
Neurodifferentiation and Neuroprotection Potential of Mesenchymal Stromal Cell-Derived Secretome Produced in Different Dynamic Systems

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Bone Marrow Mesenchymal Stromal Cell (BMSC) Cultures
2.2. BMSC Cultures under Static Conditions
2.3. Microcarrier Preparation
2.3.1. Inoculation of the Vertical-Wheel Bioreactor (VWBR)
2.3.2. Inoculation of the Spinner Flask (SP)
2.4. BMSC Culture in VWBR
2.5. BMSC Culture in SP
2.6. BMSC Conditioning and Secretome Collection
2.7. Metabolite Analysis throughout Cultures
2.8. Immunophenotypic Analysis of BMSC
2.9. Multilineage Differentiation Assays of BMSC
2.10. Expansion of Human Neural Progenitor Cells and Incubation with hBMSC Secretome
2.11. Immunocytochemistry Analysis of Human Neural Progenitor Cells
2.12. Nematode Strains and Culture Conditions
2.13. Quantitative Analysis of Dopaminergic Neuronal Loss
2.14. Membrane Antibody Arrays
2.15. Statistics
3. Results
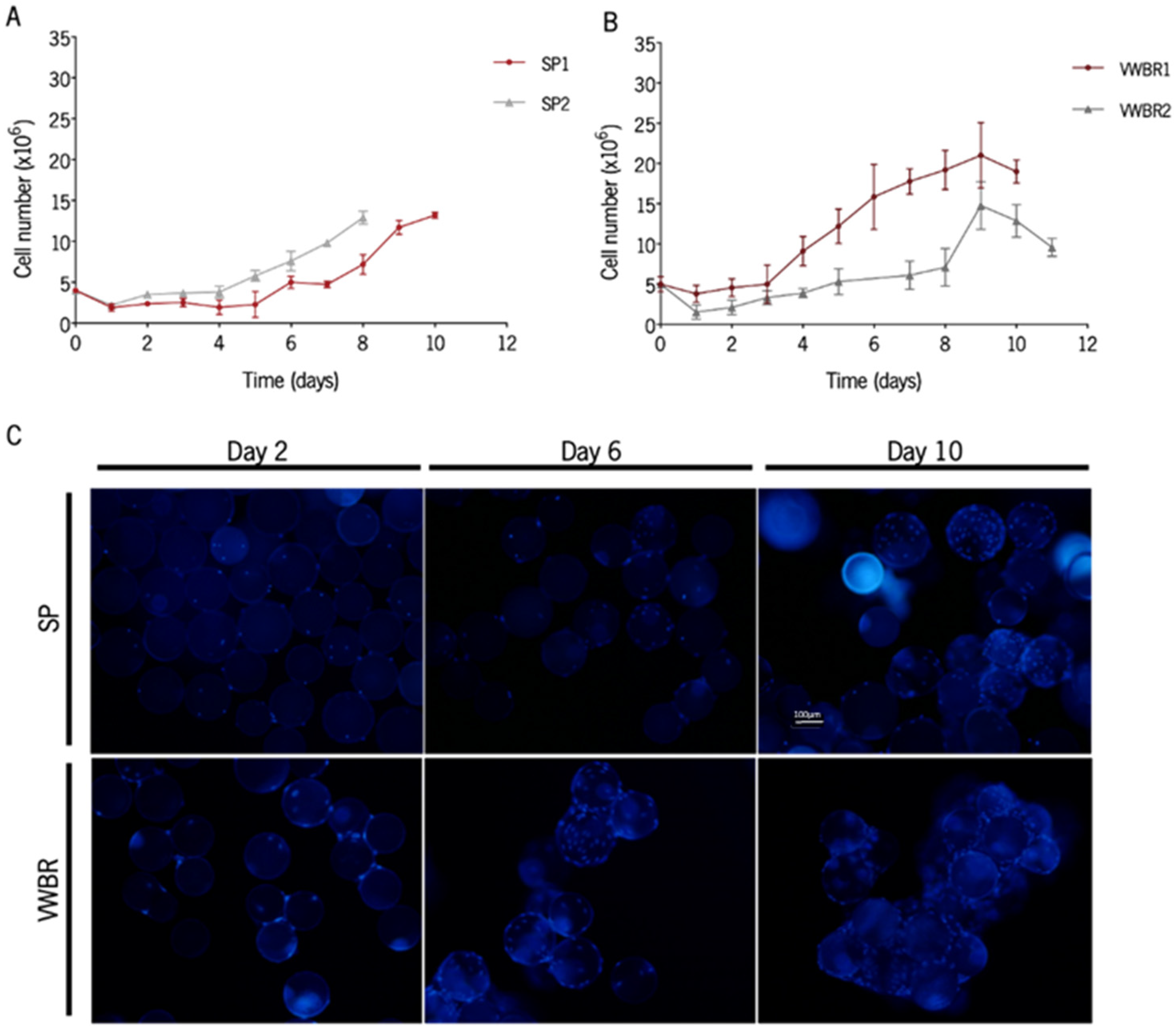
3.1. BMSCs Were Successfully Expanded in Both Dynamic Systems
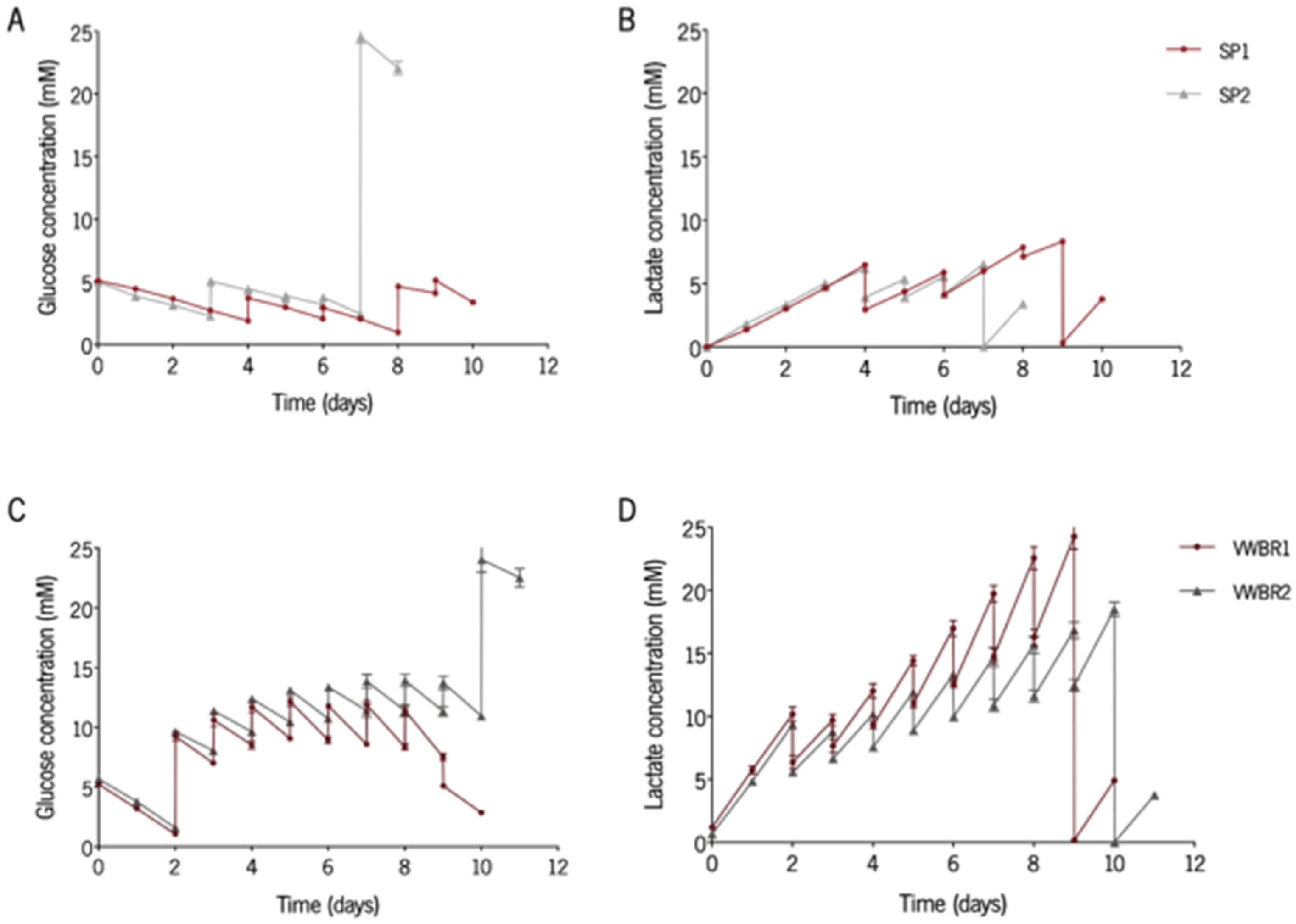
3.2. Glycolitic Metabolism Is Kept Consistent among Culturing Systems
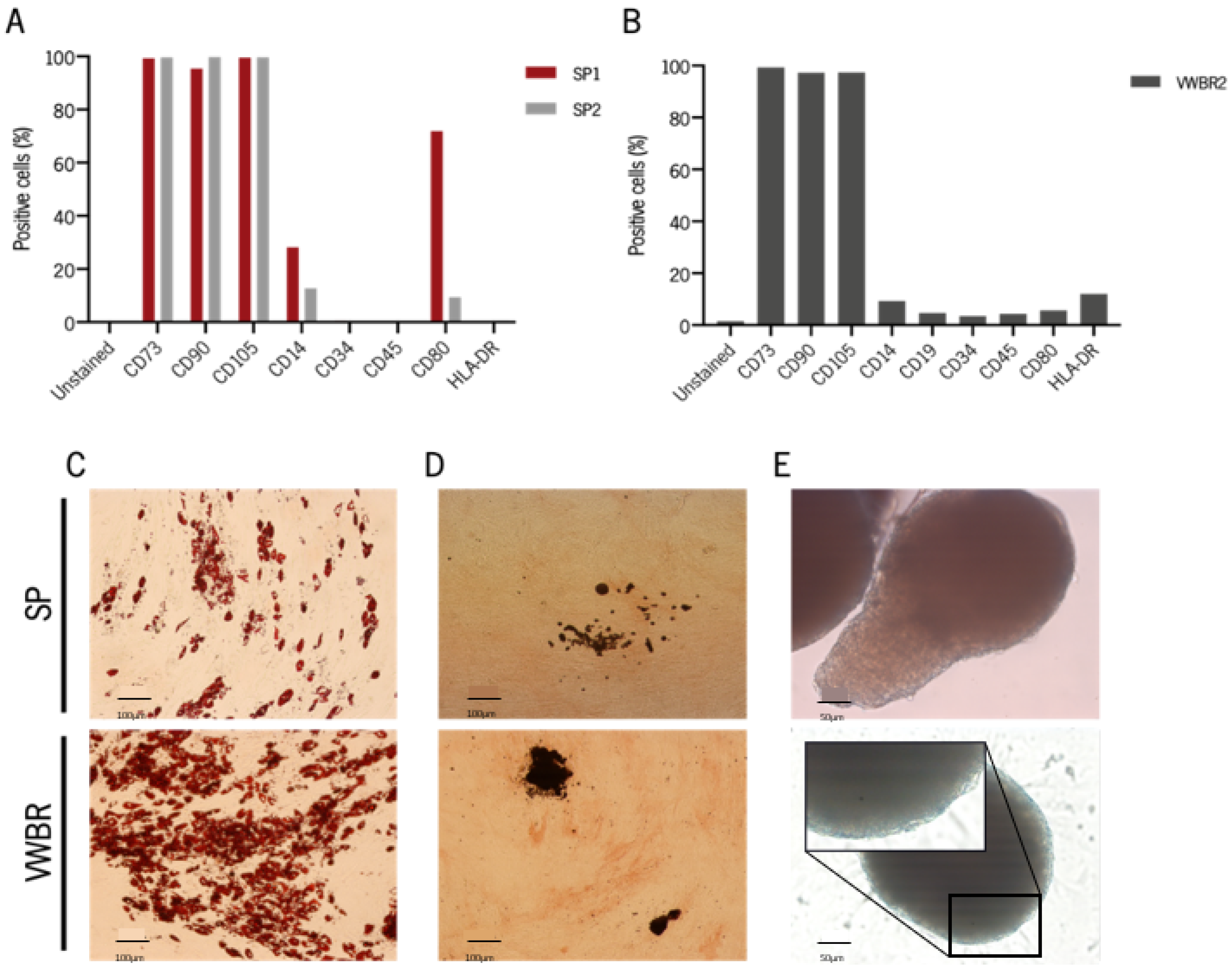
3.3. Cells Expanded in Both Systems Retain MSC Phenotype and Are Capable of Multilineage Differentiation
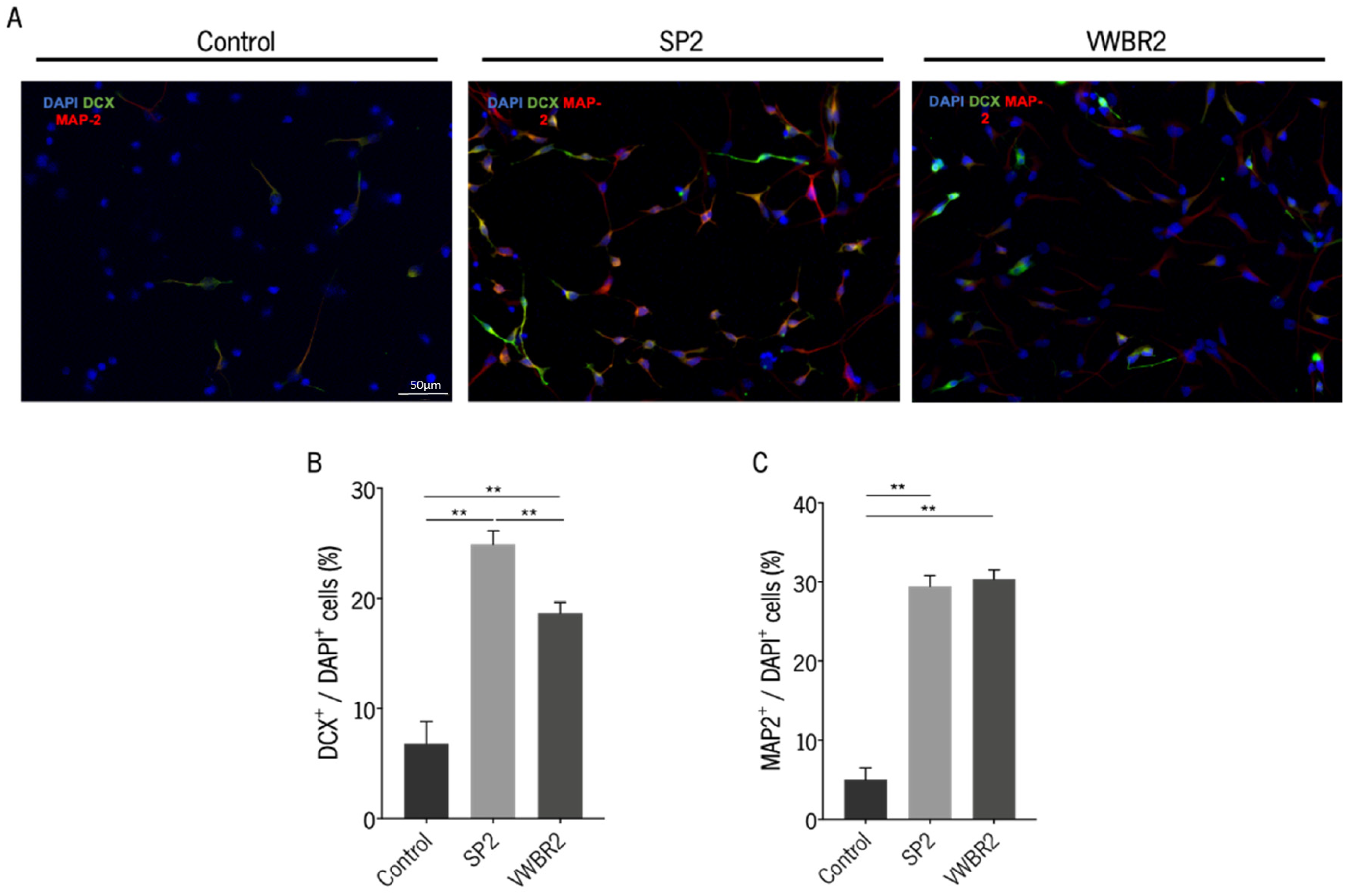
3.4. Secretomes Produced in Both Dynamic Culture Systems Were Able to Induce Neurodifferentiation
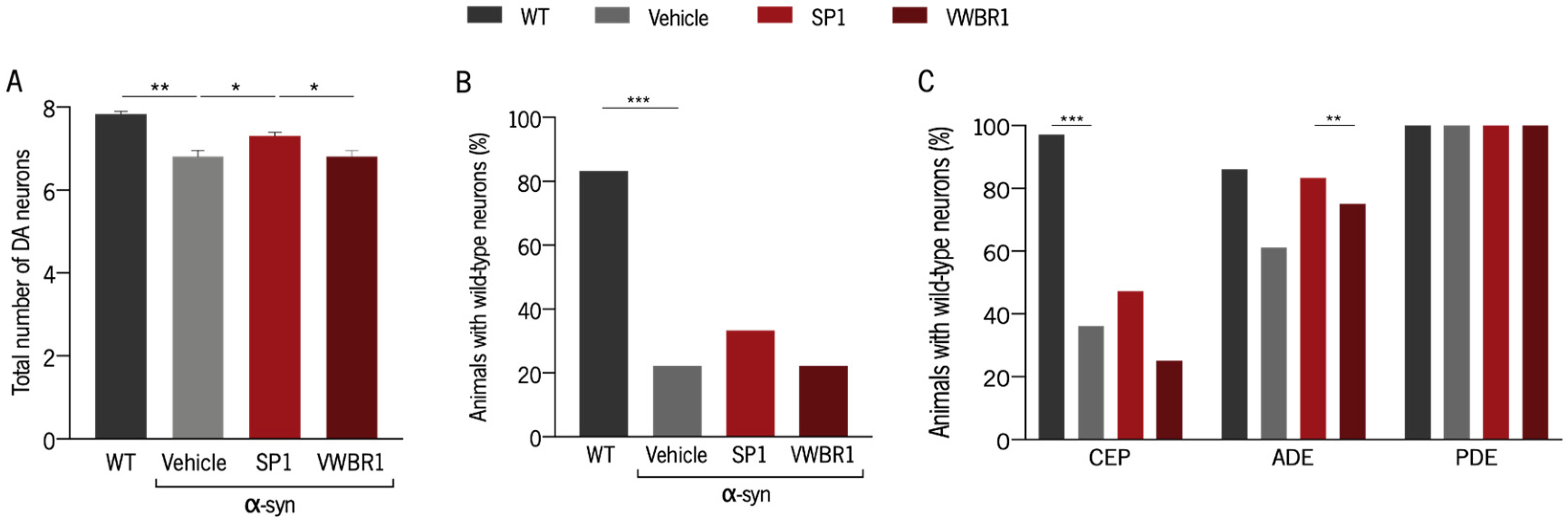
3.5. Distinct Capacity to Induce Neuroprotection Was Shown with Secretomes from Different Systems
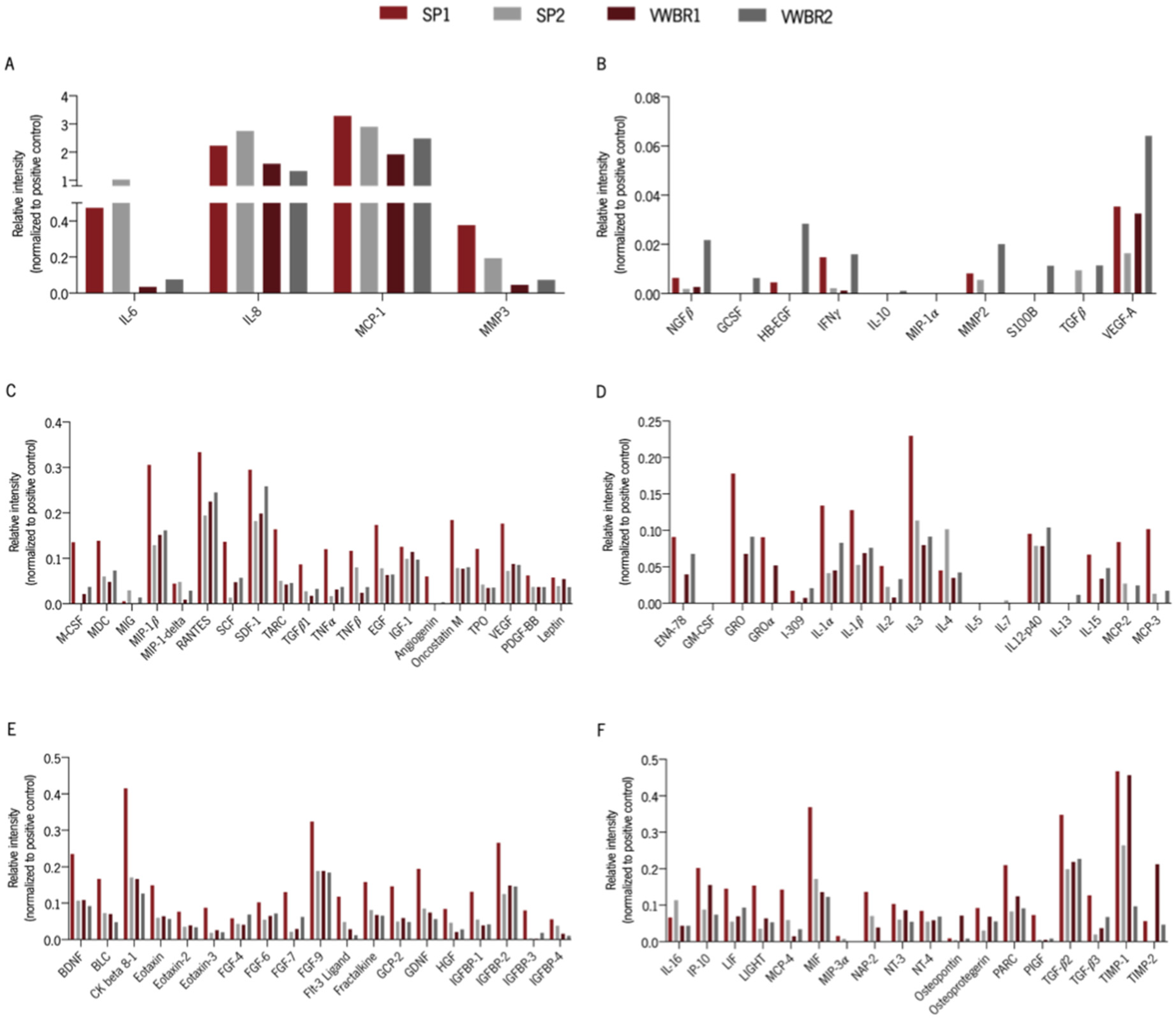
3.6. Cells Cultured in Both Systems Had Distinct Secretory Profiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bianco, P. “Mesenchymal” Stem Cells. Annu. Rev. Cell Dev. Biol. 2014, 30, 677–704. [Google Scholar] [CrossRef]
- Mushahary, D.; Spittler, A.; Kasper, C.; Weber, V.; Charwat, V. Isolation, Cultivation, and Characterization of Human Mesenchymal Stem Cells: HMSC. Cytometry 2018, 93, 19–31. [Google Scholar] [CrossRef]
- Harrell, C.R.; Fellabaum, C.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Molecular Mechanisms Responsible for Therapeutic Potential of Mesenchymal Stem Cell-Derived Secretome. Cells 2019, 8, 467. [Google Scholar] [CrossRef]
- Muhammad, S.A. Mesenchymal Stromal Cell Secretome as a Therapeutic Strategy for Traumatic Brain Injury. BioFactors 2019, 45, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Elia, C.A.; Tamborini, M.; Rasile, M.; Desiato, G.; Marchetti, S.; Swuec, P.; Mazzitelli, S.; Clemente, F.; Anselmo, A.; Matteoli, M.; et al. Intracerebral Injection of Extracellular Vesicles from Mesenchymal Stem Cells Exerts Reduced Aβ Plaque Burden in Early Stages of a Preclinical Model of Alzheimer’s Disease. Cells 2019, 8, 1059. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.G.; Carvalho, M.M.; Panchalingam, K.M.; Rodrigues, A.J.; Mendes-Pinheiro, B.; Anjo, S.; Manadas, B.; Behie, L.A.; Sousa, N.; Salgado, A.J. Impact of the Secretome of Human Mesenchymal Stem Cells on Brain Structure and Animal Behavior in a Rat Model of Parkinson’s Disease. Stem Cells Transl. Med. 2017, 6, 634–646. [Google Scholar] [CrossRef]
- Chudickova, M.; Vackova, I.; Machova Urdzikova, L.; Jancova, P.; Kekulova, K.; Rehorova, M.; Turnovcova, K.; Jendelova, P.; Kubinova, S. The Effect of Wharton Jelly-Derived Mesenchymal Stromal Cells and Their Conditioned Media in the Treatment of a Rat Spinal Cord Injury. IJMS 2019, 20, 4516. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.-E.; Lang, A.E. Parkinson Disease. Nat. Rev. Dis. Prim. 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Burbulla, L.F.; Song, P.; Mazzulli, J.R.; Zampese, E.; Wong, Y.C.; Jeon, S.; Santos, D.P.; Blanz, J.; Obermaier, C.D.; Strojny, C.; et al. Dopamine Oxidation Mediates Mitochondrial and Lysosomal Dysfunction in Parkinson’s Disease. Science 2017, 357, 1255–1261. [Google Scholar] [CrossRef]
- Wong, Y.C.; Krainc, D. α-Synuclein Toxicity in Neurodegeneration: Mechanism and Therapeutic Strategies. Nat. Med. 2017, 23, 1–13. [Google Scholar] [CrossRef]
- Villar-Piqué, A.; Lopes da Fonseca, T.; Outeiro, T.F. Structure, Function and Toxicity of Alpha-Synuclein: The Bermuda Triangle in Synucleinopathies. J. Neurochem. 2016, 139, 240–255. [Google Scholar] [CrossRef]
- Wilkins, A.; Kemp, K.; Ginty, M.; Hares, K.; Mallam, E.; Scolding, N. Human Bone Marrow-Derived Mesenchymal Stem Cells Secrete Brain-Derived Neurotrophic Factor Which Promotes Neuronal Survival in Vitro. Stem Cell Res. 2009, 3, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Baquet, Z.C.; Bickford, B.C.; Jones, K.R. Brain-Derived Neurotrophic Factor Is Required for the Establishment of the Proper Number of Dopaminergic Neurons in the Substantia Nigra Pars Compacta. J. Neurosci. 2005, 25, 6251–6259. [Google Scholar] [CrossRef]
- Mendes-Pinheiro, B.; Anjo, S.I.; Manadas, B.; Da Silva, J.D.; Marote, A.; Behie, L.A.; Teixeira, F.G.; Salgado, A.J. Bone Marrow Mesenchymal Stem Cells’ Secretome Exerts Neuroprotective Effects in a Parkinson’s Disease Rat Model. Front. Bioeng. Biotechnol. 2019, 7, 294. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.G.; Panchalingam, K.M.; Assunção-Silva, R.; Serra, S.C.; Mendes-Pinheiro, B.; Patrício, P.; Jung, S.; Anjo, S.I.; Manadas, B.; Pinto, L.; et al. Modulation of the Mesenchymal Stem Cell Secretome Using Computer-Controlled Bioreactors: Impact on Neuronal Cell Proliferation, Survival and Differentiation. Sci. Rep. 2016, 6, 27791. [Google Scholar] [CrossRef]
- Teixeira, F.G.; Vilaça-Faria, H.; Domingues, A.V.; Campos, J.; Salgado, A.J. Preclinical Comparison of Stem Cells Secretome and Levodopa Application in a 6-Hydroxydopamine Rat Model of Parkinson’s Disease. Cells 2020, 9, 315. [Google Scholar] [CrossRef]
- Das, R.; Roosloot, R.; van Pel, M.; Schepers, K.; Driessen, M.; Fibbe, W.E.; de Bruijn, J.D.; Roelofs, H. Preparing for Cell Culture Scale-out: Establishing Parity of Bioreactor- and Flask-Expanded Mesenchymal Stromal Cell Cultures. J. Transl. Med. 2019, 17, 241. [Google Scholar] [CrossRef]
- Pinto, D.S.; Ahsan, T.; Serra, J.; Fernandes-Platzgummer, A.; Cabral, J.M.S.; Silva, C.L. Modulation of the in Vitro Angiogenic Potential of Human Mesenchymal Stromal Cells from Different Tissue Sources. J. Cell. Physiol. 2020, 235, 7224–7238. [Google Scholar] [CrossRef]
- Croughan, M.S.; Giroux, D.; Fang, D.; Lee, B. Novel Single-Use Bioreactors for Scale-Up of Anchorage-Dependent Cell Manufacturing for Cell Therapies. In Stem Cell Manufacturing; Elsevier: Amsterdam, The Netherlands, 2016; pp. 105–139. ISBN 978-0-444-63265-4. [Google Scholar]
- Shukla, A.A.; Gottschalk, U. Single-Use Disposable Technologies for Biopharmaceutical Manufacturing. Trends Biotechnol. 2013, 31, 147–154. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, F.; Andrade, P.Z.; Boura, J.S.; Abecasis, M.M.; da Silva, C.L.; Cabral, J.M.S. Ex Vivo Expansion of Human Mesenchymal Stem Cells: A More Effective Cell Proliferation Kinetics and Metabolism under Hypoxia. J. Cell. Physiol. 2010, 223, 27–35. [Google Scholar] [CrossRef]
- de Almeida Fuzeta, M.; Bernardes, N.; Oliveira, F.D.; Costa, A.C.; Fernandes-Platzgummer, A.; Farinha, J.P.; Rodrigues, C.A.V.; Jung, S.; Tseng, R.-J.; Milligan, W.; et al. Scalable Production of Human Mesenchymal Stromal Cell-Derived Extracellular Vesicles Under Serum-/Xeno-Free Conditions in a Microcarrier-Based Bioreactor Culture System. Front. Cell Dev. Biol. 2020, 8, 553444. [Google Scholar] [CrossRef] [PubMed]
- Carmelo, J.G.; Fernandes-Platzgummer, A.; Diogo, M.M.; da Silva, C.L.; Cabral, J.M.S. A Xeno-Free Microcarrier-Based Stirred Culture System for the Scalable Expansion of Human Mesenchymal Stem/Stromal Cells Isolated from Bone Marrow and Adipose Tissue. Biotechnol. J. 2015, 10, 1235–1247. [Google Scholar] [CrossRef]
- Schop, D.; Janssen, F.W.; van Rijn, L.D.S.; Fernandes, H.; Bloem, R.M.; de Bruijn, J.D.; van Dijkhuizen-Radersma, R. Growth, Metabolism, and Growth Inhibitors of Mesenchymal Stem Cells. Tissue Eng. Part A 2009, 15, 1877–1886. [Google Scholar] [CrossRef] [PubMed]
- Serra, M.; Brito, C.; Sousa, M.F.Q.; Jensen, J.; Tostões, R.; Clemente, J.; Strehl, R.; Hyllner, J.; Carrondo, M.J.T.; Alves, P.M. Improving Expansion of Pluripotent Human Embryonic Stem Cells in Perfused Bioreactors through Oxygen Control. J. Biotechnol. 2010, 148, 208–215. [Google Scholar] [CrossRef] [PubMed]
- de Soure, A.M.; Fernandes-Platzgummer, A.; Moreira, F.; Lilaia, C.; Liu, S.-H.; Ku, C.-P.; Huang, Y.-F.; Milligan, W.; Cabral, J.M.S.; da Silva, C.L. Integrated Culture Platform Based on a Human Platelet Lysate Supplement for the Isolation and Scalable Manufacturing of Umbilical Cord Matrix-Derived Mesenchymal Stem/Stromal Cells. J. Tissue Eng. Regen. Med. 2017, 11, 1630–1640. [Google Scholar] [CrossRef]
- Teixeira, F.G.; Panchalingam, K.M.; Anjo, S.I.; Manadas, B.; Pereira, R.; Sousa, N.; Salgado, A.J.; Behie, L.A. Do Hypoxia/Normoxia Culturing Conditions Change the Neuroregulatory Profile of Wharton Jelly Mesenchymal Stem Cell Secretome? Stem Cell Res. Ther. 2015, 6, 133. [Google Scholar] [CrossRef]
- Teixeira, F.G.; Carvalho, M.M.; Neves-Carvalho, A.; Panchalingam, K.M.; Behie, L.A.; Pinto, L.; Sousa, N.; Salgado, A.J. Secretome of Mesenchymal Progenitors from the Umbilical Cord Acts as Modulator of Neural/Glial Proliferation and Differentiation. Stem Cell Rev. Rep. 2015, 11, 288–297. [Google Scholar] [CrossRef]
- Brenner, S. The Genetics of Caenorhabditis Elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Marques, C.R.; Pereira-Sousa, J.; Teixeira, F.G.; Sousa, R.A.; Teixeira-Castro, A.; Salgado, A.J. Mesenchymal Stem Cell Secretome Protects against Alpha-Synuclein-Induced Neurodegeneration in a Caenorhabditis Elegans Model of Parkinson’s Disease. Cytotherapy 2021. [Google Scholar] [CrossRef]
- dos Santos, F.; Campbell, A.; Fernandes-Platzgummer, A.; Andrade, P.Z.; Gimble, J.M.; Wen, Y.; Boucher, S.; Vemuri, M.C.; da Silva, C.L.; Cabral, J.M.S. A Xenogeneic-Free Bioreactor System for the Clinical-Scale Expansion of Human Mesenchymal Stem/Stromal Cells: Xeno-Free Production of Human MSC. Biotechnol. Bioeng. 2014, 111, 1116–1127. [Google Scholar] [CrossRef]
- Fernandes-Platzgummer, A.; Carmelo, J.G.; da Silva, C.L.; Cabral, J.M.S. Clinical-Grade Manufacturing of Therapeutic Human Mesenchymal Stem/Stromal Cells in Microcarrier-Based Culture Systems. In Mesenchymal Stem Cells; Gnecchi, M., Ed.; Springer: New York, NY, USA, 2016; Volume 1416, pp. 375–388. ISBN 978-1-4939-3582-6. [Google Scholar]
- Pinto, D.; Bandeiras, C.; Fuzeta, M.; Rodrigues, C.A.V.; Jung, S.; Hashimura, Y.; Tseng, R.; Milligan, W.; Lee, B.; Ferreira, F.C.; et al. Scalable Manufacturing of Human Mesenchymal Stromal Cells in the Vertical-Wheel Bioreactor System: An Experimental and Economic Approach. Biotechnol. J. 2019, 14, 1800716. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.A.; Silva, T.P.; Nogueira, D.E.; Fernandes, T.G.; Hashimura, Y.; Wesselschmidt, R.; Diogo, M.M.; Lee, B.; Cabral, J.M. Scalable Culture of Human Induced Pluripotent Cells on Microcarriers under Xeno-Free Conditions Using Single-Use Vertical-Wheel™ Bioreactors: Expansion of Human Induced Pluripotent Stem Cells in Vertical-Wheel Bioreactors. J. Chem. Technol. Biotechnol. 2018, 93, 3597–3606. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Hori, T.; Miyata, H.; Maehara, M.; Namba, T. Analysis of Proliferating Neuronal Progenitors and Immature Neurons in the Human Hippocampus Surgically Removed from Control and Epileptic Patients. Sci. Rep. 2019, 9, 18194. [Google Scholar] [CrossRef] [PubMed]
- Soltani, M.H.; Pichardo, R.; Song, Z.; Sangha, N.; Camacho, F.; Satyamoorthy, K.; Sangueza, O.P.; Setaluri, V. Microtubule-Associated Protein 2, a Marker of Neuronal Differentiation, Induces Mitotic Defects, Inhibits Growth of Melanoma Cells, and Predicts Metastatic Potential of Cutaneous Melanoma. Am. J. Pathol. 2005, 166, 1841–1850. [Google Scholar] [CrossRef] [PubMed]
- Hupfeld, J.; Gorr, I.H.; Schwald, C.; Beaucamp, N.; Wiechmann, K.; Kuentzer, K.; Huss, R.; Rieger, B.; Neubauer, M.; Wegmeyer, H. Modulation of Mesenchymal Stromal Cell Characteristics by Microcarrier Culture in Bioreactors. Biotechnol. Bioeng. 2014, 111, 2290–2302. [Google Scholar] [CrossRef]
- dos Santos, F.; Andrade, P.Z.; Abecasis, M.M.; Gimble, J.M.; Chase, L.G.; Campbell, A.M.; Boucher, S.; Vemuri, M.C.; da Silva, C.L.; Cabral, J.M.S. Toward a Clinical-Grade Expansion of Mesenchymal Stem Cells from Human Sources: A Microcarrier-Based Culture System Under Xeno-Free Conditions. Tissue Eng. Part C Methods 2011, 17, 1201–1210. [Google Scholar] [CrossRef]
- Mir, M.A. Introduction to Costimulation and Costimulatory Molecules. In Developing Costimulatory Molecules for Immunotherapy of Diseases; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1–43. ISBN 978-0-12-802585-7. [Google Scholar]
- Guyon, A. CXCL12 Chemokine and Its Receptors as Major Players in the Interactions between Immune and Nervous Systems. Front. Cell. Neurosci. 2014, 8, 65. [Google Scholar] [CrossRef]
- Sil, S.; Periyasamy, P.; Thangaraj, A.; Chivero, E.T.; Buch, S. PDGF/PDGFR Axis in the Neural Systems. Mol. Asp. Med. 2018, 62, 63–74. [Google Scholar] [CrossRef]
- Liu, X.-S.; Li, J.-F.; Wang, S.-S.; Wang, Y.-T.; Zhang, Y.-Z.; Yin, H.-L.; Geng, S.; Gong, H.-C.; Han, B.; Wang, Y.-L. Human Umbilical Cord Mesenchymal Stem Cells Infected with Adenovirus Expressing HGF Promote Regeneration of Damaged Neuron Cells in a Parkinson’s Disease Model. Biomed. Res. Int. 2014, 2014, 909657. [Google Scholar] [CrossRef]
- Allen, S.J.; Watson, J.J.; Shoemark, D.K.; Barua, N.U.; Patel, N.K. GDNF, NGF and BDNF as Therapeutic Options for Neurodegeneration. Pharmacol. Ther. 2013, 138, 155–175. [Google Scholar] [CrossRef]
- Bhattarai, P.; Thomas, A.K.; Cosacak, M.I.; Papadimitriou, C.; Mashkaryan, V.; Froc, C.; Reinhardt, S.; Kurth, T.; Dahl, A.; Zhang, Y.; et al. IL4/STAT6 Signaling Activates Neural Stem Cell Proliferation and Neurogenesis upon Amyloid-Β42 Aggregation in Adult Zebrafish Brain. Cell Rep. 2016, 17, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.H.; Kim, H.N.; Park, H.J.; Shin, J.Y.; Kim, D.Y.; Lee, P.H. The Cleavage Effect of Mesenchymal Stem Cell and Its Derived Matrix Metalloproteinase-2 on Extracellular α-Synuclein Aggregates in Parkinsonian Models. Stem Cells Transl. Med. 2017, 6, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Casati, S.; Giannasi, C.; Niada, S.; Della Morte, E.; Orioli, M.; Brini, A.T. Lipidomics of Cell Secretome Combined with the Study of Selected Bioactive Lipids in an In Vitro Model of Osteoarthritis. Stem Cells Transl. Med. 2022, 11, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Jossen, V.; Pörtner, R.; Kaiser, S.C.; Kraume, M.; Eibl, D.; Eibl, R. Mass Production of Mesenchymal Stem Cells—Impact of Bioreactor Design and Flow Conditions on Proliferation and Differentiation. In Cells and Biomaterials in Regenerative Medicine; Eberli, D., Ed.; InTech: London, UK, 2014; ISBN 978-953-51-1731-5. [Google Scholar]
- Ismadi, M.-Z.; Hourigan, K.; Fouras, A. Experimental Characterisation of Fluid Mechanics in a Spinner Flask Bioreactor. Processes 2014, 2, 753–772. [Google Scholar] [CrossRef]
- Diaz, M.F.; Evans, S.M.; Olson, S.D.; Cox, C.S.; Wenzel, P.L. A Co-Culture Assay to Determine Efficacy of TNF-α Suppression by Biomechanically Induced Human Bone Marrow Mesenchymal Stem Cells. Bio-Protocol 2017, 7, e2513. [Google Scholar] [CrossRef]
- Diaz, M.F.; Vaidya, A.B.; Evans, S.M.; Lee, H.J.; Aertker, B.M.; Alexander, A.J.; Price, K.M.; Ozuna, J.A.; Liao, G.P.; Aroom, K.R.; et al. Biomechanical Forces Promote Immune Regulatory Function of Bone Marrow Mesenchymal Stromal Cells. Stem Cells 2017, 35, 1259–1272. [Google Scholar] [CrossRef]
- Jones, S.A.; Jenkins, B.J. Recent Insights into Targeting the IL-6 Cytokine Family in Inflammatory Diseases and Cancer. Nat. Rev. Immunol. 2018, 18, 773–789. [Google Scholar] [CrossRef]
- Zhang, J.-M.; An, J. Cytokines, Inflammation, and Pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Swamynathan, P.; Venugopal, P.; Kannan, S.; Thej, C.; Kolkundar, U.; Bhagwat, S.; Ta, M.; Majumdar, A.S.; Balasubramanian, S. Are Serum-Free and Xeno-Free Culture Conditions Ideal for Large Scale Clinical Grade Expansion of Wharton’s Jelly Derived Mesenchymal Stem Cells? A Comparative Study. Stem Cell Res. Ther. 2014, 5, 88. [Google Scholar] [CrossRef]
- Yoshida, K.; Nakashima, A.; Doi, S.; Ueno, T.; Okubo, T.; Kawano, K.; Kanawa, M.; Kato, Y.; Higashi, Y.; Masaki, T. Serum-Free Medium Enhances the Immunosuppressive and Antifibrotic Abilities of Mesenchymal Stem Cells Utilized in Experimental Renal Fibrosis. Stem Cells Transl. Med. 2018, 7, 893–905. [Google Scholar] [CrossRef]
- Abdelrazik, H.; Spaggiari, G.M.; Chiossone, L.; Moretta, L. Mesenchymal Stem Cells Expanded in Human Platelet Lysate Display a Decreased Inhibitory Capacity on T- and NK-Cell Proliferation and Function. Eur. J. Immunol. 2011, 41, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Oikonomopoulos, A.; van Deen, W.K.; Manansala, A.-R.; Lacey, P.N.; Tomakili, T.A.; Ziman, A.; Hommes, D.W. Optimization of Human Mesenchymal Stem Cell Manufacturing: The Effects of Animal/Xeno-Free Media. Sci. Rep. 2015, 5, 16570. [Google Scholar] [CrossRef] [PubMed]
- Weil, B.R.; Abarbanell, A.M.; Herrmann, J.L.; Wang, Y.; Meldrum, D.R. High Glucose Concentration in Cell Culture Medium Does Not Acutely Affect Human Mesenchymal Stem Cell Growth Factor Production or Proliferation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1735–R1743. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dynamic System | Culture Medium Used for Cell Expansion | Culture Medium Used for Conditioning | Agitation Rate (rpm) | Cell Adhesion Efficiency (%) | Specific Growth Rate (Day−1) | Fold Expansion | Doubling Time (Day) |
|---|---|---|---|---|---|---|---|
| VWBR1 | DMEM 5% UltraGRO™-PURE | AlphaMEM | 30 | 76 | 0.31 | 4.19 | 2.24 |
| VWBR2 | DMEM 5% UltraGRO™-PURE | Neurobasal-A | 25 | 31 | 0.24 | 2.58 | 2.90 |
| SP1 | StemPro MSC SFM | AlphaMEM | 40 | 48 | 0.36 | 2.93 | 1.92 |
| SP2 | StemPro MSC SFM | Neurobasal-A | 40 | 55 | 0.31 | 2.45 | 2.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, C.R.; Fuzeta, M.d.A.; dos Santos Cunha, R.M.; Pereira-Sousa, J.; Silva, D.; Campos, J.; Teixeira-Castro, A.; Sousa, R.A.; Fernandes-Platzgummer, A.; da Silva, C.L.; et al. Neurodifferentiation and Neuroprotection Potential of Mesenchymal Stromal Cell-Derived Secretome Produced in Different Dynamic Systems. Biomedicines 2023, 11, 1240. https://doi.org/10.3390/biomedicines11051240
Marques CR, Fuzeta MdA, dos Santos Cunha RM, Pereira-Sousa J, Silva D, Campos J, Teixeira-Castro A, Sousa RA, Fernandes-Platzgummer A, da Silva CL, et al. Neurodifferentiation and Neuroprotection Potential of Mesenchymal Stromal Cell-Derived Secretome Produced in Different Dynamic Systems. Biomedicines. 2023; 11(5):1240. https://doi.org/10.3390/biomedicines11051240
Chicago/Turabian StyleMarques, Cláudia Raquel, Miguel de Almeida Fuzeta, Raquel Medina dos Santos Cunha, Joana Pereira-Sousa, Deolinda Silva, Jonas Campos, Andreia Teixeira-Castro, Rui Amandi Sousa, Ana Fernandes-Platzgummer, Cláudia L. da Silva, and et al. 2023. "Neurodifferentiation and Neuroprotection Potential of Mesenchymal Stromal Cell-Derived Secretome Produced in Different Dynamic Systems" Biomedicines 11, no. 5: 1240. https://doi.org/10.3390/biomedicines11051240