

Impact of Sacubitril/Valsartan on Circulating microRNA in Patients with Heart Failure
 , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Plasma Preparation
2.3. Total RNA Purification
2.4. MicroRNA Screening
2.5. Single miRNA Assays
2.6. Statistical Analysis
3. Results
3.1. Characteristics of the Study Population
3.2. Profiling of Circulating miRNA Expression
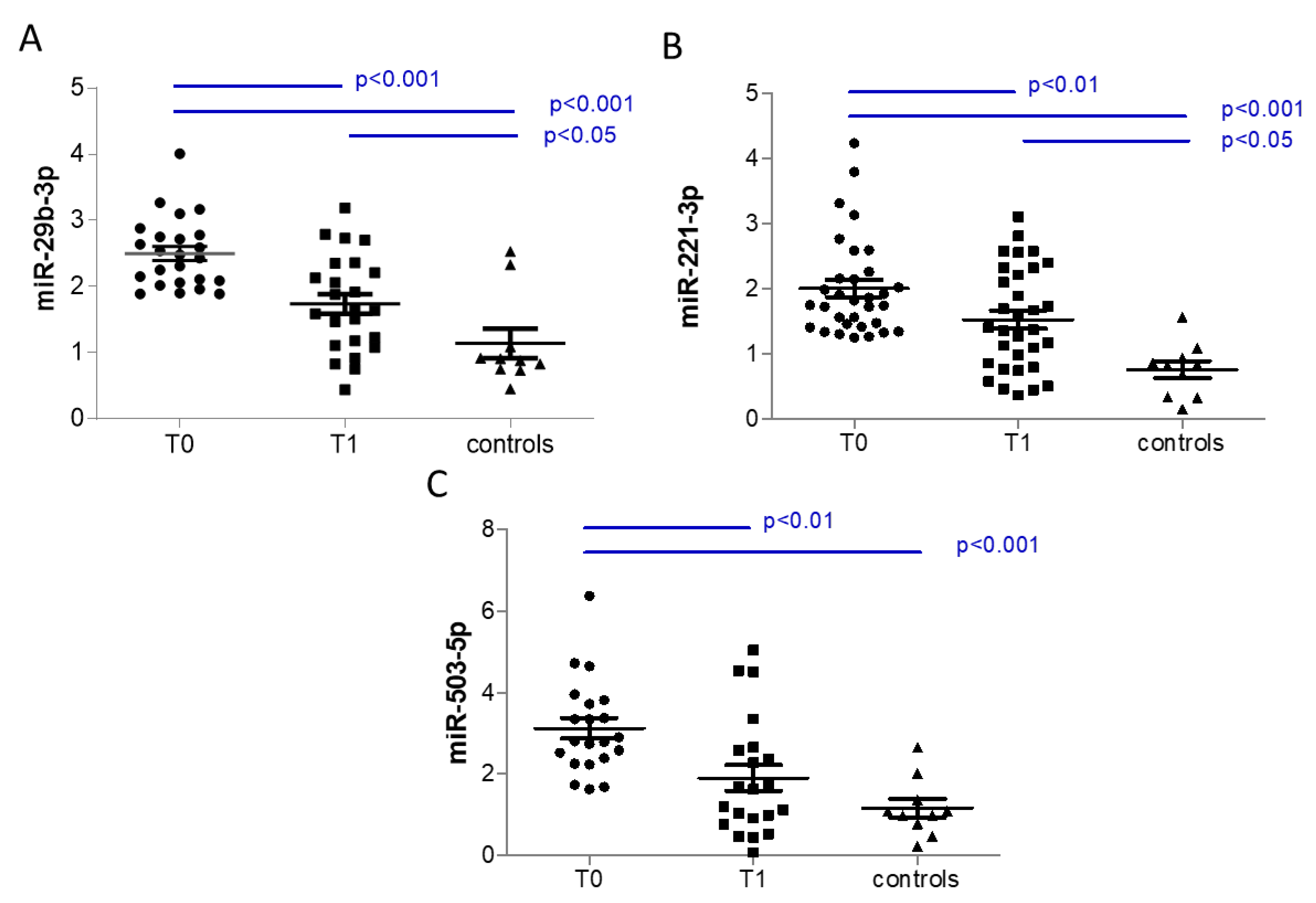
3.3. Analysis of Selected Circulating miRNAs
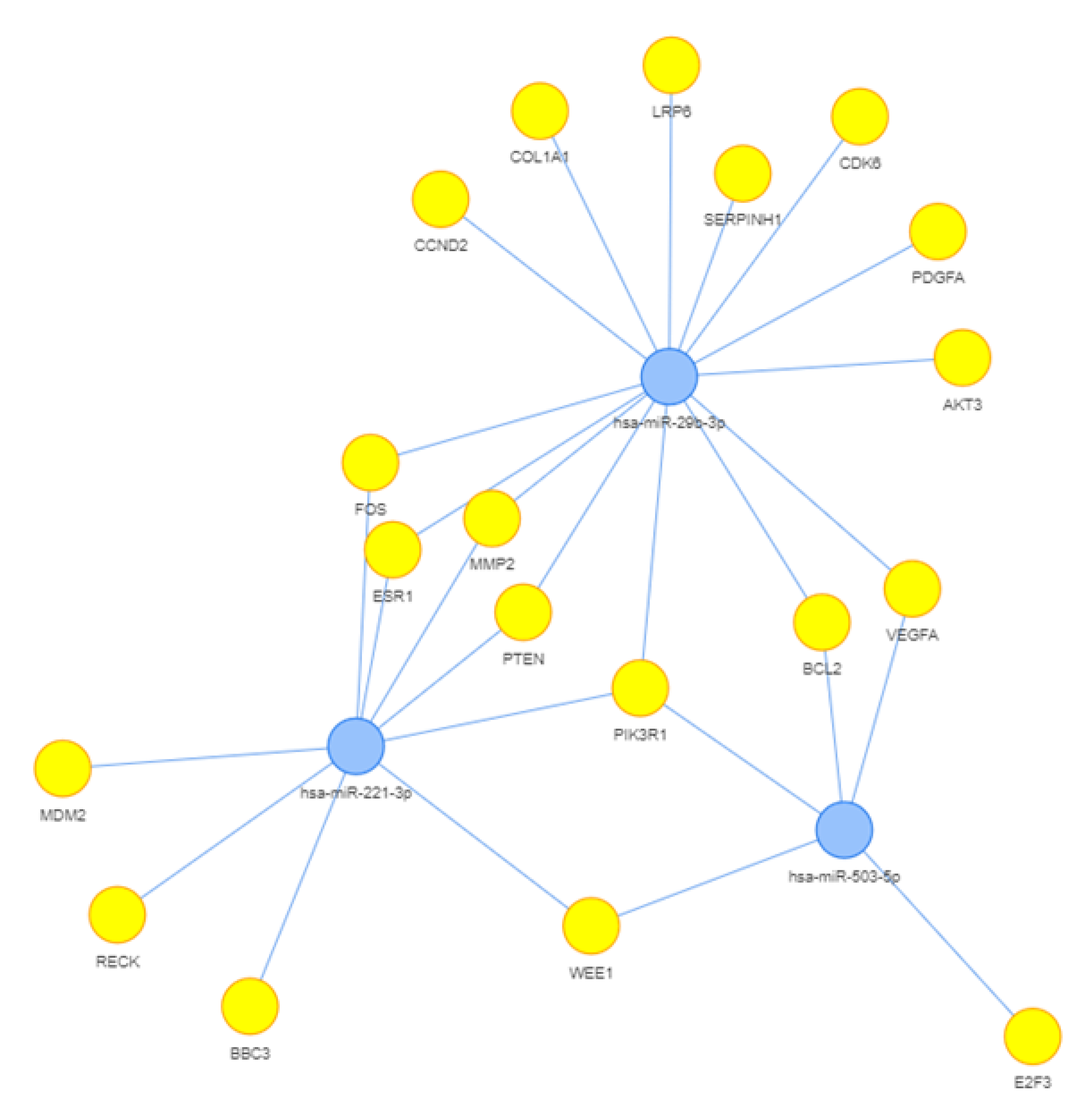
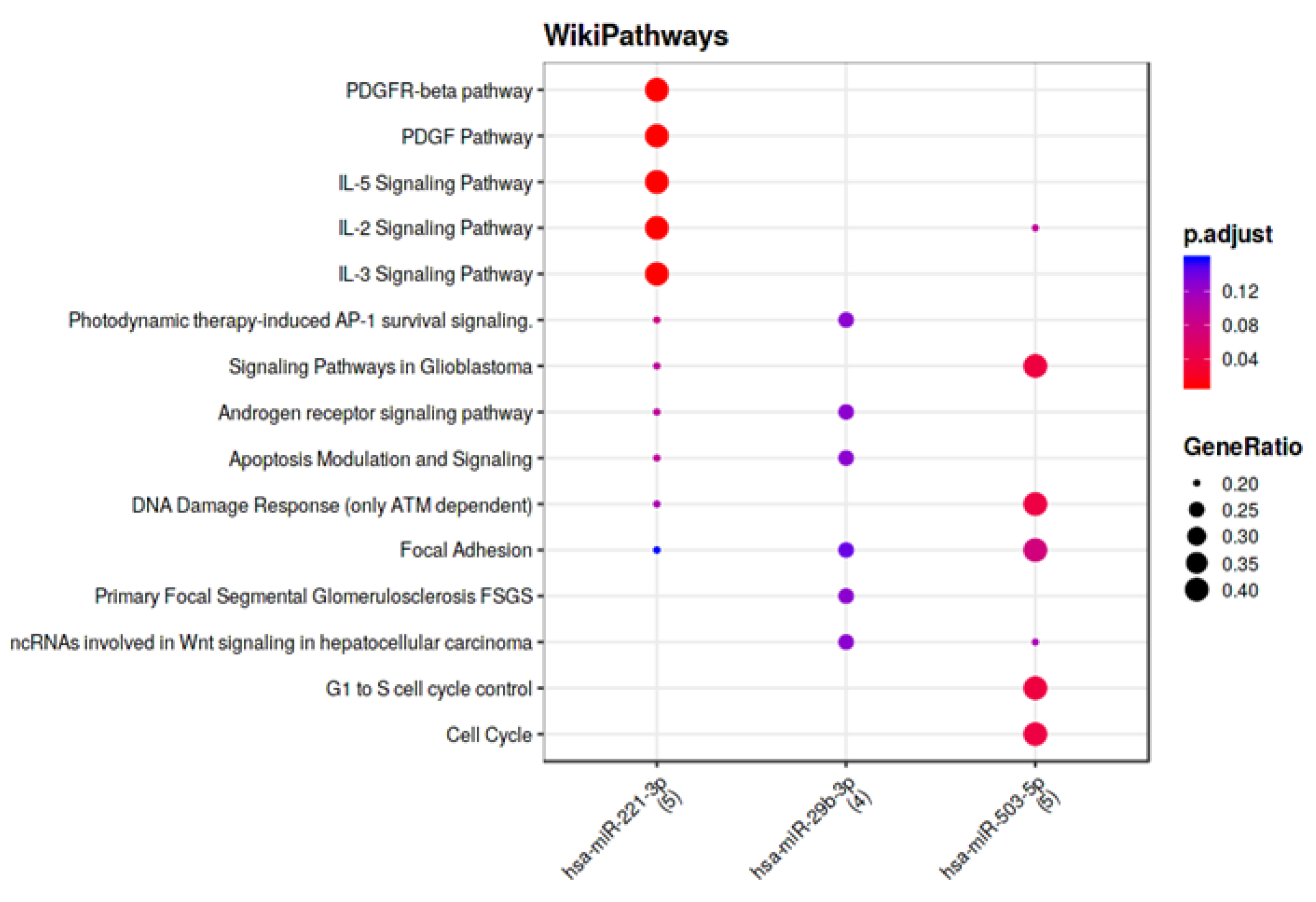
3.4. Computational Analysis of miRNA Targets
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McDonagh, T.A.; Metra, M.; Adamo, M.; Gardner, R.S.; Baumbach, A.; Bohm, M.; Burri, H.; Butler, J.; Celutkiene, J.; Chioncel, O.; et al. 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur. Heart J. 2021, 42, 3599–3726. [Google Scholar] [CrossRef]
- McMurray, J.J.; Packer, M.; Desai, A.S.; Gong, J.; Lefkowitz, M.P.; Rizkala, A.R.; Rouleau, J.; Shi, V.C.; Solomon, S.D.; Swedberg, K.; et al. Dual angiotensin receptor and neprilysin inhibition as an alternative to angiotensin-converting enzyme inhibition in patients with chronic systolic heart failure: Rationale for and design of the Prospective comparison of ARNI with ACEI to Determine Impact on Global Mortality and morbidity in Heart Failure trial (PARADIGM-HF). Eur. J. Heart Fail. 2013, 15, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Mantegazza, V.; Volpato, V.; Mapelli, M.; Sassi, V.; Salvioni, E.; Mattavelli, I.; Tamborini, G.; Agostoni, P.; Pepi, M. Cardiac Reverse Remodelling by 2D and 3D Echocardiography in Heart Failure Patients Treated with Sacubitril/Valsartan. Diagnostics 2021, 11, 1845. [Google Scholar] [CrossRef] [PubMed]
- Mapelli, M.; Mattavelli, I.; Salvioni, E.; Banfi, C.; Ghilardi, S.; De Martino, F.; Gugliandolo, P.; Mantegazza, V.; Volpato, V.; Basile, C.; et al. Impact of Sacubitril/Valsartan on surfactant binding proteins, central sleep apneas, lung function tests and heart failure biomarkers: Hemodynamic or pleiotropism? Front. Cardiovasc. Med. 2022, 9, 971108. [Google Scholar] [CrossRef] [PubMed]
- Gargiulo, P.; Banfi, C.; Ghilardi, S.; Magri, D.; Giovannardi, M.; Bonomi, A.; Salvioni, E.; Battaia, E.; Filardi, P.P.; Tremoli, E.; et al. Surfactant-derived proteins as markers of alveolar membrane damage in heart failure. PLoS ONE 2014, 9, e115030. [Google Scholar] [CrossRef]
- Banfi, C.; Agostoni, P. Surfactant protein B: From biochemistry to its potential role as diagnostic and prognostic marker in heart failure. Int. J. Cardiol. 2016, 221, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Magri, D.; Brioschi, M.; Banfi, C.; Schmid, J.P.; Palermo, P.; Contini, M.; Apostolo, A.; Bussotti, M.; Tremoli, E.; Sciomer, S.; et al. Circulating plasma surfactant protein type B as biological marker of alveolar-capillary barrier damage in chronic heart failure. Circ. Heart Fail. 2009, 2, 175–180. [Google Scholar] [CrossRef]
- Magri, D.; Banfi, C.; Maruotti, A.; Farina, S.; Vignati, C.; Salvioni, E.; Morosin, M.; Brioschi, M.; Ghilardi, S.; Tremoli, E.; et al. Plasma immature form of surfactant protein type B correlates with prognosis in patients with chronic heart failure. A pilot single-center prospective study. Int. J. Cardiol. 2015, 201, 394–399. [Google Scholar] [CrossRef]
- Campodonico, J.; Mapelli, M.; Spadafora, E.; Ghilardi, S.; Agostoni, P.; Banfi, C.; Sciomer, S. Surfactant proteins changes after acute hemodynamic improvement in patients with advanced chronic heart failure treated with Levosimendan. Respir. Physiol. Neurobiol. 2018, 252–253, 47–51. [Google Scholar] [CrossRef]
- Dutka, M.; Bobinski, R.; Ulman-Wlodarz, I.; Hajduga, M.; Bujok, J.; Pajak, C.; Cwiertnia, M. Sodium glucose cotransporter 2 inhibitors: Mechanisms of action in heart failure. Heart Fail. Rev. 2021, 26, 603–622. [Google Scholar] [CrossRef]
- Pillai, R.S.; Bhattacharyya, S.N.; Filipowicz, W. Repression of protein synthesis by miRNAs: How many mechanisms? Trends Cell Biol. 2007, 17, 118–126. [Google Scholar] [CrossRef]
- Pozniak, T.; Shcharbin, D.; Bryszewska, M. Circulating microRNAs in Medicine. Int. J. Mol. Sci. 2022, 23, 3996. [Google Scholar] [CrossRef]
- Bar, C.; Thum, T.; de Gonzalo-Calvo, D. Circulating miRNAs as mediators in cell-to-cell communication. Epigenomics 2019, 11, 111–113. [Google Scholar] [CrossRef]
- Berry, R.B.; Brooks, R.; Gamaldo, C.; Harding, S.M.; Lloyd, R.M.; Quan, S.F.; Troester, M.T.; Vaughn, B.V. AASM Scoring Manual Updates for 2017 (Version 2.4). J. Clin. Sleep Med. 2017, 13, 665–666. [Google Scholar] [CrossRef]
- D’Alessandra, Y.; Chiesa, M.; Carena, M.C.; Beltrami, A.P.; Rizzo, P.; Buzzetti, M.; Ricci, V.; Ferrari, R.; Fucili, A.; Livi, U.; et al. Differential Role of Circulating microRNAs to Track Progression and Pre-Symptomatic Stage of Chronic Heart Failure: A Pilot Study. Biomedicines 2020, 8, 597. [Google Scholar] [CrossRef] [PubMed]
- Sassi, Y.; Avramopoulos, P.; Ramanujam, D.; Gruter, L.; Werfel, S.; Giosele, S.; Brunner, A.D.; Esfandyari, D.; Papadopoulou, A.S.; De Strooper, B.; et al. Cardiac myocyte miR-29 promotes pathological remodeling of the heart by activating Wnt signaling. Nat. Commun. 2017, 8, 1614. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Verma, G.; Vig, S.; Srivastava, S.; Srivastava, A.K.; Datta, M. miR-29a levels are elevated in the db/db mice liver and its overexpression leads to attenuation of insulin action on PEPCK gene expression in HepG2 cells. Mol. Cell. Endocrinol. 2011, 332, 125–133. [Google Scholar] [CrossRef]
- Vaskova, E.; Ikeda, G.; Tada, Y.; Wahlquist, C.; Mercola, M.; Yang, P.C. Sacubitril/Valsartan Improves Cardiac Function and Decreases Myocardial Fibrosis Via Downregulation of Exosomal miR-181a in a Rodent Chronic Myocardial Infarction Model. J. Am. Heart Assoc. 2020, 9, e015640. [Google Scholar] [CrossRef] [PubMed]
- Sardu, C.; Massetti, M.; Scisciola, L.; Trotta, M.C.; Santamaria, M.; Volpicelli, M.; Ducceschi, V.; Signoriello, G.; D’Onofrio, N.; Marfella, L.; et al. Angiotensin receptor/Neprilysin inhibitor effects in CRTd non-responders: From epigenetic to clinical beside. Pharmacol. Res. 2022, 182, 106303. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, G.A.; Elwahab, A.H.A.; Hasan, R.A.; ElMongy, N.F.; Ramzy, M.M.; Louka, M.L.; Schaalan, M.F. A novel protective role of sacubitril/valsartan in cyclophosphamide induced lung injury in rats: Impact of miRNA-150-3p on NF-kappaB/MAPK signaling trajectories. Sci. Rep. 2020, 10, 13045. [Google Scholar] [CrossRef]
- Mott, J.L.; Kobayashi, S.; Bronk, S.F.; Gores, G.J. mir-29 regulates Mcl-1 protein expression and apoptosis. Oncogene 2007, 26, 6133–6140. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Heaphy, C.E.; Havelange, V.; Fabbri, M.; Volinia, S.; Tsao, T.; Zanesi, N.; Kornblau, S.M.; Marcucci, G.; Calin, G.A.; et al. MicroRNA 29b functions in acute myeloid leukemia. Blood 2009, 114, 5331–5341. [Google Scholar] [CrossRef] [PubMed]
- Sardu, C.; Marfella, R.; Santulli, G.; Paolisso, G. Functional role of miRNA in cardiac resynchronization therapy. Pharmacogenomics 2014, 15, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- van Rooij, E.; Sutherland, L.B.; Thatcher, J.E.; DiMaio, J.M.; Naseem, R.H.; Marshall, W.S.; Hill, J.A.; Olson, E.N. Dysregulation of microRNAs after myocardial infarction reveals a role of miR-29 in cardiac fibrosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13027–13032. [Google Scholar] [CrossRef]
- O’Brien, J.E., Jr.; Kibiryeva, N.; Zhou, X.G.; Marshall, J.A.; Lofland, G.K.; Artman, M.; Chen, J.; Bittel, D.C. Noncoding RNA expression in myocardium from infants with tetralogy of Fallot. Circ. Cardiovasc. Genet. 2012, 5, 279–286. [Google Scholar] [CrossRef]
- Yang, Q.; Wu, F.; Mi, Y.; Wang, F.; Cai, K.; Yang, X.; Zhang, R.; Liu, L.; Zhang, Y.; Wang, Y.; et al. Aberrant expression of miR-29b-3p influences heart development and cardiomyocyte proliferation by targeting NOTCH2. Cell Prolif. 2020, 53, e12764. [Google Scholar] [CrossRef]
- van den Akker, N.M.; Molin, D.G.; Peters, P.P.; Maas, S.; Wisse, L.J.; van Brempt, R.; van Munsteren, C.J.; Bartelings, M.M.; Poelmann, R.E.; Carmeliet, P.; et al. Tetralogy of fallot and alterations in vascular endothelial growth factor-A signaling and notch signaling in mouse embryos solely expressing the VEGF120 isoform. Circ. Res. 2007, 100, 842–849. [Google Scholar] [CrossRef]
- MacGrogan, D.; Luna-Zurita, L.; de la Pompa, J.L. Notch signaling in cardiac valve development and disease. Birth Defects Res. A Clin. Mol. Teratol. 2011, 91, 449–459. [Google Scholar] [CrossRef]
- Su, M.; Wang, J.; Wang, C.; Wang, X.; Dong, W.; Qiu, W.; Wang, Y.; Zhao, X.; Zou, Y.; Song, L.; et al. MicroRNA-221 inhibits autophagy and promotes heart failure by modulating the p27/CDK2/mTOR axis. Cell Death Differ. 2015, 22, 986–999. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yan, C.; Fan, J.; Hou, Z.; Han, Y. MiR-221-3p targets Hif-1alpha to inhibit angiogenesis in heart failure. Lab. Investig. 2021, 101, 104–115. [Google Scholar] [CrossRef]
- Wang, C.; Wang, S.; Zhao, P.; Wang, X.; Wang, J.; Wang, Y.; Song, L.; Zou, Y.; Hui, R. MiR-221 promotes cardiac hypertrophy in vitro through the modulation of p27 expression. J. Cell. Biochem. 2012, 113, 2040–2046. [Google Scholar] [CrossRef] [PubMed]
- Caporali, A.; Emanueli, C. MicroRNA-503 and the extended microRNA-16 family in angiogenesis. Trends Cardiovasc. Med. 2011, 21, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Caporali, A.; Meloni, M.; Vollenkle, C.; Bonci, D.; Sala-Newby, G.B.; Addis, R.; Spinetti, G.; Losa, S.; Masson, R.; Baker, A.H.; et al. Deregulation of microRNA-503 contributes to diabetes mellitus-induced impairment of endothelial function and reparative angiogenesis after limb ischemia. Circulation 2011, 123, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.; Hou, J.; Xuan, W.; Zhang, C.; Meng, X. The relationship of plasma miR-503 and coronary collateral circulation in patients with coronary artery disease. Life Sci. 2018, 207, 145–151. [Google Scholar] [CrossRef]
- Zhang, H.; Pan, Q.; Xie, Z.; Chen, Y.; Wang, J.; Bihl, J.; Zhong, W.; Chen, Y.; Zhao, B.; Ma, X. Implication of MicroRNA503 in Brain Endothelial Cell Function and Ischemic Stroke. Transl. Stroke Res. 2020, 11, 1148–1164. [Google Scholar] [CrossRef]
- He, Y.; Cai, Y.; Pai, P.M.; Ren, X.; Xia, Z. The Causes and Consequences of miR-503 Dysregulation and Its Impact on Cardiovascular Disease and Cancer. Front. Pharmacol. 2021, 12, 629611. [Google Scholar] [CrossRef]
- Zhou, Y.; Deng, L.; Zhao, D.; Chen, L.; Yao, Z.; Guo, X.; Liu, X.; Lv, L.; Leng, B.; Xu, W.; et al. MicroRNA-503 promotes angiotensin II-induced cardiac fibrosis by targeting Apelin-13. J. Cell. Mol. Med. 2016, 20, 495–505. [Google Scholar] [CrossRef]
- Sanada, S.; Hakuno, D.; Higgins, L.J.; Schreiter, E.R.; McKenzie, A.N.; Lee, R.T. IL-33 and ST2 comprise a critical biomechanically induced and cardioprotective signaling system. J. Clin. Investig. 2007, 117, 1538–1549. [Google Scholar] [CrossRef]
- Schmitz, J.; Owyang, A.; Oldham, E.; Song, Y.; Murphy, E.; McClanahan, T.K.; Zurawski, G.; Moshrefi, M.; Qin, J.; Li, X.; et al. IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity 2005, 23, 479–490. [Google Scholar] [CrossRef]
- van Vark, L.C.; Lesman-Leegte, I.; Baart, S.J.; Postmus, D.; Pinto, Y.M.; Orsel, J.G.; Westenbrink, B.D.; Brunner-la Rocca, H.P.; van Miltenburg, A.J.M.; Boersma, E.; et al. Prognostic Value of Serial ST2 Measurements in Patients With Acute Heart Failure. J. Am. Coll. Cardiol. 2017, 70, 2378–2388. [Google Scholar] [CrossRef]
- Wang, E.W.; Jia, X.S.; Ruan, C.W.; Ge, Z.R. miR-487b mitigates chronic heart failure through inhibition of the IL-33/ST2 signaling pathway. Oncotarget 2017, 8, 51688–51702. [Google Scholar] [CrossRef] [PubMed]
- van Boven, N.; Akkerhuis, K.M.; Anroedh, S.S.; Rizopoulos, D.; Pinto, Y.; Battes, L.C.; Hillege, H.L.; Caliskan, K.C.; Germans, T.; Manintveld, O.C.; et al. Serially measured circulating miR-22-3p is a biomarker for adverse clinical outcome in patients with chronic heart failure: The Bio-SHiFT study. Int. J. Cardiol. 2017, 235, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.P.; Chen, J.; Seok, H.Y.; Zhang, Z.; Kataoka, M.; Hu, X.; Wang, D.Z. MicroRNA-22 regulates cardiac hypertrophy and remodeling in response to stress. Circ. Res. 2013, 112, 1234–1243. [Google Scholar] [CrossRef]
- Patane, S. Value of Serial ST2 Measurements in Acute Heart Failure: miRNA Regulation and Genetic Factors. J. Am. Coll.Cardiol. 2018, 71, 1397. [Google Scholar] [CrossRef] [PubMed]
- Neal, C.L.; Xu, J.; Li, P.; Mori, S.; Yang, J.; Neal, N.N.; Zhou, X.; Wyszomierski, S.L.; Yu, D. Overexpression of 14-3-3zeta in cancer cells activates PI3K via binding the p85 regulatory subunit. Oncogene 2012, 31, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Durrant, T.N.; Hers, I. PI3K inhibitors in thrombosis and cardiovascular disease. Clin. Transl. Med. 2020, 9, 8. [Google Scholar] [CrossRef]
- Zhan, H.; Huang, F.; Niu, Q.; Jiao, M.; Han, X.; Zhang, K.; Ma, W.; Mi, S.; Guo, S.; Zhao, Z. Downregulation of miR-128 Ameliorates Ang II-Induced Cardiac Remodeling via SIRT1/PIK3R1 Multiple Targets. Oxid. Med. Cell. Longev. 2021, 2021, 8889195. [Google Scholar] [CrossRef]
- Simon, L.M.; Edelstein, L.C.; Nagalla, S.; Woodley, A.B.; Chen, E.S.; Kong, X.; Ma, L.; Fortina, P.; Kunapuli, S.; Holinstat, M.; et al. Human platelet microRNA-mRNA networks associated with age and gender revealed by integrated plateletomics. Blood 2014, 123, e37–e45. [Google Scholar] [CrossRef]
- Soutschek, M.; Germade, T.; Germain, P.L.; Schratt, G. enrichMiR predicts functionally relevant microRNAs based on target collections. Nucleic Acids Res. 2022, 50, W280–W289. [Google Scholar] [CrossRef]
- Fazmin, I.T.; Achercouk, Z.; Edling, C.E.; Said, A.; Jeevaratnam, K. Circulating microRNA as a Biomarker for Coronary Artery Disease. Biomolecules 2020, 10, 1354. [Google Scholar] [CrossRef]
- Zhou, S.S.; Jin, J.P.; Wang, J.Q.; Zhang, Z.G.; Freedman, J.H.; Zheng, Y.; Cai, L. miRNAS in cardiovascular diseases: Potential biomarkers, therapeutic targets and challenges. Acta Pharmacol. Sin. 2018, 39, 1073–1084. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Values at Baseline |
|---|---|
| Age (y) | 64.8 ± 9.4 |
| Male (n, %) | 59 (85.5%) |
| BMI (kg/m2) | 26.9 ± 4.4 |
| Heart rate (bpm) | 67.6 ± 11.1 |
| Hemoglobin (g/dl) | 14.3 ± 1.6 |
| Risk factors | |
| Hypertension (n, %) | 39 (56.5%) |
| Stroke (n, %) | 4 (5.8%) |
| Smoker (n, %) | 9 (13%) |
| Diabetes (n, %) | 12 (17.4%) |
| COPD (n, %) | 7 (10.1%) |
| Atrial fibrillation (n, %) | 18 (26.1%) |
| Therapy | |
| ACE-I (n, %) | 52 (75.4%) |
| ARBs (n, %) | 15 (21.7%) |
| Beta blockers (n, %) | 68 (98.6 %) |
| MRA (n, %) | 48 (69.6%) |
| Diuretic (n, %) | 55 (79.7%) |
| Ivabradine (n, %) | 9 (13%) |
| Controls | HF T0 | HF T1 | |
|---|---|---|---|
| miRNA | |||
| miR-29b-3p | 0.895 [0.745–1.4] | 1.530 [1.030–2.090] * | 1.470 [0.92–1.87] |
| miR-28-3p | 0.645 [0.258–1.118] | 0.985 [0.458–1.758] | 1.150 [0.552–2.198] |
| miR-181a-1 | 0.655 [0.36–0.853] | 0.945 [0.550–2.158] | 1.197 [0.60–1.87] |
| miR-221-3p | 1.360 [0.664–2.320] | 1.045 [0.647–1.744] | 1.169 [0.650–1.816] |
| miR-320e | 1.118 [0.8–1.711] | 0.889 [0.541–1.640] | 0.89 [0.567–1.796] |
| miR-423-5p | 0.860 [0.533–1.168] | 0.875 [0.538–1.625] | 1.062 [0.635–1.866] |
| miR-450a-5p | 0.775 [0.408–1.320] | 1.017 [0.545–1.863] | 1.082 [0.575–1.797] |
| miR-503-5p | 1.035 [0.703–1.525] | 1.073 [0.524–2.357] | 1.132 [0.492–2.292] |
| Age (years) | EF (%) | Peak VO2 (mL/min) | Peak VO2 (mL/min/kg) | DLCO (mL/min/mmHg) | ST-2 (ng/mL) | NT-proBNP (pg/mL) | ||
|---|---|---|---|---|---|---|---|---|
| miRNA T0 | ||||||||
| miR-29b-3p | R | 0.411 ** | −0.132 | −0.405 ** | −0.371 ** | −0.089 | −0.067 | 0.119 |
| p values | 0.001 | 0.279 | 0.001 | 0.003 | 0.477 | 0.673 | 0.332 | |
| miR-28-3p | R | 0.277 * | 0.043 | −0.142 | −0.178 | −0.055 | −0.086 | −0.150 |
| p values | 0.021 | 0.725 | 0.258 | 0.160 | 0.663 | 0.587 | 0.220 | |
| mir-181a-1 | R | 0.189 | −0.012 | −0.132 | −0.170 | −0.020 | −0.125 | −0.141 |
| p values | 0.119 | 0.925 | 0.294 | 0.180 | 0.871 | 0.432 | 0.249 | |
| miR-221-3p | R | 0.340 ** | −0.026 | −0.261 * | −0.271 * | −0.014 | −0.052 | −0.064 |
| p values | 0.004 | 0.832 | 0.036 | 0.030 | 0.912 | 0.745 | 0.599 | |
| miR-320e | R | 0.027 | 0.015 | −0.068 | −0.155 | −0.060 | −0.036 | −0.138 |
| p values | 0.823 | 0.906 | 0.592 | 0.221 | 0.632 | 0.819 | 0.260 | |
| miR-423-5p | R | 0.155 | −0.016 | −0.123 | −0.172 | 0.005 | −0.116 | −0.220 |
| p values | 0.204 | 0.895 | 0.327 | 0.173 | 0.970 | 0.466 | 0.069 | |
| miR-450a-5p | R | −0.036 | −0.083 | −0.097 | 0.063 | 0.030 | 0.078 | 0.108 |
| p values | 0.771 | 0.499 | 0.446 | 0.621 | 0.813 | 0.623 | 0.379 | |
| miR-503-5p | R | 0.148 | −0.061 | −0.304 * | −0.209 | −0.148 | 0.286 | 0.165 |
| p values | 0.238 | 0.632 | 0.017 | 0.110 | 0.250 | 0.074 | 0.188 |
| EF (%) | Peak VO2 (mL/min) | Peak VO2 (mL/min/kg) | DLCO (mL/min/ mmHg) | ST-2 (ng/mL) | NT-proBNP (pg/mL) | ||
|---|---|---|---|---|---|---|---|
| miRNA follow-up | |||||||
| miR-29b-3p | R | 0.003 | −0.134 | 0.030 | −0.166 | 0.357 ** | 0.141 |
| p values | 0.982 | 0.291 | 0.816 | 0.196 | 0.009 | 0.252 | |
| miR-28-3p | R | 0.184 | −0.092 | 0.062 | −0.094 | 0.189 | 0.000 |
| p values | 0.132 | 0.469 | 0.625 | 0.465 | 0.176 | 0.998 | |
| mir-181a-1 | R | 0.049 | −0.032 | 0.111 | 0.016 | 0.212 | −0.029 |
| p values | 0.690 | 0.802 | 0.381 | 0.904 | 0.127 | 0.815 | |
| miR-221-3p | R | 0.051 | −0.212 | −0.045 | −0.097 | 0.329 * | 0.093 |
| p values | 0.680 | 0.092 | 0.727 | 0.452 | 0.016 | 0.449 | |
| miR-320e | R | −0.160 | −0.133 | −0.072 | −0.010 | 0.416 ** | 0.263 * |
| p values | 0.191 | 0.295 | 0.572 | 0.936 | 0.002 | 0.031 | |
| miR-423-5p | R | 0.010 | −0.052 | −0.005 | 0.081 | 0.285 * | 0.033 |
| p values | 0.933 | 0.683 | 0.966 | 0.531 | 0.039 | 0.786 | |
| miR-450a-5p | R | −0.117 | −0.011 | 0.154 | 0.067 | 0.419 ** | 0.058 |
| p values | 0.343 | 0.932 | 0.225 | 0.608 | 0.002 | 0.641 | |
| miR-503-5p | R | 0.189 | −0.129 | −0.006 | −0.070 | 0.235 | 0.051 |
| p values | 0.135 | 0.328 | 0.961 | 0.603 | 0.100 | 0.690 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brioschi, M.; D’Alessandra, Y.; Mapelli, M.; Mattavelli, I.; Salvioni, E.; Eligini, S.; Mallia, A.; Ricci, V.; Gianazza, E.; Ghilardi, S.; et al. Impact of Sacubitril/Valsartan on Circulating microRNA in Patients with Heart Failure. Biomedicines 2023, 11, 1037. https://doi.org/10.3390/biomedicines11041037
Brioschi M, D’Alessandra Y, Mapelli M, Mattavelli I, Salvioni E, Eligini S, Mallia A, Ricci V, Gianazza E, Ghilardi S, et al. Impact of Sacubitril/Valsartan on Circulating microRNA in Patients with Heart Failure. Biomedicines. 2023; 11(4):1037. https://doi.org/10.3390/biomedicines11041037
Chicago/Turabian StyleBrioschi, Maura, Yuri D’Alessandra, Massimo Mapelli, Irene Mattavelli, Elisabetta Salvioni, Sonia Eligini, Alice Mallia, Veronica Ricci, Erica Gianazza, Stefania Ghilardi, and et al. 2023. "Impact of Sacubitril/Valsartan on Circulating microRNA in Patients with Heart Failure" Biomedicines 11, no. 4: 1037. https://doi.org/10.3390/biomedicines11041037