
The Role of Genetic Polymorphisms in Differentiated Thyroid Cancer: A 2023 Update

Abstract
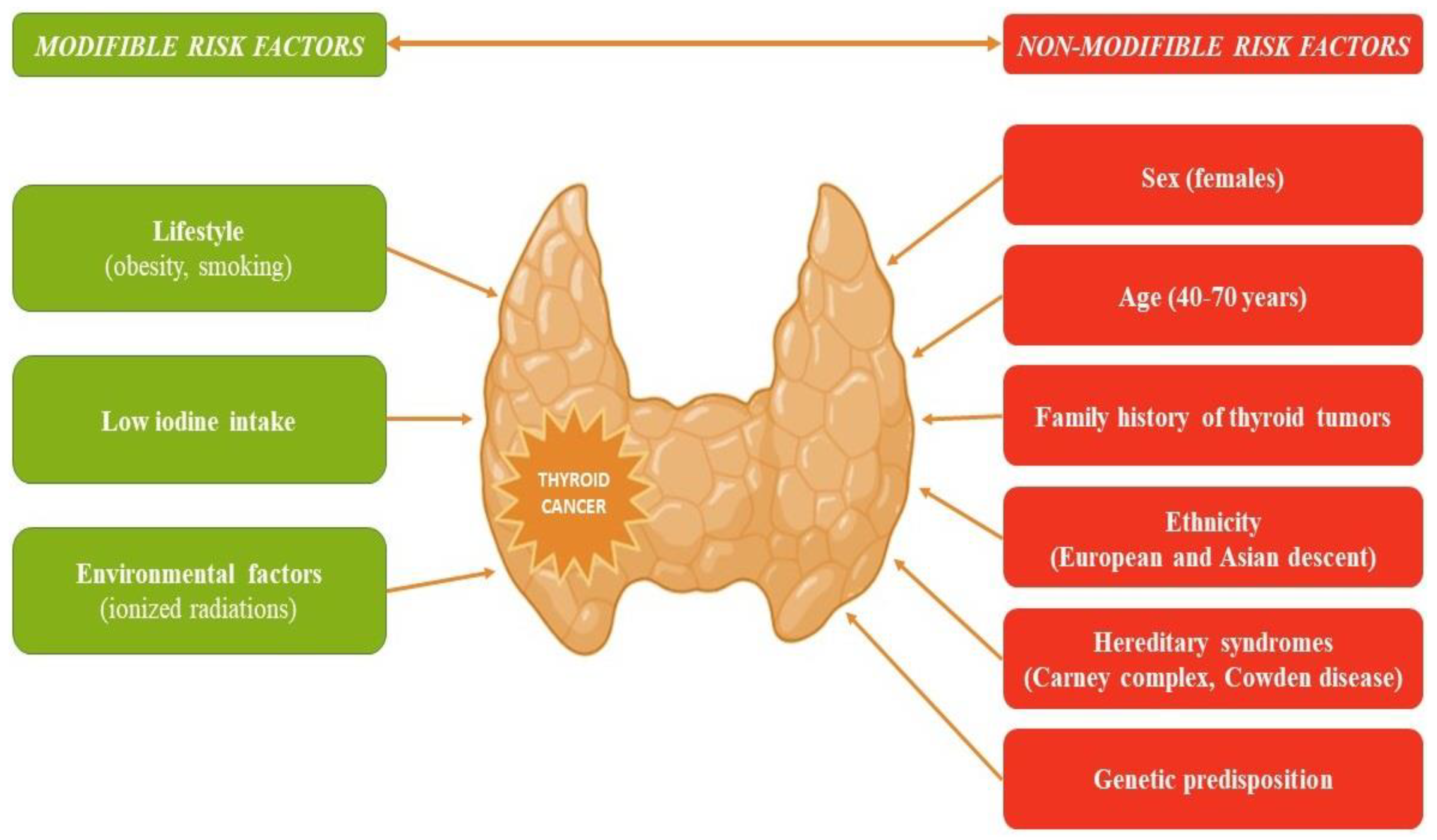
:1. Introduction
2. Methodology
3. Single Nucleotide Polymorphism and Thyroid Cancer
3.1. SNPs of Tyrosine-Kinase Family Genes
3.1.1. SNPs of RET Gene
3.1.2. SNPs of MET Gene
3.2. SNPs of Genes Involved in Apoptosis, Genome Stability, and DNA Repair
3.2.1. SNPs of BAX Gene
3.2.2. SNPs of TP53 and p21 Gene
3.2.3. SNPs of HOTAIR Gene
3.2.4. SNPs of XRCC1 Gene
3.3. SNPs of the VDR Gene
3.4. SNPs of Extracellular Matrix Genes with Roles in Cellular Proliferation and Differentiation
3.4.1. SNPs of SPARC and SPP1 Gene
3.4.2. SNPs of MMP-9 Gene
3.4.3. SNPs of NRG1 Gene
3.5. SNPs of Genes Involved in Thyroid Morphogenesis and Function
3.5.1. SNPs of FOXE1 Gene
3.5.2. SNPs of TSH-β Gene
3.6. SNPs of Genes Involved in Folate Metabolism
3.7. SNPs of Genes Involved in Inflammation
3.7.1. SNPs of IL-10 Gene
3.7.2. SNPs of IL-1 Gene
3.7.3. SNPs of NOS3 Gene
3.8. SNP in lncRNAs
4. Future Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanabria, A.; Kowalski, L.P.; Shah, J.P.; Nixon, I.J.; Angelos, P.; Williams, M.D.; Rinaldo, A.; Ferlito, A. Growing incidence of thyroid carcinoma in recent years: Factors underlying overdiagnosis. Head Neck 2018, 40, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Shah, J.P. Thyroid carcinoma: Epidemiology, histology, and diagnosis. Clin. Adv. Hematol. Oncol. 2015, 13 (Suppl. S4), 3–6. [Google Scholar] [PubMed]
- Liu, Y.; Su, L.; Xiao, H. Review of Factors Related to the Thyroid Cancer Epidemic. Int. J. Endocrinol. 2017, 2017, 5308635. [Google Scholar] [CrossRef] [Green Version]
- Orosco, R.K.; Hussain, T.; Brumund, K.T.; Oh, D.K.; Chang, D.C.; Bouvet, M. Analysis of age and disease status as predictors of thyroid cancer-specific mortality using the Surveillance, Epidemiology, and End Results database. Thyroid 2015, 25, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, Q.T.; Lee, E.J.; Huang, M.G.; Park, Y.I.; Khullar, A.; Plodkowski, R.A. Diagnosis and treatment of patients with thyroid cancer. Am. Health Drug Benefits 2015, 8, 30–40. [Google Scholar] [PubMed]
- Yang, J.; Gong, Y.; Yan, S.; Chen, H.; Qin, S.; Gong, R. Association between TERT promoter mutations and clinical behaviors in differentiated thyroid carcinoma: A systematic review and meta-analysis. Endocrine 2020, 67, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Melo, M.; da Rocha, A.G.; Batista, R.; Vingare, J.; Martins, M.J.; Costa, G.; Ribeiro, C.; Carrilho, F.; Leite, V.; Lobo, C.; et al. TERT, BRAF, and NRAS in Primary Thyroid Cancer and Metastatic Disease. J. Clin. Endocrinol. Metab. 2017, 102, 1898–1907. [Google Scholar] [CrossRef] [PubMed]
- Vuong, H.G.; Altibi, A.M.A.; Duong, U.N.P.; Hassell, L. Prognostic implication of BRAF and TERT promoter mutation combination in papillary thyroid carcinoma-A meta-analysis. Clin. Endocrinol. 2017, 87, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, R.; Shen, X.; Zhu, G.; Li, B.; Xing, M. The Genetic Duet of BRAF V600E and TERT Promoter Mutations Robustly Predicts Loss of Radioiodine Avidity in Recurrent Papillary Thyroid Cancer. J. Nucl. Med. 2020, 61, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Zhang, T.; Song, X.; Dong, G.; Xu, L.; Jiang, F. SNP-Target Genes Interaction Perturbing the Cancer Risk in the Post-GWAS. Cancers 2022, 14, 5636. [Google Scholar] [CrossRef]
- Huang, G.; Chen, J.; Zhou, J.; Xiao, S.; Zeng, W.; Xia, J.; Zeng, X. Epigenetic modification and BRAF gene mutation in thyroid carcinoma. Cancer Cell Int. 2021, 21, 687. [Google Scholar] [CrossRef] [PubMed]
- Kyrodimos, E.; Chrysovergis, A.; Mastronikolis, N.; Papanastasiou, G.; Tsiambas, E.; Spyropoulou, D.; Katsinis, S.; Manoli, A.; Papouliakos, S.; Pantos, P.; et al. The Landscape of Single Nucleotide Polymorphisms in Papillary Thyroid Carcinoma. Cancer Diagn. Progn. 2023, 3, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ares, M.; Cameselle-García, S.; Abdulkader-Nallib, I.; Rogriguez-Carnero, G.; Beiras-Sarasquete, C.; Puñal-Rodríguez, J.A.; Cameselle-Teijeiro, J.M. Susceptibility genes and chromosomal regions associated with non-syndromic familial non-medullary thyroid carcinoma: Some pathogenetic and diagnostic keys. Front. Endocrinol. 2022, 13, 829103. [Google Scholar] [CrossRef]
- Verrienti, A.; Carbone, A.; Sponziello, M.; Pecce, V.; Cito, D.S.; Bruno, R. Papillary thyroid carcinoma as first and isolated neoplastic disease in a Lynch syndrome family member with a germline MLH1 mutation. Endocrine 2022, 77, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Saenko, V.A.; Rogounovitch, T.I. Genetic Polymorphism Predisposing to Differentiated Thyroid Cancer: A Review of Major Findings of the Genome-Wide Association Studies. Endocrinol. Metab. 2018, 33, 164–174. [Google Scholar] [CrossRef]
- Carvajal-Carmona, L.G. Challenges in the identification and use of rare disease-associated predisposition variants. Curr. Opin. Genet. Dev. 2010, 20, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.-M.; Chung, W.-Y.; Hwang, D.-Y.; Yu, C.-C.; Ke, H.-L.; Liang, P.-I.; Lin, T.-W.; Cheng, S.M.; Huang, A.-M.; Kuo, H.-T. Whole-exome sequencing identified mutational profiles of urothelial carcinoma post kidney transplantation. J. Transl. Med. 2022, 20, 324. [Google Scholar] [CrossRef]
- Fessart, D.; Villamor, I.; Chevet, E.; Delom, F.; Robert, J. Integrative analysis of genomic and transcriptomic alterations of AGR2 and AGR3 in cancer. Open Biol. 2022, 12, 220068. [Google Scholar] [CrossRef]
- Rehman, K.; Jabeen, K.; Awan, F.R.; Hussain, M.; Saddique, M.A.; Akash, M.S.H. Biochemical investigation of rs1801282 variations in PPAR-γ gene and its correlation with risk factors of diabetes mellitus in coronary artery disease. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1517–1529. [Google Scholar] [CrossRef]
- Czene, K.; Lichtenstein, P.; Hemminki, K. Environmental and heritable causes of cancer among 9.6 million individuals in the Swedish Family-Cancer Database. Int. J. Cancer 2002, 99, 260–266. [Google Scholar] [CrossRef]
- Kerber, R.A.; O’Brien, E. A cohort study of cancer risk in relation to family histories of cancer in the Utah population database. Cancer 2005, 103, 1906–1915. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Kawai, K.; Asai, N. Roles of the RET Proto-oncogene in Cancer and Development. JMA J. 2020, 3, 175–181. [Google Scholar] [PubMed]
- Santos, M.; Azevedo, T.; Martins, T.; Rodrigues, F.J.; Lemos, M.C. Association of RET genetic polymorphisms and haplotypes with papillary thyroid carcinoma in the Portuguese population: A case-control study. PLoS ONE 2014, 9, e109822. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Ma, J.; Jiang, Y.; Su, X.; Zhang, X.; Chen, W.; Ye, Z. Associations between RET tagSNPs and their haplotypes and susceptibility, clinical severity, and thyroid function in patients with differentiated thyroid cancer. PLoS ONE 2017, 12, e0187968. [Google Scholar] [CrossRef] [Green Version]
- Ran, R.; Tu, G.; Li, H.; Wang, H.; Mou, E.; Liu, C. Genetic Variants Associated with Thyroid Cancer Risk: Comprehensive Research Synopsis, Meta-Analysis, and Cumulative Epidemiological Evidence. J. Oncol. 2021, 2021, 9967599. [Google Scholar] [CrossRef]
- Trusolino, L.; Comoglio, P.M. Scatter-factor and semaphorin receptors: Cell signalling for invasive growth. Nat. Rev. Cancer 2002, 2, 289–300. [Google Scholar] [CrossRef]
- Jung, K.H.; Park, B.H.; Hong, S.S. Progress in cancer therapy targeting c-Met signaling pathway. Arch. Pharm. Res. 2012, 35, 595–604. [Google Scholar] [CrossRef]
- Ning, L.; Yu, Y.; Liu, X.; Ai, L.; Zhang, X.; Rao, W.; Shi, J.; Sun, H.; Yu, Q. Association Analysis of MET Gene Polymorphism with Papillary Thyroid Carcinoma in a Chinese Population. Int. J. Endocrinol. 2015, 2015, 405217. [Google Scholar] [CrossRef] [Green Version]
- Uhliarova, B.; Hajtman, A. Hashimoto’s thyroiditis—An independent risk factor for papillary carcinoma. Braz. J. Otorhinolaryngol. 2018, 84, 729–735. [Google Scholar] [CrossRef]
- Sun, L.; Wei, L.; Wei, L.; Li, D. Correlation between Bax gene polymorphisms and esophagus cancer. Oncol. Lett. 2018, 16, 7097–7101. [Google Scholar] [CrossRef] [Green Version]
- Sahu, S.K.; Choudhuri, T. Lack of association between Bax promoter (-248G>A) single nucleotide polymorphism and susceptibility towards cancer: Evidence from a meta-analysis. PLoS ONE 2013, 8, e77534. [Google Scholar] [CrossRef]
- Cardoso-Duarte, L.C.A.; Fratelli, C.F.; Pereira, A.S.R.; de Souza, J.N.G.; de Souza Freitas, R.; de Morais, R.M.; Sobrinho, A.B.; Silva, C.M.S.; de Oliveira, J.R.; de Oliveira, D.M.; et al. BAX gene (-248 G > A) polymorphism in a sample of patients diagnosed with thyroid cancer in the Federal District, Brazil. Int. J. Biol. Markers 2021, 36, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Pflaum, J.; Schlosser, S.; Müller, M. p53 Family and Cellular Stress Responses in Cancer. Front. Oncol. 2014, 4, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, V.; Povirk, L. Involvement of p53 in the repair of DNA double strand breaks: Multifaceted Roles of p53 in homologous recombination repair (HRR) and non-homologous end joining (NHEJ). Subcell Biochem. 2014, 85, 321–336. [Google Scholar] [PubMed] [Green Version]
- Mauro, M.; Rego, M.A.; Boisvert, R.A.; Esashi, F.; Cavallo, F.; Jasin, M.; Howlett, N.G. p21 promotes error-free replication-coupled DNA double-strand break repair. Nucleic Acids Res. 2012, 40, 8348–8360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, T.; Hayward, R.L.; Meng, L.H.; Takemura, H.; Aune, G.J.; Bonner, W.M.; Aladjem, M.I.; Kohn, K.W.; Pommier, Y. p21CDKN1A allows the repair of replication-mediated DNA double-strand breaks induced by topoisomerase I and is inactivated by the checkpoint kinase inhibitor 7-hydroxystaurosporine. Oncogene 2006, 25, 2839–2849. [Google Scholar] [CrossRef] [Green Version]
- Heidari, Z.; Harati-Sadegh, M.; Arian, A.; Maruei-Milan, R.; Salimi, S. The effect of TP53 and P21 gene polymorphisms on papillary thyroid carcinoma susceptibility and clinical/pathological features. IUBMB Life 2020, 72, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Kou, C.; Chen, N.; Jia, L.; Sun, X.; Gao, Y.; Bai, R.; Yang, M.; Cui, J. Single Nucleotide Polymorphisms in HOTAIR Are Related to Breast Cancer Risk and Prognosis in the Northeastern Chinese Population. Front. Oncol. 2021, 11, 706428. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.O.; Jun, H.H.; Kim, E.J.; Lee, J.Y.; Park, H.S.; Ryu, C.S.; Kim, S.; Oh, D.; Kim, J.W.; Kim, N.K. Genetic Variants of HOTAIR Associated With Colorectal Cancer Susceptibility and Mortality. Front. Oncol. 2020, 10, 72. [Google Scholar] [CrossRef]
- Madadi Rad, R.; Pouladi, N.; Bosharani, N.N.; Alizadeh, A. Association between HOTAIR Rs1899663 G>T Gene Polymorphism and Thyroid Cancer Susceptibility. J. Babol. Univ. Med. Sci. 2022, 24, 95–102. [Google Scholar]
- Min, L.; Mu, X.; Tong, A.; Ling, C.; Yi, T.; Zhao, X. The association between HOTAIR polymorphisms and cancer susceptibility: An updated systemic review and meta-analysis. Onco. Targets Ther. 2018, 11, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Çağlayan, M.; Wilson, S.H. Oxidant and environmental toxicant-induced effects compromise DNA ligation during base excision DNA repair. DNA Repair 2015, 35, 85–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.Z.; Tan, X.R.; Zhao, M.; Mao, X.C.; Jiang, L. Association between the X-ray repair cross-complementing group 1 Arg194Trp polymorphism and thyroid carcinoma susceptibility: A meta-analysis. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Xue, W. XRCC1 Arg194Trp polymorphism and thyroid cancer. J. Endocrinol. Invest. 2020, 43, 749–753. [Google Scholar] [CrossRef]
- Bains, A.; Mur, T.; Wallace, N.; Noordzij, J.P. The Role of Vitamin D as a Prognostic Marker in Papillary Thyroid Cancer. Cancers 2021, 13, 3516. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, H.; Zhang, Z.; Zhou, X.; Yao, J.; Zhang, R.; Liao, L.; Dong, J. Vitamin D deficiency as a risk factor for thyroid cancer: A meta-analysis of case-control studies. Nutrition 2019, 57, 5–11. [Google Scholar] [CrossRef]
- Kim, J.R.; Kim, B.H.; Kim, S.M.; Oh, M.Y.; Kim, W.J.; Jeon, Y.K.; Kim, S.S.; Lee, B.L.; Kim, Y.K.; Kim, I.J. Low serum 25 hydroxyvitamin D is associated with poor clinicopathologic characteristics in female patients with papillary thyroid cancer. Thyroid 2014, 24, 1618–1624. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, C.; Chen, P.; Li, X.; Li, M.; Guo, H.; Li, J.; Chu, R.; Wang, H. Polymorphisms in the vitamin D Receptor (VDR) and the risk of ovarian cancer: A meta-analysis. PLoS ONE 2013, 8, e66716. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Chen, M.; Hu, X.; Wang, H.; Yang, J.; Zhang, C.; Pan, F.; Sun, G. Associations between VDR gene polymorphisms and colorectal cancer susceptibility: An updated meta-analysis based on 39 case-control studies. Oncotarget 2018, 9, 13068–13076. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.U.N.; Khan, T.A. Association between Vitamin D receptor (Cdx2, Fok1, Bsm1, Apa1, Bgl1, Taq1, and Poly (A)) gene polymorphism and breast cancer: A systematic review and meta-analysis. Tumour Biol. 2017, 39, 1010428317731280. [Google Scholar] [CrossRef] [Green Version]
- Beysel, S.; Eyerci, N.; Pinarli, F.A.; Apaydin, M.; Kizilgul, M.; Caliskan, M.; Ozcelik, O.; Kan, S.; Cakal, E. VDR gene FokI polymorphism as a poor prognostic factor for papillary thyroid cancer. Tumour Biol. 2018, 40, 1010428318811766. [Google Scholar] [CrossRef] [Green Version]
- Cocolos, A.M.; Muresan, A.; Caragheorgheopol, A.; Ghemigian, M.; Ioachim, D.; Poiana, C. Vitamin D Status and VDR Polymorphisms as Prognostic Factors in Differentiated Thyroid Carcinoma. In Vivo 2022, 36, 2434–2441. [Google Scholar] [CrossRef]
- Chiodoni, C.; Colombo, M.P.; Sangaletti, S. Matricellular proteins: From homeostasis to inflammation, cancer, and metastasis. Cancer Metastasis Rev. 2010, 29, 295–307. [Google Scholar] [CrossRef]
- Tu, Y.; Chen, C.; Fan, G. Association between the expression of secreted phosphoprotein—Related genes and prognosis of human cancer. BMC Cancer 2019, 19, 1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Chen, K.; Xu, W.; Chen, D.; Tang, W.; Xia, T.S. Integrative genomic analyses of secreted protein acidic and rich in cysteine and its role in cancer prediction. Mol. Med. Rep. 2014, 10, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Xu, B.-H.; Zhou, D.-L.; Ye, Z.-L.; He, H.-C.; Yang, X.-H.; Zhang, X.; Liu, Q.; Ma, J.-J.; Shao, Q.; et al. Polymorphisms in matricellular SPP1 and SPARC contribute to susceptibility to papillary thyroid cancer. Genomics 2020, 112, 4959–4967. [Google Scholar] [CrossRef]
- Zitka, O.; Kukacka, J.; Krizkova, S.; Huska, D.; Adam, V.; Masarik, M.; Prusa, R.; Kizek, R. Matrix metalloproteinases. Curr. Med. Chem. 2010, 17, 3751–3768. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Adhikari, N.; Banerjee, S.; Amin, S.A.; Jha, T. Matrix metalloproteinase-9 (MMP-9) and its inhibitors in cancer: A minireview. Eur. J. Med. Chem. 2020, 194, 112260, Erratum in Eur. J. Med. Chem. 2020, 205, 112642. [Google Scholar] [CrossRef]
- Farina, A.R.; Mackay, A.R. Gelatinase B/MMP-9 in Tumour Pathogenesis and Progression. Cancers 2014, 6, 240–296. [Google Scholar] [CrossRef]
- Dobrescu, R.; Picu, C.; Caragheorgheopol, A.; Danda, D.; Ioachim, D.; Goldstein, A.; Badiu, C. Serum Matrix metalloproteinase-9 (MMP-9) can help identify patients with papillary thyroid cancer at high risk of persistent disease: Value and limitations of a potential marker of neoplasia. Cancer Biomark. 2020, 29, 337–346. [Google Scholar] [CrossRef]
- Dobrescu, R.; Schipor, S.; Manda, D.; Caragheorgheopol, A.; Badiu, C. Matrix metalloproteinase-9 (MMP-9) promoter -1562C/T functional polymorphism is associated with an increased risk to develop micropapillary thyroid carcinoma. Cancer Biomark. 2022, 34, 555–562. [Google Scholar] [CrossRef]
- He, H.; Li, W.; Liyanarachchi, S.; Wang, Y.; Yu, L.; Genutis, L.K.; Maharry, S.; Phay, J.E.; Shen, R.; Brock, P.; et al. The Role of NRG1 in the Predisposition to Papillary Thyroid Carcinoma. J. Clin. Endocrinol. Metab. 2018, 103, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-L.; Feng, S.-H.; Guo, S.-C.; Wei, W.-J.; Li, D.-S.; Wang, Y.; Wang, X.; Wang, Z.-Y.; Ma, Y.-Y.; Jin, L.; et al. Confirmation of papillary thyroid cancer susceptibility loci identified by genome-wide association studies of chromosomes 14q13, 9q22, 2q35 and 8p12 in a Chinese population. J. Med. Genet. 2013, 50, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Gudmundsson, J.; Sulem, P.; Gudbjartsson, D.F.; Jonasson, J.G.; Masson, G.; He, H.; Jonasdottir, A.; Sigurdsson, A.; Stacey, S.A.; Johannsdottir, H.; et al. Discovery of common variants associated with low TSH levels and thyroid cancer risk. Nat. Genet. 2012, 44, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, W.; He, R.; Zheng, C.; Liu, X.; Ge, M.; Xu, J. Investigating the Association Between rs2439302 Polymorphism and Thyroid Cancer: A Systematic Review and Meta-Analysis. Front. Surg. 2022, 9, 877206. [Google Scholar] [CrossRef]
- Credendino, S.C.; Moccia, C.; Amendola, E.; D’Avino, G.; Di Guida, L.; Clery, E.; Greco, A.; Bellevicine, C.; Brunetti, A.; De Felice, M.; et al. FOXE1 Gene Dosage Affects Thyroid Cancer Histology and Differentiation In Vivo. Int. J. Mol. Sci. 2020, 22, 25. [Google Scholar] [CrossRef]
- Penna-Martinez, M.; Epp, F.; Kahles, H.; Ramos-Lopez, E.; Hinsch, N.; Hansmann, M.-L.; Selkinski, I.; Grünwald, F.; Holzer, K.; Bechstein, W.O.; et al. FOXE1 association with differentiated thyroid cancer and its progression. Thyroid 2014, 24, 845–851. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Zhang, Y.Q. Exploration of the association between FOXE1 gene polymorphism and differentiated thyroid cancer: A meta-analysis. BMC Med. Genet. 2018, 19, 83. [Google Scholar] [CrossRef]
- Mehrazin, A.; Safarpour, H.; Davoudi, S.T.; Parsamanesh, N.; Saeedi, F.; Miri-Moghaddam, E. Network-Based Analysis Reveals Association of FOXE1 Gene Polymorphisms in Thyroid Cancer Patients; A Case-Control Study in Southeast of Iran. Asian Pac. J. Cancer Prev. 2020, 21, 2771–2776. [Google Scholar] [CrossRef]
- Carvalho, D.P.; Dupuy, C. Thyroid hormone biosynthesis and release. Mol. Cell Endocrinol. 2017, 458, 6–15. [Google Scholar] [CrossRef]
- Sasaki, S.; Matsushita, A.; Kuroda, G.; Nakamura, H.M.; Oki, Y.; Suda, T. The mechanism of negative transcriptional regulation by thyroid hormone: Lessons from the thyrotropin β subunit gene. Vitam. Horm. 2018, 106, 97–127. [Google Scholar] [PubMed]
- Partsch, C.J.; Riepe, F.G.; Krone, N.; Sippell, W.G.; Pohlenz, J. Initially elevated TSH and congenital central hypothyroidism due to a homozygous mutation of the TSH beta subunit gene: Case report and review of the literature. Exp. Clin. Endocrinol. Diabetes 2006, 114, 227–234. [Google Scholar] [CrossRef]
- Vuissoz, J.M.; Deladoëy, J.; Buyukgebiz, A.; Cemeroglu, P.; Gex, G.; Gallati, S.; Mullis, P.E. New autosomal recessive mutation of the TSH-beta subunit gene causing central isolated hypothyroidism. J. Clin. Endocrinol. Metab. 2001, 86, 4468–4471. [Google Scholar]
- AlRasheed, M.M. TSH-β gene polymorphism in Saudi patients with thyroid cancer: A case-control study. Saudi Pharm. J. 2022, 30, 1538–1542. [Google Scholar] [CrossRef] [PubMed]
- Zara-Lopes, T.; Galbiatti-Dias, A.L.S.; Castanhole-Nunes, M.M.U.; Padovani-Júnior, J.A.; Maniglia, J.V.; Pavarino, E.C.; Goloni-Bertello, E.M. Polymorphisms in MTHFR, MTR, RFC1 and CßS genes involved in folate metabolism and thyroid cancer: A case-control study. Arch. Med. Sci. 2019, 15, 522–530. [Google Scholar] [CrossRef]
- Alshatwi, A.A. Breast cancer risk, dietary intake, and methylenetetrahydrofolate reductase (MTHFR)single nucleotide polymorphisms. Food Chem. Toxicol. 2018, 48, 1881–1885, Erratum in Food Chem. Toxicol. 2018, 118, 973. [Google Scholar] [CrossRef]
- Weiner, A.S.; Boyarskikh, U.A.; Voronina, E.N.; Voronina, E.N.; Selezneva, I.A.; Sinkina, T.V.; Lazarev, A.F.; Petrova, V.D.; Filipenko, M.L. Polymorphisms in the folate-metabolizing genes MTR, MTRR, and CBS and breast cancer risk. Cancer Epidemiol. 2012, 36, e95–e100. [Google Scholar] [CrossRef]
- Zhuo, X.; Song, J.; Li, D.; Wu, Y.; Zhou, Q. MTHFR C677T polymorphism interaction with heavy alcohol consumption increases head and neck carcinoma risk. Sci. Rep. 2015, 5, 10671. [Google Scholar] [CrossRef] [Green Version]
- Prasad, V.V.; Wilkhoo, H. Association of the functional polymorphism C677T in the methylenetetrahydrofolate reductase gene with colorectal, thyroid, breast, ovarian, and cervical cancers. Onkologie 2011, 34, 422–426. [Google Scholar] [CrossRef]
- Briukhovetska, D.; Dörr, J.; Endres, S.; Libby, P.; Dinarello, C.A.; Kobold, S. Interleukins in cancer: From biology to therapy. Nat. Rev. Cancer 2021, 21, 481–499. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Lumachi, F.; Basso, S.M.; Orlando, R. Cytokines, thyroid diseases and thyroid cancer. Cytokine 2010, 50, 229–233. [Google Scholar] [CrossRef]
- Zhu, Z.; Liu, J.B.; Liu, X.; Qian, L. Association of interleukin 10 rs1800896 polymorphism with susceptibility to breast cancer: A meta-analysis. J. Int. Med. Res. 2020, 48, 300060520904863. [Google Scholar] [CrossRef] [PubMed]
- Çil, E.; Kumral, A.; Kanmaz-Özer, M.; Vural, P.; Doğru-Abbasoğlu, S.; Altuntaş, Y.; Uysal, M. Interleukin-10-1082 gene polymorphism is associated with papillary thyroid cancer. Mol. Biol. Rep. 2014, 41, 3091–3097. [Google Scholar] [CrossRef] [PubMed]
- Apte, R.N.; Dotan, S.; Elkabets, M.; White, M.R.; Reich, E.; Carmi, Y.; Song, X.; Dvozkin, T.; Krelin, Y.; Voronov, E. The involvement of IL-1 in tumorigenesis, tumor invasiveness, metastasis and tumor-host interactions. Cancer Metastasis Rev. 2006, 25, 387–408. [Google Scholar] [CrossRef]
- Park, S.W.; Kim, M.K.; Kwon, K.H.; Kim, J. Association between a promoter polymorphism (rs2192752, -1028A/C) of interleukin 1 receptor, type I (IL1R1) and location of papillary thyroid carcinoma in a Korean population. Int. J. Immunogenet. 2012, 39, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Zhang, D.; Zhang, Y.; Yang, X.; Su, Y.; Wang, Y.; Huang, H.; Han, H.; Li, W.; Fu, K.; et al. Association of genetic polymorphisms in IL-1R1 and IL-1R2 genes with IgA nephropathy in the Han Chinese population. Oncotarget 2017, 8, 50673–50679. [Google Scholar] [CrossRef] [Green Version]
- Na, Y.; Bai, R.; Zhao, Z.; Wei, Y.; Li, D.; Wang, Y.; Sun, C.; Sun, L.; Zhang, B.; Jin, T.; et al. IL1R1 gene polymorphisms are associated with knee osteoarthritis risk in the Chinese Han population. Oncotarget 2017, 8, 4228–4233. [Google Scholar] [CrossRef] [Green Version]
- Vambutas, A.; DeVoti, J.; Goldofsky, E.; Gordon, M.; Lesser, M.; Bonagura, V. Alternate splicing of interleukin-1 receptor type II (IL1R2) in vitro correlates with clinical glucocorticoid responsiveness in patients with AIED. PLoS ONE 2009, 4, e5293. [Google Scholar] [CrossRef]
- Xiong, Z.; Sun, Y.; Wu, J.; Niu, F.; Jin, T.; Li, B. Genetic polymorphisms in IL1R1 and IL1R2 are associated with susceptibility to thyroid cancer in the Chinese Han population. J. Gene Med. 2019, 21, e3093. [Google Scholar] [CrossRef]
- Li, H.; Duan, N.; Zhang, Q.; Shao, Y. IL1A & IL1B genetic polymorphisms are risk factors for thyroid cancer in a Chinese Han population. Int. Immunopharmacol. 2019, 76, 105869. [Google Scholar]
- Ghimire, K.; Altmann, H.M.; Straub, A.C.; Isenberg, J.S. Nitric oxide: What’s new to NO? Am. J. Physiol. Cell Physiol. 2017, 312, C254–C262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mijatović, S.; Savić-Radojević, A.; Plješa-Ercegovac, M.; Simić, T.; Nicoletti, F.; Maksimović-Ivanić, D. The Double-Faced Role of Nitric Oxide and Reactive Oxygen Species in Solid Tumors. Antioxidants 2020, 9, 374. [Google Scholar] [CrossRef]
- Cerqueira, A.R.D.; Fratelli, C.F.; Duarte, L.C.A.C.; Pereira, A.S.R.; de Morais, R.M.; Sobrinho, A.B.; de Souza Silva, C.M.; da Silva, I.C.R.; de Oliveira, J.R. The impact of NOS3 gene polymorphism on papillary thyroid cancer susceptibility in patients undergoing radioiodine therapy. Int. J. Biol. Markers 2020, 35, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Maruei-Milan, R.; Heidari, Z.; Aryan, A.; Asadi-Tarani, M.; Salimi, S. Long non-coding RNA ANRIL polymorphisms in papillary thyroid cancer and its severity. Br. J. Biomed. Sci. 2021, 78, 58–62. [Google Scholar] [CrossRef]
- Tano, K.; Akimitsu, N. Long non-coding RNAs in cancer progression. Front. Genet. 2012, 3, 219. [Google Scholar] [CrossRef] [Green Version]
- Tornesello, M.L.; Faraonio, R.; Buonaguro, L.; Annunziata, C.; Starita, N.; Cerasuolo, A.; Pezzuto, F.; Tornesello, A.L.; Buonaguro, F.M. The Role of microRNAs, Long Non-coding RNAs, and Circular RNAs in Cervical Cancer. Front. Oncol. 2020, 10, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, R.; Shaheen, H.; Mansoor, Q.; Abbasi, S.A.; Fatima, S.; Ammar, A.; Baig, R.M. Genetic predisposition of SNPs in miRNA-149 (rs2292832) and FOXE1 (rs3758249) in thyroid Cancer. Mol. Biol. Rep. 2021, 48, 7801–7809. [Google Scholar] [CrossRef]
- Khan, R.; Abbasi, S.A.; Mansoor, Q.; Ahmed, M.N.; Mir, K.B.; Baig, R.M. Analysis of Rare Alleles of miRNA-146a (rs2910164) and miRNA-34b/c (rs4938723) as a Prognostic Marker in Thyroid Cancer in Pakistani Population. Diagnostics 2022, 12, 2495. [Google Scholar] [CrossRef]
- Yang, M.; Tian, J.; Guo, X.; Yang, Y.; Guan, R.; Qiu, M.; Li, Y.; Sun, X.; Zhen, Y.; Zhang, Y.; et al. Long noncoding RNA are aberrantly expressed in human papillary thyroid carcinoma. Oncol. Lett. 2016, 12, 544–552. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Zhang, K.; Ma, L.; Xu, J.; Chang, W. The Role of Long Non-Coding RNAs in Thyroid Cancer. Front. Oncol. 2020, 10, 941. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Lv, Z.; An, C.; Shi, M.; Pan, W.; Zhou, L.; Yang, W.; Yang, M. Onco-lncRNA HOTAIR and its functional genetic variants in papillary thyroid carcinoma. Sci. Rep. 2016, 6, 31969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.-J.; Hao, S.; Wang, L.-L.; Hu, C.-Y.; Zhang, S.; Guo, L.-J.; Zhang, G.; Gao, B.; Jiang, Y.; Tian, W.-G.; et al. Long non-coding RNA ANRIL promotes the invasion and metastasis of thyroid cancer cells through TGF-β/Smad signaling pathway. Oncotarget 2016, 7, 57903–57918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jendrzejewski, J.; He, H.; Radomska, H.S.; Li, W.; Tomsic, J.; Liyanarachchi, S.; Davuluri, R.V.; Nagy, R.; de la Chapelle, A. The polymorphism rs944289 predisposes to papillary thyroid carcinoma through a large intergenic noncoding RNA gene of tumor suppressor type. Proc. Natl. Acad. Sci. USA 2012, 109, 8646–8651. [Google Scholar] [CrossRef] [Green Version]
- Jendrzejewski, J.; Thomas, A.; Liyanarachchi, S.; Eiterman, A.; Tomsic, J.; He, H.; Radomska, H.S.; Li, W.; Nagy, R.; Sworczak, K.; et al. PTCSC3 Is Involved in Papillary Thyroid Carcinoma Development by Modulating S100A4 Gene Expression. J. Clin. Endocrinol. Metab. 2015, 100, E1370–E1377. [Google Scholar] [CrossRef] [Green Version]
- Liao, T.; Qu, N.; Shi, R.L.; Guo, K.; Ma, B.; Cao, Y.-M.; Xiang, J.; Lu, Z.-W.; Zhu, Y.-X.; Li, D.-S.; et al. BRAF-activated LncRNA functions as a tumor suppressor in papillary thyroid cancer. Oncotarget 2017, 8, 238–247. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Yan, G.; Zhang, Y.; Jia, X.; Bu, P. Long non-coding RNA MEG3 suppresses migration and invasion of thyroid carcinoma by targeting of Rac1. Neoplasma 2015, 62, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.-K.; Ma, L.; Song, W.-H.; Lu, B.-Y.; Huang, Y.-B.; Dong, H.-M.; Ma, X.-K.; Zhu, Z.-Z.; Zhou, R. LncRNA-MALAT1 Promotes Angiogenesis of Thyroid Cancer by Modulating Tumor-Associated Macrophage FGF2 Protein Secretion. J. Cell Biochem. 2017, 118, 4821–4830. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.-D.; Ye, C.-X.; Shen, Z.-Y.; Yang, Y.; Wang, B.; Guo, P.; Gao, Z.-D.; Ye, Y.-J.; Jiang, K.-W.; et al. Long noncoding RNA HIT000218960 promotes papillary thyroid cancer oncogenesis and tumor progression by upregulating the expression of high mobility group AT-hook 2 (HMGA2) gene. Cell Cycle 2017, 16, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Zhang, Q.; Chen, W.; Zhang, H.; Lu, G.; Chen, J.; Qiu, C. Long noncoding RNA cancer susceptibility candidate 2 suppresses papillary thyroid carcinoma growth by inactivating the AKT/ERK1/2 signaling pathway. J. Cell Biochem. 2019, 120, 10380–10390. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, L.; Wang, H.; Chen, J.; Li, W.; Liu, W.; Shen, M.; Liu, H.; Fu, X. Long noncoding RNA CNALPTC1 promotes cell proliferation and migration of papillary thyroid cancer via sponging miR-30 family. Am. J. Cancer Res. 2018, 8, 192–206. [Google Scholar] [PubMed]
- Chen, B.; Li, J.; Yi, C.; Jiao, Y.; Gu, X.; Feng, X. Long non-coding RNA POLR2E rs3787016 is associated with the risk of papillary thyroid carcinoma in Chinese population. Pathol. Res. Pract. 2018, 214, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Chen, L.; Tian, H.; Li, J.; Zhang, M.; Cao, Q.; Zhang, W.; Chen, S.; Shi, L. Effect of MALAT1 Polymorphisms on Papillary Thyroid Cancer in a Chinese Population. J. Cancer 2019, 10, 5714–5721. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Huang, H.; Chen, P.; Xiao, X.; Luo, X.; Wang, Y.; Long, S.; Gao, L.; Zhang, L. Genetic predisposition to papillary thyroid carcinoma is mediated by a long non-coding RNA TINCR enhancer polymorphism. Int. Immunopharmacol. 2022, 109, 108796. [Google Scholar] [CrossRef]


{kind=link}
| Gene | Category | SNP | Main Results | Reference |
|---|---|---|---|---|
| RET | Protein-coding | S836S | Overexpressed in patients with PTC | [23] |
| G691S | Identified in tumors > 10 mm | |||
| S904S | ||||
| rs17028, rs1799939, rs1800858, rs1800860, rs2075912, rs2565200, rs2742240 |
| [24] | ||
| rs1799939 | Increased thyroid cancer susceptibility | [25] | ||
| BAX | Protein-coding | −248 G > A |
| [32] |
| TSH-β | Protein-coding | rs201857310 | Strongly associated with thyroid cancer | [74] |
| VDR | Protein-coding | rs2228570 |
| [51,52] |
| SPARC | Protein-coding | rs1054204, rs3210714, rs3549 | Reduced PTC risk | [56] |
| SPP1 | Protein-coding | rs4754 | Increased PTC risk | [56] |
| XRCC1 | Protein-coding | Arg194Trp | Increased thyroid cancer risk | [43] |
| C allele: decreased thyroid cancer risk | [44] | |||
| NRG1 | Protein-coding | rs2439302 |
| [62] |
| Increased thyroid cancer risk | [63,64,65] | |||
| HOTAIR | Non-coding | rs1899663 | No association with clinicopathological traits of thyroid cancer | [40] |
| No association with increased thyroid cancer risk | [41] | |||
| rs7958904, rs4759314, rs874945 | No association with increased thyroid cancer risk | [41] | ||
| rs12826786, rs920778 | Increased thyroid cancer risk | [41] | ||
| TP53 | Protein-coding | rs1042522 |
| [37] |
| p21 | Protein-coding | rs10559234, rs1801270 | No association with increased thyroid cancer risk | [37] |
| FOXE1 | Protein-coding | rs965513 | Increased susceptibility for DTC | [67,68] |
| rs944289, rs1867277 | [68,69] | |||
| IL-10 | Protein-coding | G-1082A |
| [84] |
| IL1R1 | Protein-coding | rs3917225 | Increased susceptibility for thyroid cancer | [90] |
| IL1R2 | Protein-coding | rs2072472, rs11674595 | Increased susceptibility for thyroid cancer | [90] |
| IL1A | Protein-coding | rs3783521, rs3783546, rs3783550, rs1609682 | Increased susceptibility for thyroid cancer | [91] |
| IL1B | Protein-coding | rs3136558, rs1143623 | Increased susceptibility for thyroid cancer | [91] |
| MET | Protein-coding | rs1621 |
| [28] |
| NOS3 | Protein-coding | Intron4 |
| [94] |
| MTHFR | Protein-coding | 677C > T | Increased risk for thyroid cancer | [75] |
| MTR | Protein-coding | 2756A > G | Associated with histopathological traits such as tumor extent and aggressive behavior | [75] |
| RFC1 | Protein-coding | 80A > G | Increased risk for thyroid cancer | [75] |
| MMP-9 | Protein-coding | −1562C/T | CT or CT + TT genotype: increased risk for PTC | [61] |
| MIR149 | Non-coding | rs2292832 | Mutation T > C was strongly associated with thyroid cancer | [98] |
| MIR34B | Non-coding | rs4938723 | Increased risk for thyroid cancer | [99] |
| ANRIL | Non-coding | rs11333048, rs4977574, rs4977574, rs1333040, rs10757274 |
| [95] |
| MALAT1 | Non-coding | rs619586 | Protective factor for PTC | [113] |
| POLR2E | Protein-coding | rs3787916 | Increased risk for thyroid cancer | [112] |
| TINCR | Protein-coding | rs8101923 | G allele: high risk for PTC | [114] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiucă, R.A.; Tiucă, O.M.; Pașcanu, I.M. The Role of Genetic Polymorphisms in Differentiated Thyroid Cancer: A 2023 Update. Biomedicines 2023, 11, 1075. https://doi.org/10.3390/biomedicines11041075
Tiucă RA, Tiucă OM, Pașcanu IM. The Role of Genetic Polymorphisms in Differentiated Thyroid Cancer: A 2023 Update. Biomedicines. 2023; 11(4):1075. https://doi.org/10.3390/biomedicines11041075
Chicago/Turabian StyleTiucă, Robert Aurelian, Oana Mirela Tiucă, and Ionela Maria Pașcanu. 2023. "The Role of Genetic Polymorphisms in Differentiated Thyroid Cancer: A 2023 Update" Biomedicines 11, no. 4: 1075. https://doi.org/10.3390/biomedicines11041075