf γδ T Cells Isolated from SSc Patients
3.6. Comparison between ACA and Anti-Scl-70 SSc Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Allanore, Y.; Simms, R.; Distler, O.; Trojanowska, M.; Pope, J.; Denton, C.P.; Varga, J. Systemic sclerosis. Nat. Rev. Dis. Prim. 2015, 1, 15002. [Google Scholar] [CrossRef] [PubMed]
- Denton, C.P.; Khanna, D. Systemic sclerosis. Lancet 2017, 390, 1685–1699. [Google Scholar] [CrossRef] [PubMed]
- Korman, B. Evolving insights into the cellular and molecular pathogenesis of fibrosis in systemic sclerosis. Transl. Res. 2019, 209, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Van Caam, A.; Vonk, M.; van den Hoogen, F.; van Lent, P.; van der Kraan, P. Unraveling SSc Pathophysiology; The Myofibroblast. Front. Immunol. 2018, 9, 2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, Y. Systemic sclerosis. J. Dermatol. 2018, 45, 128–138. [Google Scholar] [CrossRef]
- Abraham, D.J.; Varga, J. Scleroderma: From cell and molecular mechanisms to disease models. Trends Immunol. 2005, 26, 587–595. [Google Scholar] [CrossRef]
- Furue, M.; Mitoma, C.; Mitoma, H.; Tsuji, G.; Chiba, T.; Nakahara, T.; Uchi, H.; Kadono, T. Pathogenesis of systemic sclerosis-current concept and emerging treatments. Immunol. Res. 2017, 65, 790–797. [Google Scholar] [CrossRef]
- Sierra-Sepulveda, A.; Esquinca-Gonzalez, A.; Benavides-Suarez, S.A.; Sordo-Lima, D.E.; Caballero-Islas, A.E.; Cabral-Castaneda, A.R.; Rodriguez-Reyna, T.S. Systemic Sclerosis Pathogenesis and Emerging Therapies, beyond the Fibroblast. Biomed. Res. Int. 2019, 2019, 4569826. [Google Scholar] [CrossRef] [Green Version]
- Fukasawa, T.; Yoshizaki, A.; Ebata, S.; Yoshizaki-Ogawa, A.; Asano, Y.; Enomoto, A.; Miyagawa, K.; Kazoe, Y.; Mawatari, K.; Kitamori, T.; et al. Single-cell-level protein analysis revealing the roles of autoantigen-reactive B lymphocytes in autoimmune disease and the murine model. eLife 2021, 10, e67209. [Google Scholar] [CrossRef]
- Kuzumi, A.; Yoshizaki, A.; Matsuda, K.M.; Kotani, H.; Norimatsu, Y.; Fukayama, M.; Ebata, S.; Fukasawa, T.; Yoshizaki-Ogawa, A.; Asano, Y.; et al. Interleukin-31 promotes fibrosis and T helper 2 polarization in systemic sclerosis. Nat. Commun. 2021, 12, 5947. [Google Scholar] [CrossRef]
- Frost, J.; Ramsay, M.; Mia, R.; Moosa, L.; Musenge, E.; Tikly, M. Differential gene expression of MMP-1, TIMP-1 and HGF in clinically involved and uninvolved skin in South Africans with SSc. Rheumatology 2012, 51, 1049–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moinzadeh, P.; Krieg, T.; Hellmich, M.; Brinckmann, J.; Neumann, E.; Muller-Ladner, U.; Kreuter, A.; Dumitrescu, D.; Rosenkranz, S.; Hunzelmann, N. Elevated MMP-7 levels in patients with systemic sclerosis: Correlation with pulmonary involvement. Exp. Dermatol. 2011, 20, 770–773. [Google Scholar] [CrossRef] [PubMed]
- Young-Min, S.A.; Beeton, C.; Laughton, R.; Plumpton, T.; Bartram, S.; Murphy, G.; Black, C.; Cawston, T.E. Serum TIMP-1, TIMP-2, and MMP-1 in patients with systemic sclerosis, primary Raynaud’s phenomenon, and in normal controls. Ann. Rheum. Dis. 2001, 60, 846–851. [Google Scholar] [PubMed]
- Rech, T.F.; Moraes, S.B.; Bredemeier, M.; de Paoli, J.; Brenol, J.C.; Xavier, R.M.; Chies, J.A.; Simon, D. Matrix metalloproteinase gene polymorphisms and susceptibility to systemic sclerosis. Genet. Mol. Res. 2016, 15, gmr15049077. [Google Scholar] [CrossRef] [PubMed]
- Fuschiotti, P. T cells and cytokines in systemic sclerosis. Curr. Opin. Rheumatol. 2018, 30, 594–599. [Google Scholar] [CrossRef]
- Brown, M.; O’Reilly, S. The immunopathogenesis of fibrosis in systemic sclerosis. Clin. Exp. Immunol. 2018, 195, 310–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worrell, J.C.; O’Reilly, S. Bi-directional communication: Conversations between fibroblasts and immune cells in systemic sclerosis. J. Autoimmun. 2020, 113, 102526. [Google Scholar] [CrossRef]
- Manetti, M.; Pratesi, S.; Romano, E.; Bellando-Randone, S.; Rosa, I.; Guiducci, S.; Fioretto, B.S.; Ibba-Manneschi, L.; Maggi, E.; Matucci-Cerinic, M. Angiogenic T cell expansion correlates with severity of peripheral vascular damage in systemic sclerosis. PLoS ONE 2017, 12, e0183102. [Google Scholar] [CrossRef] [Green Version]
- Guggino, G.; Lo Pizzo, M.; Di Liberto, D.; Rizzo, A.; Cipriani, P.; Ruscitti, P.; Candore, G.; Gambino, C.M.; Sireci, G.; Dieli, F.; et al. Interleukin-9 over-expression and T helper 9 polarization in systemic sclerosis patients. Clin. Exp. Immunol. 2017, 190, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Zheng, Y.; Zhu, P. T cell abnormalities in systemic sclerosis. Autoimmun. Rev. 2022, 21, 103185. [Google Scholar] [CrossRef]
- Liu, M.; Wu, W.; Sun, X.; Yang, J.; Xu, J.; Fu, W.; Li, M. New insights into CD4(+) T cell abnormalities in systemic sclerosis. Cytokine Growth Factor. Rev. 2016, 28, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Beurier, P.; Ricard, L.; Eshagh, D.; Malard, F.; Siblany, L.; Fain, O.; Mohty, M.; Gaugler, B.; Mekinian, A. TFH cells in systemic sclerosis. J. Transl. Med. 2021, 19, 375. [Google Scholar] [CrossRef] [PubMed]
- Fox, D.A.; Lundy, S.K.; Whitfield, M.L.; Berrocal, V.; Campbell, P.; Rasmussen, S.; Ohara, R.; Stinson, A.; Gurrea-Rubio, M.; Wiewiora, E.; et al. Lymphocyte subset abnormalities in early diffuse cutaneous systemic sclerosis. Arthritis Res. Ther. 2021, 23, 10. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, M.; Gomes, J.; Laranjeira, P.; Duarte, C.; Pedreiro, S.; Antunes, B.; Ribeiro, T.; Santos, F.; Martinho, A.; Fardilha, M.; et al. Immunomodulatory effect of human bone marrow-derived mesenchymal stromal/stem cells on peripheral blood T cells from rheumatoid arthritis patients. J. Tissue Eng. Regen. Med. 2019, 14, 16–28. [Google Scholar] [CrossRef]
- Laranjeira, P.; Pedrosa, M.; Pedreiro, S.; Gomes, J.; Martinho, A.; Antunes, B.; Ribeiro, T.; Santos, F.; Trindade, H.; Paiva, A. Effect of human bone marrow mesenchymal stromal cells on cytokine production by peripheral blood naive, memory, and effector T cells. Stem Cell Res. Ther. 2015, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, A.; Laranjeira, P.; Mendes, S.; Velada, I.; Leite, C.; Andrade, P.; Santos, F.; Henriques, A.; Graos, M.; Cardoso, C.M.; et al. Mesenchymal stem cells from umbilical cord matrix, adipose tissue and bone marrow exhibit different capability to suppress peripheral blood B, natural killer and T cells. Stem Cell Res. Ther. 2013, 4, 125. [Google Scholar] [CrossRef] [Green Version]
- Laranjeira, P.; Duque, M.; Vojtek, M.; Inacio, M.J.; Silva, I.; Mamede, A.C.; Laranjo, M.; Pedreiro, S.; Carvalho, M.J.; Moura, P.; et al. Amniotic membrane extract differentially regulates human peripheral blood T cell subsets, monocyte subpopulations and myeloid dendritic cells. Cell Tissue Res. 2018, 373, 459–476. [Google Scholar] [CrossRef]
- Bukiri, H.; Volkmann, E.R. Current advances in the treatment of systemic sclerosis. Curr. Opin. Pharmacol. 2022, 64, 102211. [Google Scholar] [CrossRef]
- Rozier, P.; Maria, A.; Goulabchand, R.; Jorgensen, C.; Guilpain, P.; Noel, D. Mesenchymal Stem Cells in Systemic Sclerosis: Allogenic or Autologous Approaches for Therapeutic Use? Front. Immunol. 2018, 9, 2938. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, N.; Chang, Y.D.; Hung, S.C.; Lan, J.L.; Liao, J.W.; Fu, S.T.; Lee, C.C. Mesenchymal stem cells suppressed skin and lung inflammation and fibrosis in topoisomerase I-induced systemic sclerosis associated with lung disease mouse model. Cell Tissue Res. 2023, 391, 323–337. [Google Scholar] [CrossRef]
- Rozier, P.; Maumus, M.; Maria, A.T.J.; Toupet, K.; Lai-Kee-Him, J.; Jorgensen, C.; Guilpain, P.; Noel, D. Mesenchymal stromal cells-derived extracellular vesicles alleviate systemic sclerosis via miR-29a-3p. J. Autoimmun. 2021, 121, 102660. [Google Scholar] [CrossRef] [PubMed]
- Gregorius, J.; Wang, C.; Stambouli, O.; Hussner, T.; Qi, Y.; Tertel, T.; Borger, V.; Mohamud Yusuf, A.; Hagemann, N.; Yin, D.; et al. Small extracellular vesicles obtained from hypoxic mesenchymal stromal cells have unique characteristics that promote cerebral angiogenesis, brain remodeling and neurological recovery after focal cerebral ischemia in mice. Basic Res. Cardiol. 2021, 116, 40. [Google Scholar] [CrossRef] [PubMed]
- Granel, B.; Daumas, A.; Jouve, E.; Harle, J.R.; Nguyen, P.S.; Chabannon, C.; Colavolpe, N.; Reynier, J.C.; Truillet, R.; Mallet, S.; et al. Safety, tolerability and potential efficacy of injection of autologous adipose-derived stromal vascular fraction in the fingers of patients with systemic sclerosis: An open-label phase I trial. Ann. Rheum. Dis. 2015, 74, 2175–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Liang, J.; Tang, X.; Wang, D.; Feng, X.; Wang, F.; Hua, B.; Wang, H.; Sun, L. Sustained benefit from combined plasmapheresis and allogeneic mesenchymal stem cells transplantation therapy in systemic sclerosis. Arthritis Res. Ther. 2017, 19, 165. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Lee, Y.J.; Koh, J.H.; Lee, J.; Min, H.K.; Kim, M.Y.; Kim, K.J.; Lee, S.J.; Rhie, J.W.; Kim, W.U.; et al. Clinical Efficacy and Safety of Injection of Stromal Vascular Fraction Derived from Autologous Adipose Tissues in Systemic Sclerosis Patients with Hand Disability: A Proof-Of-Concept Trial. J. Clin. Med. 2020, 9, 3023. [Google Scholar] [CrossRef]
- Farge, D.; Loisel, S.; Resche-Rigon, M.; Lansiaux, P.; Colmegna, I.; Langlais, D.; Charles, C.; Pugnet, G.; Maria, A.T.J.; Chatelus, E.; et al. Safety and preliminary efficacy of allogeneic bone marrow-derived multipotent mesenchymal stromal cells for systemic sclerosis: A single-centre, open-label, dose-escalation, proof-of-concept, phase 1/2 study. Lancet Rheumatol. 2022, 4, e91–e104. [Google Scholar] [CrossRef]
- Cui, J.; Jin, L.; Ding, M.; He, J.; Yang, L.; Cui, S.; Wang, X.; Ma, J.; Liu, A. Efficacy and safety of mesenchymal stem cells in the treatment of systemic sclerosis: A systematic review and meta-analysis. Stem Cell Res. Ther. 2022, 13, 118. [Google Scholar] [CrossRef]
- Daumas, A.; Magalon, J.; Jouve, E.; Casanova, D.; Philandrianos, C.; Abellan Lopez, M.; Mallet, S.; Veran, J.; Auquit-Auckbur, I.; Farge, D.; et al. Adipose tissue-derived stromal vascular fraction for treating hands of patients with systemic sclerosis: A multicentre randomized trial Autologous AD-SVF versus placebo in systemic sclerosis. Rheumatology 2022, 61, 1936–1947. [Google Scholar] [CrossRef]
- Cipriani, P.; Guiducci, S.; Miniati, I.; Cinelli, M.; Urbani, S.; Marrelli, A.; Dolo, V.; Pavan, A.; Saccardi, R.; Tyndall, A.; et al. Impairment of endothelial cell differentiation from bone marrow-derived mesenchymal stem cells: New insight into the pathogenesis of systemic sclerosis. Arthritis Rheum. 2007, 56, 1994–2004. [Google Scholar] [CrossRef]
- Griffin, M.; Ryan, C.M.; Pathan, O.; Abraham, D.; Denton, C.P.; Butler, P.E. Characteristics of human adipose derived stem cells in scleroderma in comparison to sex and age matched normal controls: Implications for regenerative medicine. Stem Cell Res. Ther. 2017, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Hegner, B.; Schaub, T.; Catar, R.; Kusch, A.; Wagner, P.; Essin, K.; Lange, C.; Riemekasten, G.; Dragun, D. Intrinsic Deregulation of Vascular Smooth Muscle and Myofibroblast Differentiation in Mesenchymal Stromal Cells from Patients with Systemic Sclerosis. PLoS ONE 2016, 11, e0153101. [Google Scholar] [CrossRef] [PubMed]
- Vanneaux, V.; Farge-Bancel, D.; Lecourt, S.; Baraut, J.; Cras, A.; Jean-Louis, F.; Brun, C.; Verrecchia, F.; Larghero, J.; Michel, L. Expression of transforming growth factor beta receptor II in mesenchymal stem cells from systemic sclerosis patients. BMJ Open 2013, 3, e001890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taki, Z.; Gostjeva, E.; Thilly, W.; Yaseen, B.; Lopez, H.; Mirza, M.; Hassuji, Z.; Vigneswaran, S.; Ahmed Abdi, B.; Hart, A.; et al. Pathogenic Activation of Mesenchymal Stem Cells Is Induced by the Disease Microenvironment in Systemic Sclerosis. Arthritis Rheumatol. 2020, 72, 1361–1374. [Google Scholar] [CrossRef] [PubMed]
- Christopeit, M.; Schendel, M.; Foll, J.; Muller, L.P.; Keysser, G.; Behre, G. Marked improvement of severe progressive systemic sclerosis after transplantation of mesenchymal stem cells from an allogeneic haploidentical-related donor mediated by ligation of CD137L. Leukemia 2008, 22, 1062–1064. [Google Scholar] [CrossRef] [PubMed]
- Keyszer, G.; Christopeit, M.; Fick, S.; Schendel, M.; Taute, B.M.; Behre, G.; Muller, L.P.; Schmoll, H.J. Treatment of severe progressive systemic sclerosis with transplantation of mesenchymal stromal cells from allogeneic related donors: Report of five cases. Arthritis Rheum. 2011, 63, 2540–2542. [Google Scholar] [CrossRef]
- Guiducci, S.; Porta, F.; Saccardi, R.; Guidi, S.; Ibba-Manneschi, L.; Manetti, M.; Mazzanti, B.; dal Pozzo, S.; Milia, A.F.; Bellando-Randone, S.; et al. Autologous mesenchymal stem cells foster revascularization of ischemic limbs in systemic sclerosis: A case report. Ann. Intern. Med. 2010, 153, 650–654. [Google Scholar] [CrossRef]
- Ebata, S.; Yoshizaki, A.; Oba, K.; Kashiwabara, K.; Ueda, K.; Uemura, Y.; Watadani, T.; Fukasawa, T.; Miura, S.; Yoshizaki-Ogawa, A.; et al. Safety and efficacy of rituximab in systemic sclerosis (DESIRES): Open-label extension of a double blind, investigators-initiated, randomized, placebo-controlled trial. Lancet Rheumatol. 2022, 4, e546–e555. [Google Scholar] [CrossRef]
- Tang, R.; Yu, J.; Shi, Y.; Zou, P.; Zeng, Z.; Tang, B.; Wang, Y.; Ling, G.; Luo, M.; Xiao, R. Safety and efficacy of Rituximab in systemic sclerosis: A systematic review and meta-analysis. Int. Immunopharmacol. 2020, 83, 106389. [Google Scholar] [CrossRef]
- Khanna, D.; Furst, D.E.; Clements, P.J.; Allanore, Y.; Baron, M.; Czirjak, L.; Distler, O.; Foeldvari, I.; Kuwana, M.; Matucci-Cerinic, M.; et al. Standardization of the modified Rodnan skin score for use in clinical trials of systemic sclerosis. J. Scleroderma Relat. Disord. 2017, 2, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Laranjeira, P.; Pedrosa, M.; Duarte, C.; Pedreiro, S.; Antunes, B.; Ribeiro, T.; dos Santos, F.; Martinho, A.; Fardilha, M.; Domingues, M.R.; et al. Human Bone Marrow Mesenchymal Stromal/Stem Cells Regulate the Proinflammatory Response of Monocytes and Myeloid Dendritic Cells from Patients with Rheumatoid Arthritis. Pharmaceutics 2022, 14, 404. [Google Scholar] [CrossRef]
- Pearson, D.R.; Werth, V.P.; Pappas-Taffer, L. Systemic sclerosis: Current concepts of skin and systemic manifestations. Clin. Dermatol. 2018, 36, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, Y. Autoantibody profiles in systemic sclerosis: Predictive value for clinical evaluation and prognosis. J. Dermatol. 2010, 37, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Chrabaszcz, M.; Malyszko, J.; Sikora, M.; Alda-Malicka, R.; Stochmal, A.; Matuszkiewicz-Rowinska, J.; Rudnicka, L. Renal Involvement in Systemic Sclerosis: An Update. Kidney Blood Press. Res. 2020, 45, 532–548. [Google Scholar] [CrossRef]
- Likhit, O.; Louthrenoo, W.; Pattanakitsakul, S.N.; Suttitheptumrong, A.; Hannongbua, S.; Rungrotmongkol, T.; Noguchi, H.; Takeuchi, F.; Boonnak, K. Determination of T Cell Responses in Thai Systemic Sclerosis Patients. J. Immunol. Res. 2022, 2022, 5072154. [Google Scholar] [CrossRef]
- Rabquer, B.J.; Tsou, P.S.; Hou, Y.; Thirunavukkarasu, E.; Haines, G.K., 3rd; Impens, A.J.; Phillips, K.; Kahaleh, B.; Seibold, J.R.; Koch, A.E. Dysregulated expression of MIG/CXCL9, IP-10/CXCL10 and CXCL16 and their receptors in systemic sclerosis. Arthritis Res. Ther. 2011, 13, R18. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Li, L.; Tan, Z.; Li, Y.; Yang, J.; Tian, F.; Wang, L.; Ren, Y.; Xu, G.; He, X.; et al. Up-regulation of CC chemokine ligand 20 and its receptor CCR6 in the lesional skin of early systemic sclerosis. Eur. J. Dermatol. 2011, 21, 731–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Banna, N.A.; Vaci, M.; Slauenwhite, D.; Johnston, B.; Issekutz, T.B. CCR4 and CXCR3 play different roles in the migration of T cells to inflammation in skin, arthritic joints, and lymph nodes. Eur. J. Immunol. 2014, 44, 1633–1643. [Google Scholar] [CrossRef]
- Essien, K.I.; Katz, E.L.; Strassner, J.P.; Harris, J.E. Regulatory T Cells Require CCR6 for Skin Migration and Local Suppression of Vitiligo. J. Invest. Dermatol. 2022, 142, 3158–3166.e7. [Google Scholar] [CrossRef]
- Fenoglio, D.; Battaglia, F.; Parodi, A.; Stringara, S.; Negrini, S.; Panico, N.; Rizzi, M.; Kalli, F.; Conteduca, G.; Ghio, M.; et al. Alteration of Th17 and Treg cell subpopulations co-exist in patients affected with systemic sclerosis. Clin. Immunol. 2011, 139, 249–257. [Google Scholar] [CrossRef]
- Yang, C.; Lei, L.; Pan, J.; Zhao, C.; Wen, J.; Qin, F.; Dong, F.; Wei, W. Altered CD4+ T cell and cytokine levels in peripheral blood and skin samples from systemic sclerosis patients and IL-35 in CD4+ T cell growth. Rheumatology 2022, 61, 794–805. [Google Scholar] [CrossRef]
- Boin, F.; de Fanis, U.; Bartlett, S.J.; Wigley, F.M.; Rosen, A.; Casolaro, V. T cell polarization identifies distinct clinical phenotypes in scleroderma lung disease. Arthritis Rheum. 2008, 58, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Truchetet, M.E.; Brembilla, N.C.; Montanari, E.; Allanore, Y.; Chizzolini, C. Increased frequency of circulating Th22 in addition to Th17 and Th2 lymphocytes in systemic sclerosis: Association with interstitial lung disease. Arthritis Res. Ther. 2011, 13, R166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, S.; Nakayamada, S.; Miyazaki, Y.; Yoshikawa, M.; Yoshinari, H.; Satoh, Y.; Todoroki, Y.; Nakano, K.; Satoh, M.; Smith, V.; et al. Distinctive association of peripheral immune cell phenotypes with capillaroscopic microvascular patterns in systemic sclerosis. Rheumatology 2019, 58, 2273–2283. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Li, M.; Zeng, X. Correlation of Th17 cells and CD4(+)CD25(+) regulatory T cells with clinical parameters in patients with systemic sclerosis. Chin. Med. J. 2014, 127, 3557–3561. [Google Scholar] [PubMed]
- Balanescu, P.; Balanescu, E.; Balanescu, A. IL-17 and Th17 cells in systemic sclerosis: A comprehensive review. Rom. J. Intern. Med. 2017, 55, 198–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, C.; Zeng, Z.; Deng, Q.; Ding, Y.; Xiao, R. Imbalance between T helper 17 and regulatory T cell subsets plays a significant role in the pathogenesis of systemic sclerosis. Biomed. Pharmacother. 2018, 108, 177–183. [Google Scholar] [CrossRef]
- Kobayashi, S.; Nagafuchi, Y.; Shoda, H.; Fujio, K. The Pathophysiological Roles of Regulatory T Cells in the Early Phase of Systemic Sclerosis. Front. Immunol. 2022, 13, 900638. [Google Scholar] [CrossRef]
- Ly, N.T.M.; Ueda-Hayakawa, I.; Nguyen, C.T.H.; Huynh, T.N.M.; Kishimoto, I.; Fujimoto, M.; Okamoto, H. Imbalance toward TFH 1 cells playing a role in aberrant B cell differentiation in systemic sclerosis. Rheumatology 2021, 60, 1553–1562. [Google Scholar] [CrossRef]
- Wei, L.; Abraham, D.; Ong, V. The Yin and Yang of IL-17 in Systemic Sclerosis. Front. Immunol. 2022, 13, 885609. [Google Scholar] [CrossRef]
- Truchetet, M.E.; Brembilla, N.C.; Montanari, E.; Lonati, P.; Raschi, E.; Zeni, S.; Fontao, L.; Meroni, P.L.; Chizzolini, C. Interleukin-17A+ cell counts are increased in systemic sclerosis skin and their number is inversely correlated with the extent of skin involvement. Arthritis Rheum. 2013, 65, 1347–1356. [Google Scholar] [CrossRef]
- Dufour, A.M.; Alvarez, M.; Russo, B.; Chizzolini, C. Interleukin-6 and Type-I Collagen Production by Systemic Sclerosis Fibroblasts Are Differentially Regulated by Interleukin-17A in the Presence of Transforming Growth Factor-Beta 1. Front. Immunol. 2018, 9, 1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.J.; Park, Y.; Choi, J.W.; Baek, J.A.; Jeong, H.Y.; Na, H.S.; Moon, Y.M.; Cho, M.L.; Park, S.H. Establishment of a humanized animal model of systemic sclerosis in which T helper-17 cells from patients with systemic sclerosis infiltrate and cause fibrosis in the lungs and skin. Exp. Mol. Med. 2022, 54, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Fukasawa, T.; Yoshizaki, A.; Ebata, S.; Fukayama, M.; Kuzumi, A.; Norimatsu, Y.; Matsuda, K.M.; Kotani, H.; Sumida, H.; Yoshizaki-Ogawa, A.; et al. Interleukin-17 pathway inhibition with brodalumab in early systemic sclerosis: Analysis of a single-arm, open-label, phase 1 trial. J. Am. Acad. Dermatol. 2023, in press. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Larregina, A.T.; Domsic, R.T.; Stolz, D.B.; Medsger, T.A., Jr.; Lafyatis, R.; Fuschiotti, P. Skin-Resident Effector Memory CD8(+)CD28(-) T Cells Exhibit a Profibrotic Phenotype in Patients with Systemic Sclerosis. J. Invest. Dermatol. 2017, 137, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Fuschiotti, P.; Larregina, A.T.; Ho, J.; Feghali-Bostwick, C.; Medsger, T.A., Jr. Interleukin-13-producing CD8+ T cells mediate dermal fibrosis in patients with systemic sclerosis. Arthritis Rheum. 2013, 65, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Schmalzing, M.; Almanzar, G.; Benoit, S.; Hamm, H.; Tony, H.P.; Goebeler, M.; Prelog, M. Contribution of CD8+ T cells to inflammatory cytokine production in systemic sclerosis (SSc). Autoimmunity 2016, 49, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Parel, Y.; Aurrand-Lions, M.; Scheja, A.; Dayer, J.M.; Roosnek, E.; Chizzolini, C. Presence of CD4+CD8+ double-positive T cells with very high interleukin-4 production potential in lesional skin of patients with systemic sclerosis. Arthritis Rheum. 2007, 56, 3459–3467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radstake, T.R.; van Bon, L.; Broen, J.; Wenink, M.; Santegoets, K.; Deng, Y.; Hussaini, A.; Simms, R.; Cruikshank, W.W.; Lafyatis, R. Increased frequency and compromised function of T regulatory cells in systemic sclerosis (SSc) is related to a diminished CD69 and TGFbeta expression. PLoS ONE 2009, 4, e5981. [Google Scholar] [CrossRef]
- Slobodin, G.; Ahmad, M.S.; Rosner, I.; Peri, R.; Rozenbaum, M.; Kessel, A.; Toubi, E.; Odeh, M. Regulatory T cells (CD4(+)CD25(bright)FoxP3(+)) expansion in systemic sclerosis correlates with disease activity and severity. Cell. Immunol. 2010, 261, 77–80. [Google Scholar] [CrossRef]
- Ugor, E.; Simon, D.; Almanzar, G.; Pap, R.; Najbauer, J.; Nemeth, P.; Balogh, P.; Prelog, M.; Czirjak, L.; Berki, T. Increased proportions of functionally impaired regulatory T cell subsets in systemic sclerosis. Clin. Immunol. 2017, 184, 54–62. [Google Scholar] [CrossRef]
- Wynn, T.A. Fibrotic disease and the T(H)1/T(H)2 paradigm. Nat. Rev. Immunol. 2004, 4, 583–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, K.G.; Dawson, N.A.J.; Huang, Q.; Dunne, J.V.; Levings, M.K.; Broady, R. Regulatory T cells produce profibrotic cytokines in the skin of patients with systemic sclerosis. J. Allergy Clin. Immunol. 2015, 135, 946–955.e9. [Google Scholar] [CrossRef] [PubMed]
- Ricard, L.; Jachiet, V.; Malard, F.; Ye, Y.; Stocker, N.; Riviere, S.; Senet, P.; Monfort, J.B.; Fain, O.; Mohty, M.; et al. Circulating follicular helper T cells are increased in systemic sclerosis and promote plasmablast differentiation through the IL-21 pathway which can be inhibited by ruxolitinib. Ann. Rheum. Dis. 2019, 78, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.K.; Mittereder, N.; Kuta, E.; Delaney, T.; Burwell, T.; Dacosta, K.; Zhao, W.; Cheng, L.I.; Brown, C.; Boutrin, A.; et al. T follicular helper-like cells contribute to skin fibrosis. Sci. Transl. Med. 2018, 10, eaaf5307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricard, L.; Malard, F.; Riviere, S.; Laurent, C.; Fain, O.; Mohty, M.; Gaugler, B.; Mekinian, A. Regulatory B cell imbalance correlates with Tfh expansion in systemic sclerosis. Clin. Exp. Rheumatol. 2021, 39 (Suppl. 131), 20–24. [Google Scholar] [CrossRef]
- Wichner, K.; Stauss, D.; Kampfrath, B.; Kruger, K.; Muller, G.; Rehm, A.; Lipp, M.; Hopken, U.E. Dysregulated development of IL-17- and IL-21-expressing follicular helper T cells and increased germinal center formation in the absence of RORgammat. FASEB J. 2016, 30, 761–774. [Google Scholar] [CrossRef]
- Lv, Y.; Ricard, L.; Gaugler, B.; Huang, H.; Ye, Y. Biology and clinical relevance of follicular cytotoxic T cells. Front. Immunol. 2022, 13, 1036616. [Google Scholar] [CrossRef]
- Giacomelli, R.; Cipriani, P.; Fulminis, A.; Barattelli, G.; Matucci-Cerinic, M.; D’Alo, S.; Cifone, G.; Tonietti, G. Circulating gamma/delta T lymphocytes from systemic sclerosis (SSc) patients display a T helper (Th) 1 polarization. Clin. Exp. Immunol. 2001, 125, 310–315. [Google Scholar] [CrossRef]
- Henriques, A.; Silva, C.; Santiago, M.; Henriques, M.J.; Martinho, A.; Trindade, H.; da Silva, J.A.; Silva-Santos, B.; Paiva, A. Subset-specific alterations in frequencies and functional signatures of gammadelta T cells in systemic sclerosis patients. Inflamm. Res. 2016, 65, 985–994. [Google Scholar] [CrossRef]
- Giacomelli, R.; Matucci-Cerinic, M.; Cipriani, P.; Ghersetich, I.; Lattanzio, R.; Pavan, A.; Pignone, A.; Cagnoni, M.L.; Lotti, T.; Tonietti, G. Circulating Vdelta1+ T cells are activated and accumulate in the skin of systemic sclerosis patients. Arthritis Rheum. 1998, 41, 327–334. [Google Scholar] [CrossRef]
- Ueda-Hayakawa, I.; Hasegawa, M.; Hamaguchi, Y.; Takehara, K.; Fujimoto, M. Circulating gamma/delta T cells in systemic sclerosis exhibit activated phenotype and enhance gene expression of proalpha2(I) collagen of fibroblasts. J. Dermatol. Sci. 2013, 69, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Migalovich Sheikhet, H.; Villacorta Hidalgo, J.; Fisch, P.; Balbir-Gurman, A.; Braun-Moscovici, Y.; Bank, I. Dysregulated CD25 and Cytokine Expression by gammadelta T Cells of Systemic Sclerosis Patients Stimulated With Cardiolipin and Zoledronate. Front. Immunol. 2018, 9, 753. [Google Scholar] [CrossRef] [PubMed]
- Segawa, S.; Goto, D.; Horikoshi, M.; Kondo, Y.; Umeda, N.; Hagiwara, S.; Yokosawa, M.; Hirota, T.; Miki, H.; Tsuboi, H.; et al. Involvement of CD161+ Vdelta1+ gammadelta T cells in systemic sclerosis: Association with interstitial pneumonia. Rheumatology 2014, 53, 2259–2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muschaweckh, A.; Petermann, F.; Korn, T. IL-1beta and IL-23 Promote Extrathymic Commitment of CD27(+)CD122(-) gammadelta T Cells to gammadeltaT17 Cells. J. Immunol. 2017, 199, 2668–2679. [Google Scholar] [CrossRef] [Green Version]
- Schirmer, L.; Rothhammer, V.; Hemmer, B.; Korn, T. Enriched CD161high CCR6+ gammadelta T cells in the cerebrospinal fluid of patients with multiple sclerosis. JAMA Neurol. 2013, 70, 345–351. [Google Scholar] [CrossRef] [Green Version]






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Gender | Age | SSc Subtype | Disease Duration (Years) | Auto-Antibody Profile | Organ Involvement | Digital Ulcers | mRSS | SSc Treatment | SSc Conditions for Treatment | Comorbidities and Treatment |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SSc 1 | F | 57 | dcSSc | 5 | anti-Scl-70 | None | No | 4 | Mycophenolate mofetil, Prednisolone | Skin involvement | Breast carcinoma: Anastrozole Dyslipidemia: Atorvastatin, Ezetimibe |
| SSc 2 | F | 33 | dcSSc | 4 | anti-Scl-70 | Lung | Yes | 18 | Mycophenolate mofetil, Nifedipine | Lung involvement, Raynaud phenomenon, digital ulcers | Depression: Fluoxetine |
| SSc 3 | F | 72 | lcSSc | 7 | ACA | Heart | No | 10 | Bosentan, Sildenafil | Pulmonary arterial hipertension | Hypothyroidism: Levothyroxine Depression: Mirtazapine, Quetiapione, Alprazolam |
| SSc 4 | F | 36 | lcSSc | 5 | anti-Scl-70 | None | No | 0 | Pentoxifylline, Nifedipine | Raynaud phenomenon | None |
| SSc 5 | F | 69 | lcSSc | 10 | ACA | None | No | 0 | Pentoxifylline | Raynaud phenomenon | Depression: Escitalopram Osteoporosis: Alendronic acid+Calcium +VitaminD Dyslipidemia: Rosuvastatin Arrhythmia: Bisoprolol |
| SSc 6 | F | 40 | lcSSc | 5 | anti-Scl-70 | None | No | 10 | Methotrexate, Prednisolone, Amlodipine | Skin involvement, Raynaud phenomenon | Depression: Trazodone, Paroxetine |
| SSc 7 | F | 64 | lcSSc | 5 | ACA | None | Yes | 6 | Pentoxifylline, ASA 100 | Raynaud phenomenon, digital ulcers | Cardiac insufficiency: Furosemide, Sacubitril+Valsartan, Bisoprolol, Spironolactone, Ivabradine, ASA 100 Gastric Ulcer: Pantoprazole Depression: Escitalopram, Zolpidem |
| SSc 8 | F | 82 | lcSSc | 9 | ACA | None | Yes | 6 | Azathioprine, Pentoxifylline | Raynaud phenomenon | Dyslipidemia: Rosuvastatin Cardiac insufficiency: Furosemide, Valsartan, Spironolactone Osteoporosis: Alendronic acid+Calcium +VitaminD |
| SSc 9 | F | 62 | dcSSc | 3 | anti-Scl-70 | Lung | Yes | 21 | Mycophenolate mofetil, Bosentan, ASA 100, Nifedipine | Lung involvement, Raynaud phenomenon, digital ulcers | Osteoporosis: Alendronic acid+Calcium +VitaminD Depression: Venlafaxine, Alprazolam |
| Fluorochrome | FITC | PE | PerCP-Cy5.5 | PC7 | APC | Alexa Fluor 700 | APC Alexa 750 | V450 | BV510 | BV605 |
|---|---|---|---|---|---|---|---|---|---|---|
| Antibody marker | TCRγδ | CXCR3 | CCR6 | CD25 | CD4 | CD8 | CD3 | HLA-DR | CD127 | CXCR5 |
| Clone | IMMU 510 | 1C6 | 11A9 | B1.49.9 | 13B8.2 | B9.11 | UCHT1 | L243 | HIL-7R-M21 | J252D4 |
| Commercial source | Beckman Coulter | BD Pharmingen | BD Pharmingen | Beckman Coulter | Beckman Coulter | Beckman Coulter | Beckman Coulter | BD | BD | BioLegend |
| Number of T Cell Subsets Whose Early Activation (CD25+) is Downregulated by MSCs | Number of T Cell Subsets Whose Late Activation (HLA-DR+) Is Downregulated by MSCs | Number of T Cell Subsets Whose (Early or Late) Activation Is Downregulated by MSCs | Number of T Cell Subsets Whose Polarization is Modulated by MSCs | |
|---|---|---|---|---|
| HC | ||||
| CD4+ T cells | 5 out of 8 | 11 out of 16 | 11 out of 16 | 10 out of 18 |
| CD8+ T cells | 3 out of 7 | 6 out of 9 | 6 out of 9 | 3 out of 14 |
| CD4+CD8+ T cells | 1 out of 5 | 3 out of 7 | 3 out of 7 | 3 out of 12 |
| CD4−CD8− T cells | 2 out of 6 | 4 out of 6 | 5 out of 6 | 3 out of 10 |
| γδ T cells | 2 out of 4 | 4 out of 4 | 4 out of 4 | 3 out of 10 |
| Total (HC) | 13 out of 30 | 28 out of 42 | 29 out of 42 | 22 out of 64 |
| SSc | ||||
| CD4+ T cells | 4 out of 8 | 10 out of 16 | 10 out of 16 | 10 out of 18 |
| CD8+ T cells | 2 out of 7 | 6 out of 9 | 7 out of 9 | 1 out of 14 |
| CD4+CD8+ T cells | 2 out of 5 | 0 out of 7 | 2 out of 7 | 2 out of 10 |
| CD4−CD8− T cells | 1 out of 5 | 3 out of 5 | 3 out of 5 | 0 out of 10 |
| γδ T cells | 1 out of 1 | 4 out of 4 | 4 out of 4 | 0 out of 6 |
| Total (SSc) | 10 out of 26 | 23 out of 41 | 26 out of 41 | 13 out of 58 |
| HC (n = 6) Mean ± Standard Deviation | ACA SSc (n = 4) Mean ± Standard Deviation | Anti-Scl-70 SSc (n = 5) Mean ± Standard Deviation | p Value (ACA vs. Anti-Scl-70) | |
|---|---|---|---|---|
| CD4+ T cells | ||||
| % Total CD4+ CD25+ T cells | 4.53 ± 1.53 | 19 ± 5.74 | 10 ± 4.75 | p < 0.07 |
| % Th17 CD25+ | 21 ± 6.51 | 29 ± 4.93 | 18 ± 4.00 | p < 0.05 |
| % Th CXCR3−CCR6− CD25+ | 3.85 ± 1.06 | 19 ± 7.83 | 10 ± 3.20 | p < 0.05 |
| % Tfh CD25+ | 17 ± 6.47 | 35 ± 14.6 | 21 ± 4.66 | p < 0.07 |
| % Tfh CXCR3−CCR6− CD25+ | 20 ± 6.41 | 31 ± 6.73 | 23 ± 3.58 | p < 0.07 |
| CD8+ T cells | ||||
| % Tc1 | 69 ± 8.20 | 73 ± 6.81 | 51 ± 14.5 | p < 0.05 |
| % Tc CXCR3−CCR6− | 29 ± 7.80 | 24 ± 4.08 | 45 ± 16.8 | p < 0.05 |
| % CD8+ Tf CXCR3−CCR6− | 9.45 ± 2.47 | 6.14 ± 4.39 | 15 ± 4.85 | p < 0.05 |
| CD4+CD8+ T cells | ||||
| % CD4+CD8+ Treg cells | 1.09 ± 0.65 | 0.27 ± 0.16 | 2.10 ± 1.89 | p < 0.05 |
| CD4−CD8− T cells | ||||
| % CD4−CD8− CXCR3−CCR6− HLA-DR+ | 32 ± 18.4 | 78 ± 16.1 | 43 ± 15.3 | p < 0.07 |
| % CD4−CD8− Tf CXCR3−CCR6− | 22 ± 2.86 | 11 ± 10.5 | 30 ± 15.0 | p < 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laranjeira, P.; dos Santos, F.; Salvador, M.J.; Simões, I.N.; Cardoso, C.M.P.; Silva, B.M.; Henriques-Antunes, H.; Corte-Real, L.; Couceiro, S.; Monteiro, F.; et al. Umbilical-Cord-Derived Mesenchymal Stromal Cells Modulate 26 Out of 41 T Cell Subsets from Systemic Sclerosis Patients. Biomedicines 2023, 11, 1329. https://doi.org/10.3390/biomedicines11051329
Laranjeira P, dos Santos F, Salvador MJ, Simões IN, Cardoso CMP, Silva BM, Henriques-Antunes H, Corte-Real L, Couceiro S, Monteiro F, et al. Umbilical-Cord-Derived Mesenchymal Stromal Cells Modulate 26 Out of 41 T Cell Subsets from Systemic Sclerosis Patients. Biomedicines. 2023; 11(5):1329. https://doi.org/10.3390/biomedicines11051329
Chicago/Turabian StyleLaranjeira, Paula, Francisco dos Santos, Maria João Salvador, Irina N. Simões, Carla M. P. Cardoso, Bárbara M. Silva, Helena Henriques-Antunes, Luísa Corte-Real, Sofia Couceiro, Filipa Monteiro, and et al. 2023. "Umbilical-Cord-Derived Mesenchymal Stromal Cells Modulate 26 Out of 41 T Cell Subsets from Systemic Sclerosis Patients" Biomedicines 11, no. 5: 1329. https://doi.org/10.3390/biomedicines11051329