
Disturbances in Switching between Canonical and Non-Canonical Wnt Signaling Characterize Developing and Postnatal Kidneys of Dab1−/− (yotari) Mice
 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Sample Collection
2.3. Immunofluorescence Staining
2.4. Data Acquisition and Analysis
2.5. Statistical Analyses
3. Results
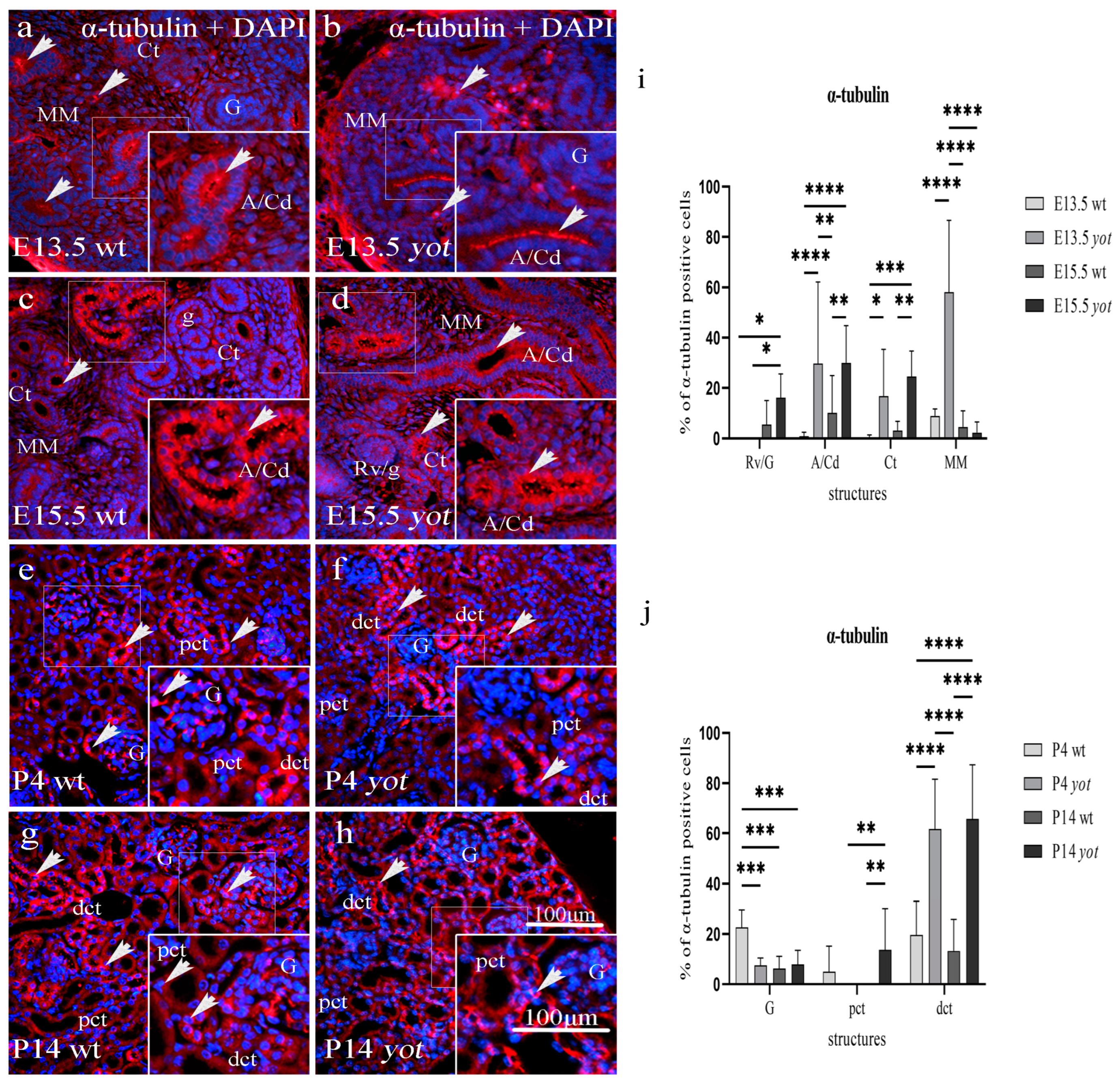
3.1. Acetylated α-Tubulin Immunoexpression
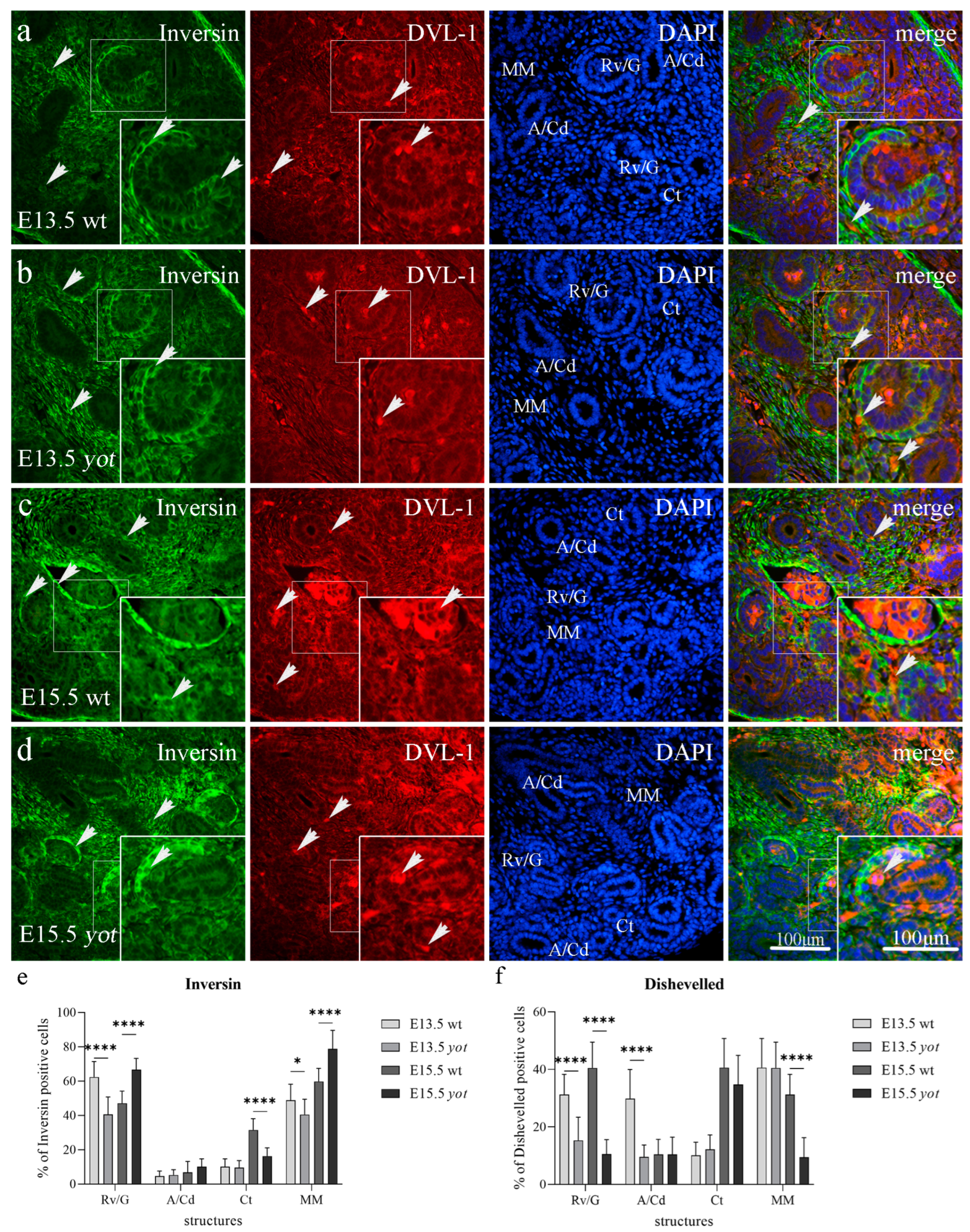
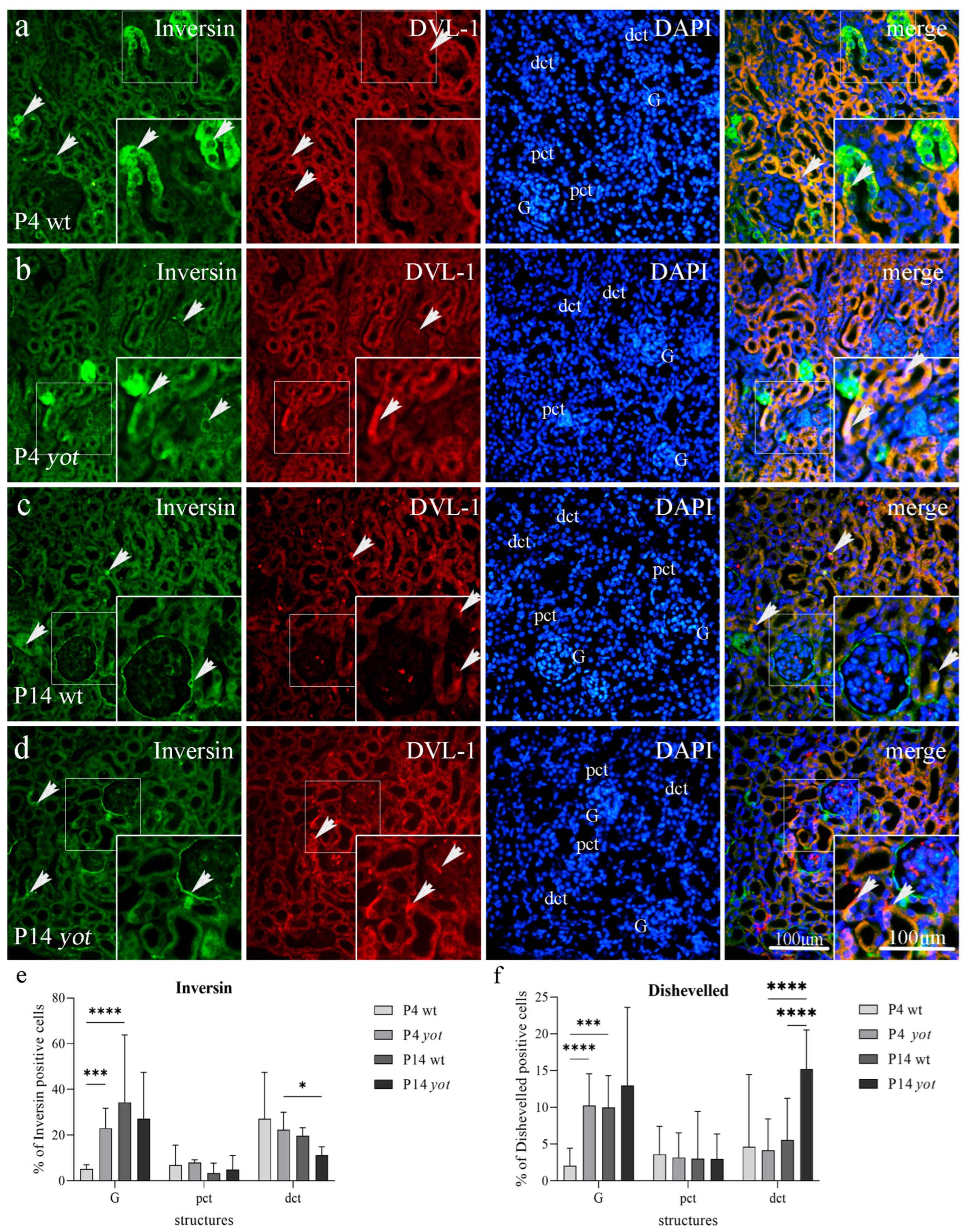
3.2. Double Immunofluorescence Staining to Inversin and DVL-1 in Developing and Postnatal Wild-Type and Yotari Mouse Kidneys
3.2.1. Inversin and DVL-1 Expression in Developing Kidney of Wild-Type and Yotari Mice
3.2.2. Inversin and DVL-1 Expression in Postnatal Kidney of Wild-Type and Yotari Mice
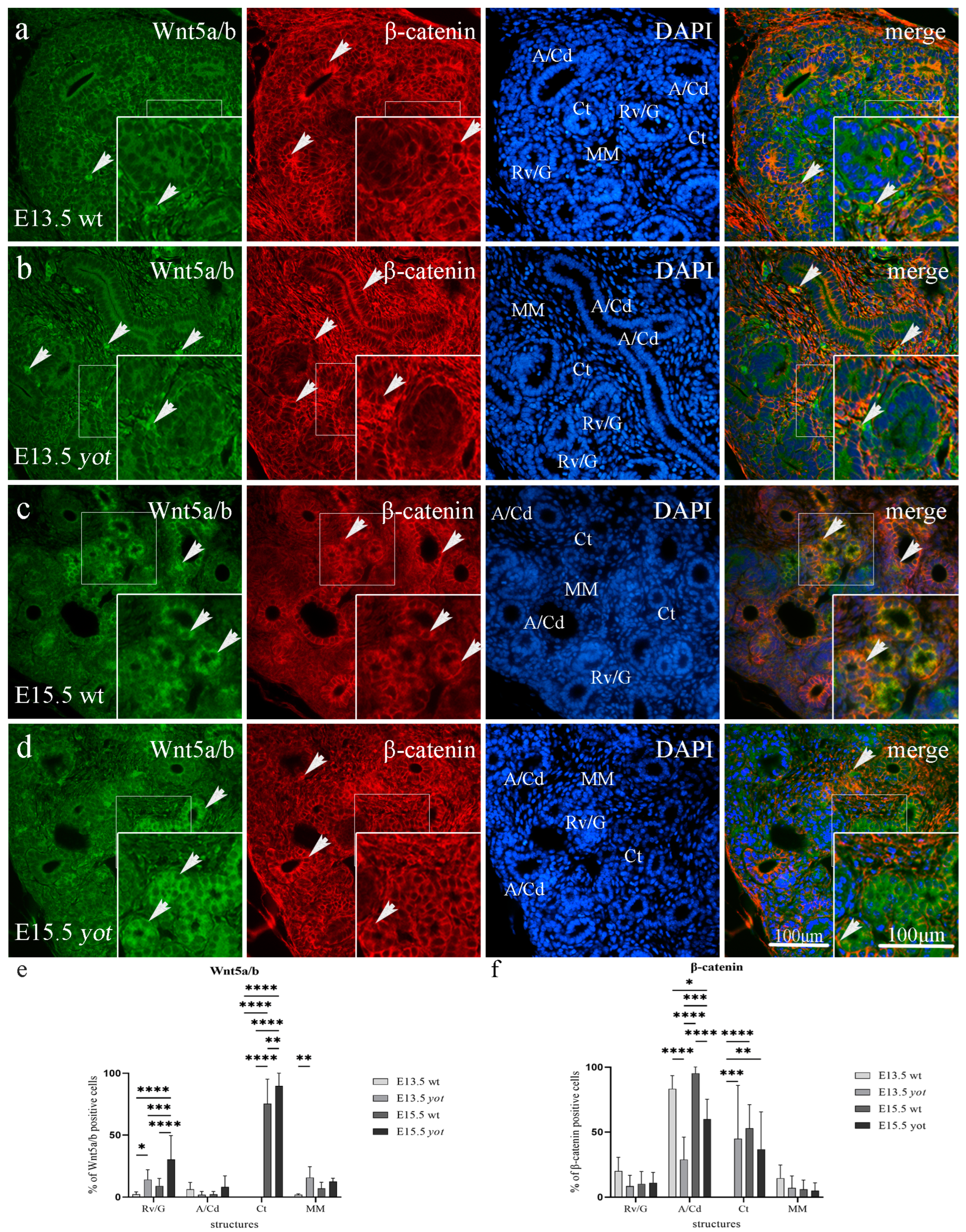
3.3. Double Immunofluorescence Staining to Wnt5a/b and β-Catenin in Developing and Postnatal Wild-Type and Yotari Mouse Kidneys
3.3.1. Wnt5a/b and β-Catenin Expression in Developing Kidney of Wild-Type and Yotari Mice
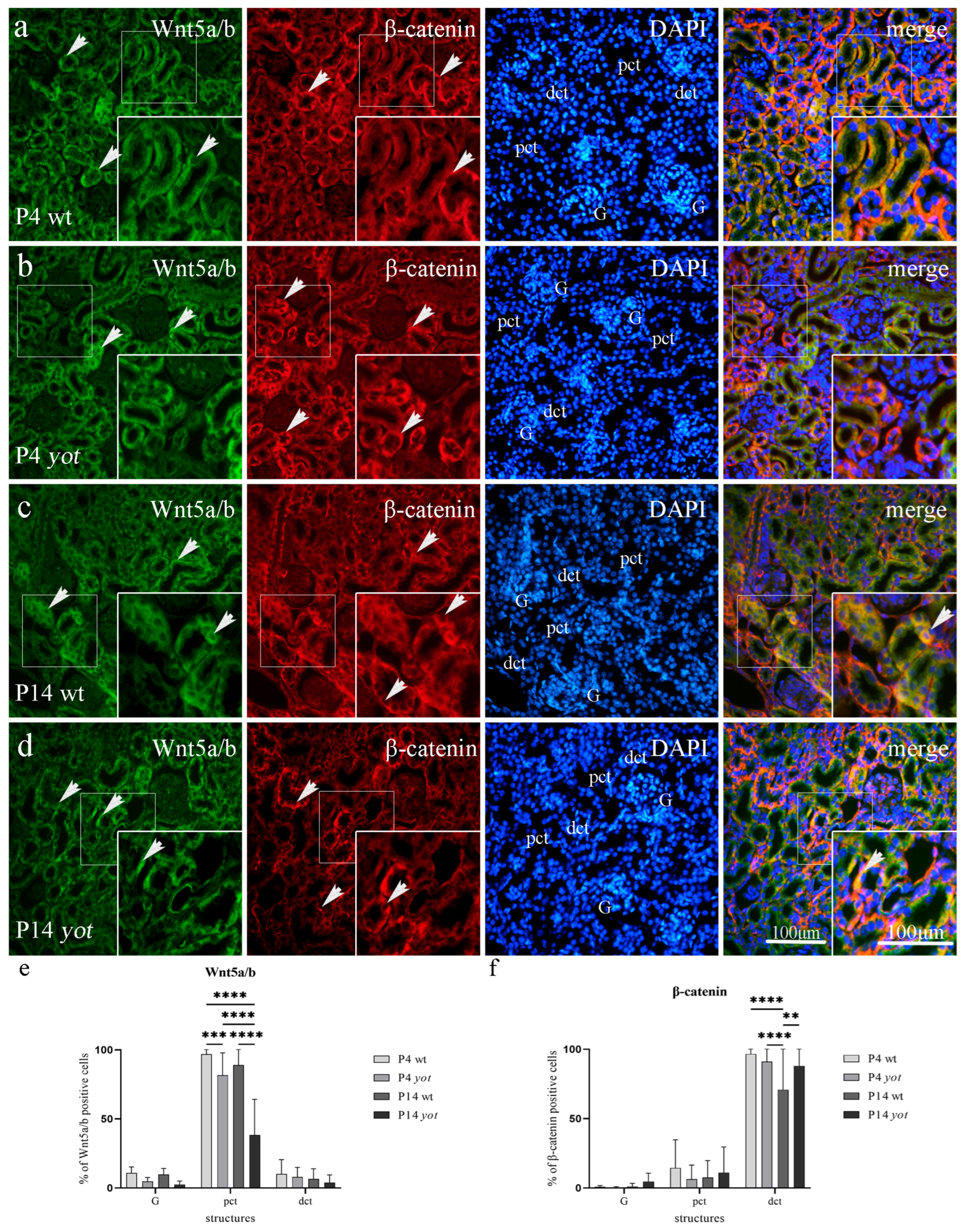
3.3.2. Wnt5a/b and β-Catenin Expression in Postnatal Kidney of Wild-Type and Yotari Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saxen, L.; Sariola, H. Early organogenesis of the kidney. Pediatr. Nephrol. 1987, 1, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Georgas, K.; Rumballe, B.; Valerius, M.T.; Chiu, H.S.; Thiagarajan, R.D.; Lesieur, E.; Aronow, B.J.; Brunskill, E.W.; Combes, A.N.; Tang, D.; et al. Analysis of early nephron patterning reveals a role for distal RV proliferation in fusion to the ureteric tip via a cap mesenchyme-derived connecting segment. Dev. Biol. 2009, 332, 273–286. [Google Scholar] [CrossRef]
- Arndt, M.; Bigelow, B. The impact of individual and contextual factors on nursing care and the implementation of AHCPR practice guidelines. Med. Care Rev. 1994, 51, 61–82. [Google Scholar] [CrossRef]
- Kao, R.M.; Vasilyev, A.; Miyawaki, A.; Drummond, I.A.; McMahon, A.P. Invasion of distal nephron precursors associates with tubular interconnection during nephrogenesis. J. Am. Soc. Nephrol. JASN 2012, 23, 1682–1690. [Google Scholar] [CrossRef]
- Pei, D.; Shu, X.; Gassama-Diagne, A.; Thiery, J.P. Mesenchymal-epithelial transition in development and reprogramming. Nat. Cell Biol. 2019, 21, 44–53. [Google Scholar] [CrossRef]
- Loane, M.; Dolk, H.; Kelly, A.; Teljeur, C.; Greenlees, R.; Densem, J. Paper 4: EUROCAT statistical monitoring: Identification and investigation of ten year trends of congenital anomalies in Europe. Birth Defects Res. Part A Clin. Mol. Teratol. 2011, 91 (Suppl. S1), S31–S43. [Google Scholar] [CrossRef] [PubMed]
- Sanna-Cherchi, S.; Sampogna, R.V.; Papeta, N.; Burgess, K.E.; Nees, S.N.; Perry, B.J.; Choi, M.; Bodria, M.; Liu, Y.; Weng, P.L.; et al. Mutations in DSTYK and dominant urinary tract malformations. N. Engl. J. Med. 2013, 369, 621–629. [Google Scholar] [CrossRef]
- Dlugosz, P.; Teufl, M.; Schwab, M.; Kohl, K.E.; Nimpf, J. Disabled 1 Is Part of a Signaling Pathway Activated by Epidermal Growth Factor Receptor. Int. J. Mol. Sci. 2021, 22, 1745. [Google Scholar] [CrossRef] [PubMed]
- Racetin, A.; Filipovic, N.; Lozic, M.; Ogata, M.; Gudelj Ensor, L.; Kelam, N.; Kovacevic, P.; Watanabe, K.; Katsuyama, Y.; Saraga-Babic, M.; et al. A Homozygous Dab1(−/−) Is a Potential Novel Cause of Autosomal Recessive Congenital Anomalies of the Mice Kidney and Urinary Tract. Biomolecules 2021, 11, 609. [Google Scholar] [CrossRef]
- Gao, Z.; Godbout, R. Reelin-Disabled-1 signaling in neuronal migration: Splicing takes the stage. Cell. Mol. Life Sci. CMLS 2013, 70, 2319–2329. [Google Scholar] [CrossRef]
- Nlandu-Khodo, S.; Osaki, Y.; Scarfe, L.; Yang, H.; Phillips-Mignemi, M.; Tonello, J.; Saito-Diaz, K.; Neelisetty, S.; Ivanova, A.; Huffstater, T.; et al. Tubular beta-catenin and FoxO3 interactions protect in chronic kidney disease. JCI Insight 2020, 5, e135454. [Google Scholar] [CrossRef]
- Kim, K.; Lu, Z.; Hay, E.D. Direct evidence for a role of beta-catenin/LEF-1 signaling pathway in induction of EMT. Cell Biol. Int. 2002, 26, 463–476. [Google Scholar] [CrossRef]
- Sugimura, R.; Li, L. Noncanonical Wnt signaling in vertebrate development, stem cells, and diseases. Birth Defects Res. Part C Embryo Today Rev. 2010, 90, 243–256. [Google Scholar] [CrossRef]
- Veeman, M.T.; Axelrod, J.D.; Moon, R.T. A second canon. Functions and mechanisms of beta-catenin-independent Wnt signaling. Dev. Cell 2003, 5, 367–377. [Google Scholar] [CrossRef]
- Wood, A.L.; O’Brien, S.J.; Geddes, A.M. Initial management of suspected meningococcal infection. Chief medical officer’s guidelines are right. BMJ 1994, 309, 1661–1662. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Jung, J.K.; Im, S.S.; Lee, S.R.; Jang, B.C.; Park, K.M.; Kim, J.I. Deficiency of primary cilia in kidney epithelial cells induces epithelial to mesenchymal transition. Biochem. Biophys. Res. Commun. 2018, 496, 450–454. [Google Scholar] [CrossRef]
- Wheatley, D.N.; Wang, A.M.; Strugnell, G.E. Expression of primary cilia in mammalian cells. Cell Biol. Int. 1996, 20, 73–81. [Google Scholar] [CrossRef]
- Liu, Y. Epithelial to mesenchymal transition in renal fibrogenesis: Pathologic significance, molecular mechanism, and therapeutic intervention. J. Am. Soc. Nephrol. JASN 2004, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Liu, Y.; Zhu, B.; Ding, K.; Yao, T.P.; Chen, F.; Zhan, L.; Xu, P.; Ehrlich, M.; Liang, T.; et al. Loss of alpha-Tubulin Acetylation Is Associated with TGF-beta-induced Epithelial-Mesenchymal Transition. J. Biol. Chem. 2016, 291, 5396–5405. [Google Scholar] [CrossRef]
- Solic, I.; Racetin, A.; Filipovic, N.; Mardesic, S.; Bocina, I.; Galesic-Ljubanovic, D.; Glavina Durdov, M.; Saraga-Babic, M.; Vukojevic, K. Expression Pattern of alpha-Tubulin, Inversin and Its Target Dishevelled-1 and Morphology of Primary Cilia in Normal Human Kidney Development and Diseases. Int. J. Mol. Sci. 2021, 22, 3500. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, T.; Saijoh, Y.; Tsuchiya, K.; Shirayoshi, Y.; Takai, S.; Taya, C.; Yonekawa, H.; Yamada, K.; Nihei, H.; Nakatsuji, N.; et al. Cloning of inv, a gene that controls left/right asymmetry and kidney development. Nature 1998, 395, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Otto, E.A.; Schermer, B.; Obara, T.; O’Toole, J.F.; Hiller, K.S.; Mueller, A.M.; Ruf, R.G.; Hoefele, J.; Beekmann, F.; Landau, D.; et al. Mutations in INVS encoding inversin cause nephronophthisis type 2, linking renal cystic disease to the function of primary cilia and left-right axis determination. Nat. Genet. 2003, 34, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Ocbina, P.J.; Tuson, M.; Anderson, K.V. Primary cilia are not required for normal canonical Wnt signaling in the mouse embryo. PLoS ONE 2009, 4, e6839. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Gleeson, J.G. Cystic kidney disease: The role of Wnt signaling. Trends Mol. Med. 2010, 16, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Jonassen, J.A.; San Agustin, J.; Follit, J.A.; Pazour, G.J. Deletion of IFT20 in the mouse kidney causes misorientation of the mitotic spindle and cystic kidney disease. J. Cell Biol. 2008, 183, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Nishita, M.; Qiao, S.; Miyamoto, M.; Okinaka, Y.; Yamada, M.; Hashimoto, R.; Iijima, K.; Otani, H.; Hartmann, C.; Nishinakamura, R.; et al. Role of Wnt5a-Ror2 signaling in morphogenesis of the metanephric mesenchyme during ureteric budding. Mol. Cell. Biol. 2014, 34, 3096–3105. [Google Scholar] [CrossRef] [PubMed]
- Luyten, A.; Su, X.; Gondela, S.; Chen, Y.; Rompani, S.; Takakura, A.; Zhou, J. Aberrant regulation of planar cell polarity in polycystic kidney disease. J. Am. Soc. Nephrol. JASN 2010, 21, 1521–1532. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Valerius, M.T.; McMahon, A.P. Wnt/beta-catenin signaling regulates nephron induction during mouse kidney development. Development 2007, 134, 2533–2539. [Google Scholar] [CrossRef]
- Hu, L.; Ding, M.; He, W. Emerging Therapeutic Strategies for Attenuating Tubular EMT and Kidney Fibrosis by Targeting Wnt/beta-Catenin Signaling. Front. Pharmacol. 2021, 12, 830340. [Google Scholar] [CrossRef]
- Tan, R.J.; Zhou, D.; Zhou, L.; Liu, Y. Wnt/beta-catenin signaling and kidney fibrosis. Kidney Int. Suppl. 2014, 4, 84–90. [Google Scholar] [CrossRef]
- Ye, B.; Ge, Y.; Perens, G.; Hong, L.; Xu, H.; Fishbein, M.C.; Li, F. Canonical Wnt/beta-catenin signaling in epicardial fibrosis of failed pediatric heart allografts with diastolic dysfunction. Cardiovasc. Pathol. Off. J. Soc. Cardiovasc. Pathol. 2013, 22, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Arimitsu, N.; Mizukami, Y.; Shimizu, J.; Takai, K.; Suzuki, T.; Suzuki, N. Defective Reelin/Dab1 signaling pathways associated with disturbed hippocampus development of homozygous yotari mice. Mol. Cell. Neurosci. 2021, 112, 103614. [Google Scholar] [CrossRef] [PubMed]
- Yoneshima, H.; Nagata, E.; Matsumoto, M.; Yamada, M.; Nakajima, K.; Miyata, T.; Ogawa, M.; Mikoshiba, K. A novel neurological mutant mouse, yotari, which exhibits reeler-like phenotype but expresses CR-50 antigen/reelin. Neurosci. Res. 1997, 29, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Gotay, A. Establishing security awareness guidelines in a hospital setting. J. Healthc. Prot. Manag. Publ. Int. Assoc. Hosp. Secur. 1994, 10, 80–83. Available online: http://www.ncbi.nlm.nih.gov/pubmed/10136049 (accessed on 24 January 2023).
- Toubas, J.; Beck, S.; Pageaud, A.L.; Huby, A.C.; Mael-Ainin, M.; Dussaule, J.C.; Chatziantoniou, C.; Chadjichristos, C.E. Alteration of connexin expression is an early signal for chronic kidney disease. Am. J. Physiol. Ren. Physiol. 2011, 301, F24–F32. [Google Scholar] [CrossRef]
- Wu, J.C.Y. Wu X, Ying Y, Tai Y, He, M. Transcardiac Perfusion of the Mouse for Brain Tissue Dissection and Fixation. Biol. Protoc. 2021, 11, e3988. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ; U.S. National Institutes of Health: Bethesda, ML, USA, 1997–2018. Available online: https://imagej.nih.gov/ij/ (accessed on 5 December 2022).
- Cicchetti, D.V. Guidelines, Criteria and Rules of Thumb for Evaluating Normed and Standardized Assessment Instruments in Psychology. Psychol. Assess. 1994, 6, 284–290. [Google Scholar] [CrossRef]
- Nicolaou, N.; Renkema, K.Y.; Bongers, E.M.; Giles, R.H.; Knoers, N.V. Genetic, environmental, and epigenetic factors involved in CAKUT. Nat. Rev. Nephrol. 2015, 11, 720–731. [Google Scholar] [CrossRef]
- Warady, B.A.; Chadha, V. Chronic kidney disease in children: The global perspective. Pediatr. Nephrol. 2007, 22, 1999–2009. [Google Scholar] [CrossRef]
- Saito, S.; Tampe, B.; Muller, G.A.; Zeisberg, M. Primary cilia modulate balance of canonical and non-canonical Wnt signaling responses in the injured kidney. Fibrogenes. Tissue Repair 2015, 8, 6. [Google Scholar] [CrossRef]
- Iglesias, D.M.; Hueber, P.A.; Chu, L.; Campbell, R.; Patenaude, A.M.; Dziarmaga, A.J.; Quinlan, J.; Mohamed, O.; Dufort, D.; Goodyer, P.R. Canonical WNT signaling during kidney development. Am. J. Physiol. Ren. Physiol. 2007, 293, F494–F500. [Google Scholar] [CrossRef]
- Gallegos, T.F.; Kouznetsova, V.; Kudlicka, K.; Sweeney, D.E.; Bush, K.T.; Willert, K.; Farquhar, M.G.; Nigam, S.K. A protein kinase A and Wnt-dependent network regulating an intermediate stage in epithelial tubulogenesis during kidney development. Dev. Biol. 2012, 364, 11–21. [Google Scholar] [CrossRef]
- Carroll, T.J.; Das, A. Planar cell polarity in kidney development and disease. Organogenesis 2011, 7, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Li, Y.; Lin, L.; Zhou, L.; Igarashi, P.; Liu, Y. Tubule-specific ablation of endogenous beta-catenin aggravates acute kidney injury in mice. Kidney Int. 2012, 82, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Bellavia, S.; Dahan, K.; Terryn, S.; Cosyns, J.P.; Devuyst, O.; Pirson, Y. A homozygous mutation in INVS causing juvenile nephronophthisis with abnormal reactivity of the Wnt/beta-catenin pathway. Nephrol. Dial. Transplant. 2010, 25, 4097–4102. [Google Scholar] [CrossRef] [PubMed]
- Schueler, M.; Braun, D.A.; Chandrasekar, G.; Gee, H.Y.; Klasson, T.D.; Halbritter, J.; Bieder, A.; Porath, J.D.; Airik, R.; Zhou, W.; et al. DCDC2 mutations cause a renal-hepatic ciliopathy by disrupting Wnt signaling. Am. J. Hum. Genet. 2015, 96, 81–92. [Google Scholar] [CrossRef]
- Simons, M.; Gloy, J.; Ganner, A.; Bullerkotte, A.; Bashkurov, M.; Kronig, C.; Schermer, B.; Benzing, T.; Cabello, O.A.; Jenny, A.; et al. Inversin, the gene product mutated in nephronophthisis type II, functions as a molecular switch between Wnt signaling pathways. Nat. Genet. 2005, 37, 537–543. [Google Scholar] [CrossRef]
- Hertig, A.; Verine, J.; Mougenot, B.; Jouanneau, C.; Ouali, N.; Sebe, P.; Glotz, D.; Ancel, P.Y.; Rondeau, E.; Xu-Dubois, Y.C. Risk factors for early epithelial to mesenchymal transition in renal grafts. Am. J. Transplant. Off. J. Am. Soc. Transplant. Am. Soc. Transpl. Surg. 2006, 6, 2937–2946. [Google Scholar] [CrossRef]
- Kawakami, T.; Ren, S.; Duffield, J.S. Wnt signalling in kidney diseases: Dual roles in renal injury and repair. J. Pathol. 2013, 229, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Xiao, A.; Choi, S.Y.; Kan, Q.; Zhou, W.; Chacon-Heszele, M.F.; Ryu, Y.K.; McKenna, S.; Zuo, X.; Kuruvilla, R.; et al. Wnt5a is necessary for normal kidney development in zebrafish and mice. Nephron. Exp. Nephrol. 2014, 128, 80–88. [Google Scholar] [CrossRef]
- Qian, D.; Jones, C.; Rzadzinska, A.; Mark, S.; Zhang, X.; Steel, K.P.; Dai, X.; Chen, P. Wnt5a functions in planar cell polarity regulation in mice. Dev. Biol. 2007, 306, 121–133. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Catalog Number | Host | Dilution | Source | |
|---|---|---|---|---|---|
| Primary | Anti-acetyl-α-tubulin (Lys40) (6-11B-1) | 12152S | Mouse | 1:500 | Cell Signaling Technology (CST), (Danvers, MA, USA) |
| Anti-inversin | ab65187 | Rabbit | 1:100 | Abcam (Cambridge, UK) | |
| Anti-dishevelled-1 (3F12) | sc-8025 | Mouse | 1:50 | Santa Cruz Biotechnology (Dallas, TX, USA) | |
| Anti-Wnt5a/b (C27E8) | 2530S | Rabbit | 1:100 | Cell Signaling Technology (CST), (Danvers, MA, USA) | |
| Anti-β-catenin (L54E2) | 2677S | Mouse | 1:200 | Cell Signaling Technology (CST), (Danvers, MA, USA) | |
| Secondary | Anti-Rabbit IgG, Alexa Fluor® 488 | 711-545-152 | Donkey | 1:300 | Jackson Immuno Research Laboratories, Inc., (Baltimore, PA, USA) |
| Anti-Mouse IgG, Rhodamine Red™-X | 715-295-151 | Donkey | 1:300 | Jackson Immuno Research Laboratories, Inc., (Baltimore, PA, USA) |
| Embryonic Day (E) | Animal | Structure | Antibody | ||||
|---|---|---|---|---|---|---|---|
| α-Tubulin | Inversin | DVL-1 | Wnt5a/b | β-Catenin | |||
| E13.5 | wild-type | Rv/G | − | ++ | + | − | + |
| A/Cd | ++ | − | −/+ | + | +++ | ||
| Ct | −/+ | − | − | − | + | ||
| MM | + | ++ | + | ++ | ++ | ||
| yotari | Rv/G | − | ++ | + | + | + | |
| A/Cd | +++ | + | − | − | +++ | ||
| Ct | + | −/+ | −/+ | − | +++ | ||
| MM | ++ | +++ | ++ | ++ | ++ | ||
| E15.5 | wild-type | Rv/G | −/+ | ++ | ++ | + | −/+ |
| A/Cd | +++ | −/+ | −/+ | − | ++ | ||
| Ct | + | + | − | ++ | ++ | ||
| MM | + | +++ | + | + | + | ||
| yotari | Rv/G | + | ++ | ++ | + | − | |
| A/Cd | +++ | − | + | + | +++ | ||
| Ct | ++ | + | −/+ | ++ | ++ | ||
| MM | + | +++ | ++ | + | + | ||
| Postnatal Day (P) | Animal | Structure | Antibody | ||||
|---|---|---|---|---|---|---|---|
| α-Tubulin | Inversin | DVL-1 | Wnt5a/b | β-Catenin | |||
| P4 | wild-type | G | + | −/+ | + | −/+ | −/+ |
| pct | + | −/+ | − | ++ | + | ||
| dct | ++ | +++ | ++ | − | ++ | ||
| yotari | G | −/+ | + | + | − | − | |
| pct | + | − | − | +++ | + | ||
| dct | ++ | +++ | ++ | − | +++ | ||
| P14 | wild-type | G | + | ++ | ++ | −/+ | − |
| pct | − | − | + | ++ | −/+ | ||
| dct | + | + | + | − | ++ | ||
| yotari | G | −/+ | ++ | ++ | + | + | |
| pct | + | −/+ | + | +++ | + | ||
| dct | ++ | + | + | + | +++ | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perutina, I.; Kelam, N.; Maglica, M.; Racetin, A.; Ogorevc, M.; Filipović, N.; Katsuyama, Y.; Mišković, J.; Vukojević, K. Disturbances in Switching between Canonical and Non-Canonical Wnt Signaling Characterize Developing and Postnatal Kidneys of Dab1−/− (yotari) Mice. Biomedicines 2023, 11, 1321. https://doi.org/10.3390/biomedicines11051321
Perutina I, Kelam N, Maglica M, Racetin A, Ogorevc M, Filipović N, Katsuyama Y, Mišković J, Vukojević K. Disturbances in Switching between Canonical and Non-Canonical Wnt Signaling Characterize Developing and Postnatal Kidneys of Dab1−/− (yotari) Mice. Biomedicines. 2023; 11(5):1321. https://doi.org/10.3390/biomedicines11051321
Chicago/Turabian StylePerutina, Ilija, Nela Kelam, Mirko Maglica, Anita Racetin, Marin Ogorevc, Natalija Filipović, Yu Katsuyama, Josip Mišković, and Katarina Vukojević. 2023. "Disturbances in Switching between Canonical and Non-Canonical Wnt Signaling Characterize Developing and Postnatal Kidneys of Dab1−/− (yotari) Mice" Biomedicines 11, no. 5: 1321. https://doi.org/10.3390/biomedicines11051321