4.2. mtDNA Polymorphism and Its Relation to Diseases and Aging
Single-nucleotide polymorphism studies allowed to determine genotypes responsible for a specific disease in monogenic diseases, and recently also genotypes with a high risk of multigene diseases [
32,
33]. One of the possible consequences of an mtDNA mutation—monogenic diseases—is rare. For example, various mtDNA mutations may be responsible for the monogenic mitochondrial disease—Leber’s hereditary optic neuropathy (LHON)—and 90% of patients have one of them: 11778G/A; 3460G/A; and 14484T/C. These mutations in the population are found with a frequency of 1/300 [
34]. Changes in the nucleotide sequence in mtDNA are easy, easier than in the nuclear genome, which is associated with exposure of the mitochondrial genome to contact with continuously produced reactive oxygen species, as well as reduced mtDNA repair possibilities [
35,
36]. More frequent occurrence of mtDNA mutations in older age groups of patients was observed by Michikawa et al. [
37]. Changes in the mtDNA nucleotide sequence occurred in the majority (in 57% of the studied patients) of the studied patients over 65 years of age, which, however, were not observed in the groups of younger patients. With age, the progressive increase in mtDNA mutations as a result of reaction to reactive oxygen species is secondary to lipid oxidation and the modification of mitochondrial proteins in the cell’s respiratory chain [
38,
39]. MtDNA mutations change the structure of the respiratory chain polypeptides encoded in the mitochondrial genome, reducing mitochondrial metabolic activity and the formation of high-energy compounds [
40]. Respiratory chain enzymes encoded by altered mtDNAs disrupt electron transport, increase electron leakage from the respiratory chain, and increase the amount of free oxygen radicals produced, further damaging the mitochondria and creating a vicious circle effect. This mechanism leads to the deterioration of the functioning of organs and tissues during the aging process. Modification of apoptosis signaling secondary to mitochondrial damage has been observed in in vivo and in vitro studies. This thesis is confirmed by the results of studies by Wei et al. on skin fibroblasts [
41]. The above-mentioned researchers observed greater disturbances in fibroblast bioenergetics in the elderly compared to younger people, which was assessed on the basis of a higher concentration of hydrogen peroxide, a high level of superoxide dismutase activity, and a decrease in the activity of cytochrome c oxidase, as well as the oxygen consumption rate in the older age group. At the same time, a decrease in pyruvate dehydrogenase (PDH) expression and an increase in lactate dehydrogenase kinase were observed.
4.3. Study Subjects and Comparison of Results with Previous Studies
Despite the relationships of mtDNA polymorphism with age repeatedly described in the scientific literature, our studies did not show significant differences in age between the groups of women studied with changes in nucleotide sequences in the D-loop and without mtDNA changes. MtDNA changes with age, hence, we studied women over 40 (between 40–60 years old). The restricted age range of the research group may have prevented age disparities between women with mtDNA nucleotide sequence alterations and those without. Disturbances in the physiological functions of mitochondria may depend not only on the direct effect of the mutation on the respiratory chain, but may also occur secondary to the existing multigene disease and dysfunction of the mitochondrial respiratory chain in its course. However, it should be emphasized that certain mtDNA mutations may be beneficial. There are publications regarding changes in nucleotide sequences in mtDNA accompanying longevity. Studies by Kokaze et al. found that the mtDNA 5178 C/A polymorphism, which is associated with longevity, may prevent the onset of diabetes [
42]. It has been shown that this genotype reduces the number of mtDNA mutations in oocytes, as well as the rate of mtDNA mutation formation and their accumulation in somatic cells in Japanese centenarians. The mtDNA 5178 C/A polymorphism not only prevents diabetes but is probably responsible for inhibiting the development of myocardial infarction [
43]. Zhang et al., in the Italian population, studied the frequency of the C150T mutation located near the sequences responsible for mtDNA heavy strand synthesis. Its occurrence was more frequent in older age groups [
44]. It appeared in approximately 17% of people (33/52) aged 99–106, while in younger people (aged 18–98) only in 3.4% (3/117).
Howell et al. and others found that mutations associated with multigenetic illnesses commonly occur in the D-loop region, a 1122 bp non-coding stretch of mtDNA containing two hypervariable regions, HV1 and HV2 [
45,
46]. This area has higher mtDNA polymorphism than others. Del Bo et al. found more mutations in the HV1 and HV2 hypervariable regions of the D-loop than other mtDNA segments in aged people [
47]. Mutations in D-loop mtDNA nucleotide sequences, which occur often, disrupt mitochondrial genome replication and transcription. A single-nucleotide polymorphism in the D-loop and 12S RNA coding sequence of mtDNA was detected in 23% of our respondents. Like the aforementioned authors, we found more frequent changes in the hypervariable segments HV1 and HV2 of the D-loop, which occurred in 21.0% of respondents, somewhat more often in the HV1 segment (14.0% of respondents) than in HV2. In the HV1 area, 85.7% of mutations were homoplasmic and non-coding, and just one patient implicated nucleotide 16319, the beginning site for mtDNA synthesis and light strand engaged in replication. Nucleotide sequence alterations in the HV2 region were homoplasmic in 90.1% of instances, connected to the transcription factor binding site in CBS3 (conserved block) in 36%, and occurred in nearly 30% of responses. The 12S rDNA region has 10% non-coding nucleotides, 90% 1438AG and 750AG, and 20% 930GA. Rydzanicz et al. found polymorphisms in the 12S rDNA region (G709A, G750A, G930A, T1243C, T1420C, and G1438A) at a frequency greater than 1% [
48]. This study found two mtDNA polymorphisms in the HV2 hypervariable section of the mitochondrial genome that have not been previously reported. A postmenopausal woman with arterial hypertension and metabolic syndrome had a polymorphism. The non-coding nucleotide 340C/A in the H strand origin region between the DNA replication primer and the CBS3 block was changed. The second nucleotide sequence change included the non-coding nucleotide 362T/C and the conserved CBS3 block. Our postmenopausal control patient showed another polymorphism. It was in the mtDNA 12S RNA coding sequence and associated with the non-coding nucleotide 812A/C. Most of the other alterations in the investigated population include mutations associated with multigene illnesses, coronary artery disease, hypertension, diabetes, and neoplastic diseases, according to the literature [
26,
49,
50]. In multigene diseases, the disease process in such cases is not caused by a single change in nucleotide sequences in mtDNA, but by changes in many genes. The presence of mtDNA polymorphisms is not a prerequisite for clinical symptoms of the disease, and changes in the mitochondrial genome occur only in some patients with clinical symptoms of the disease. Homoplastic mutations, which do not always translate into the phenotype of clinical disease symptoms, are often diagnosed at random. This fact is explained by many authors by the direct influence on the mitochondrial genome of the nuclear genome and the influence of epigenetic factors, while the disclosure of heterozygous mutations is conditioned by the proportion of mutated and normal mtDNA [
51,
52]. Most polymorphisms occur as a variant of the genotype not associated with the occurrence of a given disease, and changes in nucleotide sequences are often located only in the vicinity of genes responsible for a given disease entity. As a result of changes in mtDNA, the same mutation may cause various sets of clinical symptoms or be asymptomatic. In summary, mitochondrial mutations may cause phenotypic effects that are difficult to predict and may occur as pathogenic or only potentially pathogenic mutations.
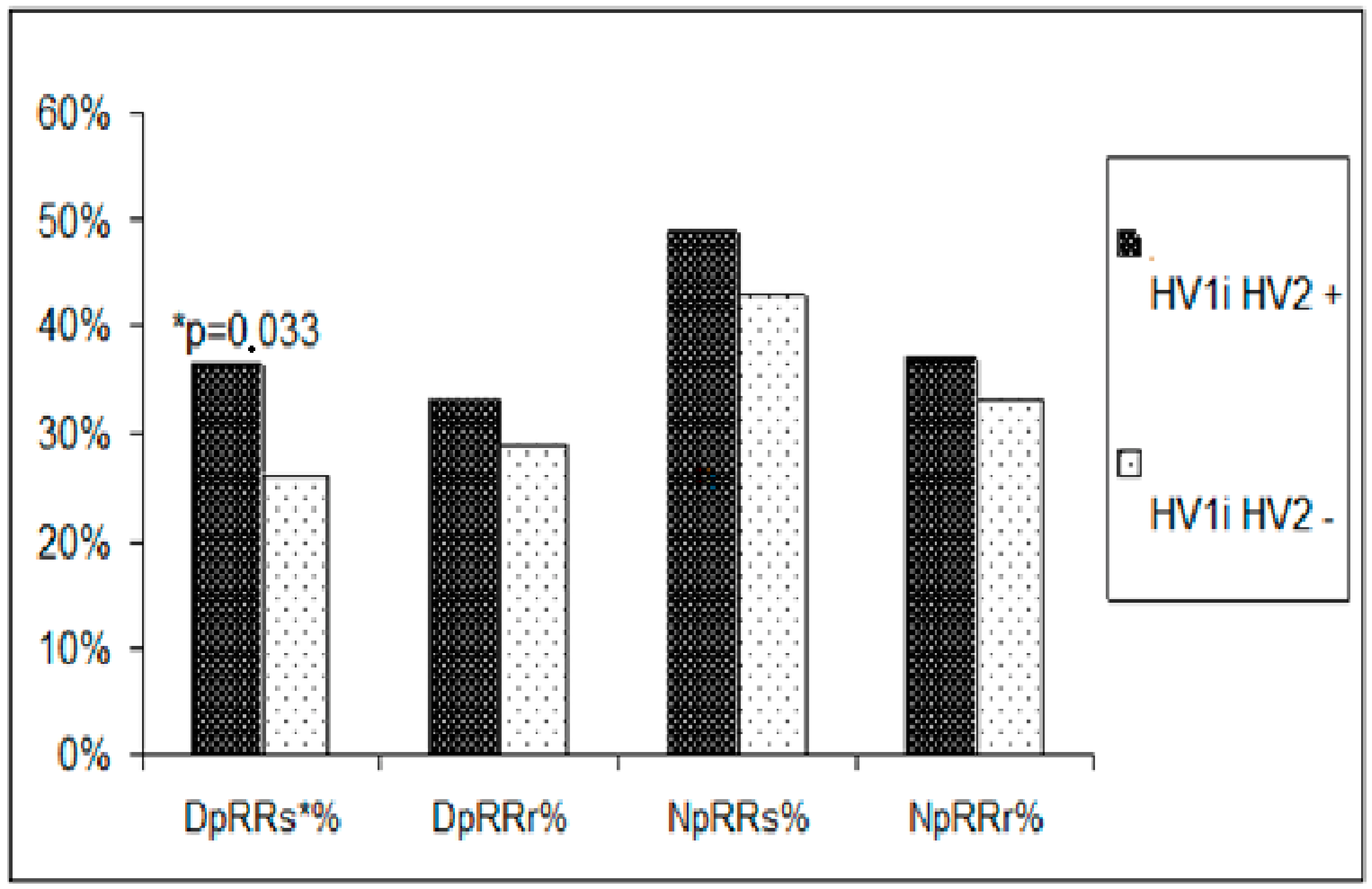
The study found that alterations in mtDNA in HV1, HV2, and 12S rDNA may impact arterial hypertension by increasing blood pressure day and night and heart rate, suggesting an increased adrenergic system tone in these people. The HV2 segment (239TC, 243AG, 247GA, 250TC, 260GA, 277CT, and 284CT) alterations mostly affected transcription factor binding sites and non-coding nucleotides in the 12S RNA and HV1 coding regions. According to Pejovic et al., the nucleotide sequences in the hypervariable D-loop regions that replicate mtDNA and the degree of transcription factor binding can alter mtDNA synthesis and cell number [
53]. The examined women’s HV2 region alterations, which impact mtDNA synthesis and transcription factor binding, may affect blood pressure. Several studies link hypertension to mitochondrial metabolism and free oxygen radicals [
49,
54]. On the other hand, the authors of experimental studies describe various forms of damage to the mitochondrial respiratory chain, leading to an increase in the production of reactive oxygen species that affect the course of hypertension. The source of reactive oxygen species in blood vessels are vascular endothelial cells, fibroblasts, and vascular smooth muscle, in which NAD (P) H or NADH oxidase (nicotinamide adenine dinucleotide in reduced form) catalyzing the reduction of oxygen causes the formation of O2 - and large amounts of other free radical oxygen. NAD (P) H oxidase activation occurs under the influence of TNF-α, angiotensin, and nitric oxide synthase. Hydrogen peroxide is a vasoactive compound with vasoconstrictor properties. According to Rubanyi et al., peroxygen hydrogen chloride in reaction with nitric oxide can transform into peroxynitrite anion (ONOO–), which reduces the availability of nitric oxide and thus contributes to the development of hypertension [
55]. According to Pryor et al., hydrogen peroxide directly affects the opening of potassium and calcium channels, and therefore is also responsible for vasodilation. In turn, nitric oxide synthase is a source of not only NO, but also O
2, which reduces the availability of NO [
56]. According to the authors cited above, the balance between NO and O
2, is essential for the damage to the vessel wall, the state of vascular tone, and the development of arterial hypertension. The production of large amounts of free oxygen radicals activates the tyrosine phosphatase and tyrosine kinase pathways, influences the expression of transcription factors and mitogen-activated protein kinases, and changes the activity of ion channels. ROS directly increases the concentration of calcium ions in the cell, leading to vessel wall dysfunction and remodeling. The changes in mtDNA observed in the group of women we have studied, accompanying higher blood pressure values, are probably the result of damage to the mitochondria and the formation of ROS. Many of the available publications on mitochondrial mutations in studied patients with maternal hypertension refer to Asian populations [
57,
58,
59,
60,
61]. Various degrees of arterial hypertension recognized in the presence of mutations were observed: mutation 4435A > G, with a 30% reduction in mitochondrial metabolism and mitochondrial tRNA transcription (Met); mutation 4263A > G, located at the site of transcription for isoleucine (5’ end of tRNA (Ile)), which decreased the efficiency of the tRNA replication process by about 46%; the 4401A > G nucleotide mutation located directly at the 5’ end of the tRNA (Met) and tRNA (Gln), with a reduction in the mitochondrial translation index and a reduction in the mitochondrial respiratory efficiency index; and mutation T3308C on the dehydrogenase subunit (ND1), in which the translation initiating amino acid-methionine has been replaced with tyronine in ND1, with the alteration of RNA precursor strand processing or destabilization of ND1-mtRNA dehydrogenase.
The results of this study and other investigations suggest that alterations in the mtDNA nucleotide sequence may cause arterial hypertension. A shortage of postmenopausal sex hormones may damage the mitochondrial respiratory chain, causing mtDNA alterations and hypertension as a multigene illness. The literature shows multigene alterations in mtDNA coding and non-coding nucleotides in arterial hypertension, comparable to our findings [
62]. The function of mtDNA D-loop hypervariable regions in arterial hypertension was examined by Liu et al. Changes in the HV2 area 152T-> C, 182C-> T, and 247G-> A and HV1 segment 16187C-> T, 16189T-> C, 16264C-> T, 16270C-> T, and 16311T-> C predispose to essential hypertension. According to these authors, the development of hypertension does not correlate to the severity of the mutation’s influence on the illness’s clinical picture, and the environment and nuclear gene modifiers in these individuals also modify the mutation’s effect on the disease. In a Japanese population investigation on the association between mtDNA alterations and arterial hypertension, Soji et al. found that the mtDNA genotype 16223TC is more prevalent in hypertensive patients than in those without hypertension and is associated with higher hypertension risk [
63]. They found no connection with other genotypes, including C16362T. As in this study, studies on the mitochondrial genome generally focused on single or many gene coding or non-coding agents and their link with disease entities, such as hypertension.
4.4. Strength and Limitations of the Study
One of the limitations of our research is that it involved women from 40 to 60 years of age, who could be expected to have changes in mtDNA with age. Another issue perhaps would be the small age range of the study group, which meant that the differences in the age of the studied women with changes in mtDNA nucleotide sequences compared to the patients without changes did not occur.
The investigators did not investigate other factors leading to hypertension, such as nutrition, which is a drawback of the study. Obesity can induce hypertension and therefore have an impact on changes in the mitochondrial genome sequence.
Our discovery may have clinical relevance in that Hormone Replacement Therapy may have a protective effect on mtDNA mutation. Several experimental medicines have already reached the clinical phase with extremely promising findings, yet the likelihood of enrolling patients in clinical trials is limited.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}