
Development of Cellular Signaling Pathways by Bioceramic Heat Treatment (Sintering) in Osteoblast Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Sample Preparation
2.3. X-ray Diffraction (XRD)
2.4. Field-Emission Scanning Electron Microscopy (FE-SEM)
2.5. X-ray Photoelectron Spectroscopy (XPS)
2.6. Preparation of Bioceramic Plates
2.7. Western Blotting
2.8. Inhibitors
2.9. Intracellular Calcium Measurements
2.10. Statistical Analysis
3. Results
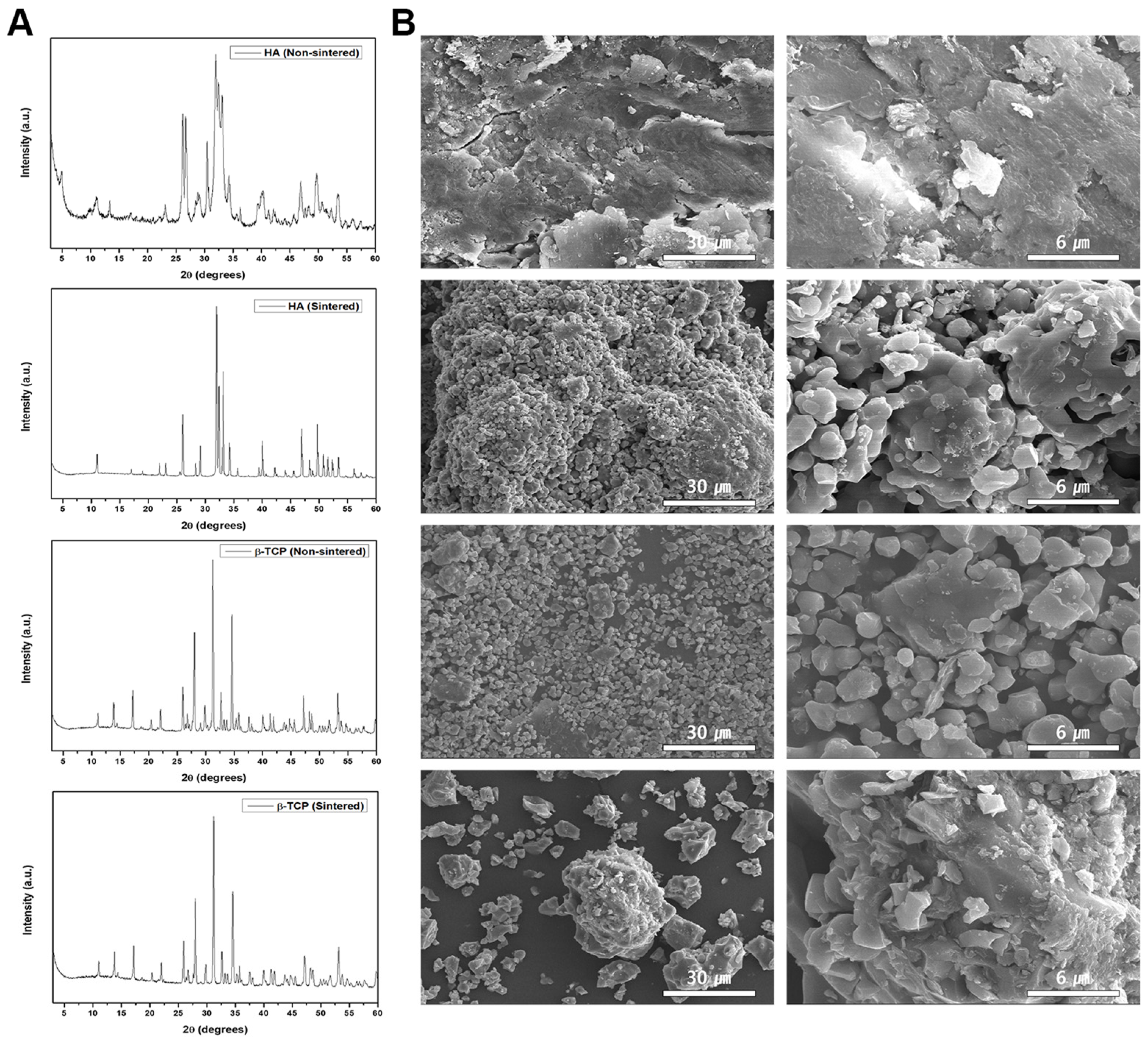
3.1. Phase Characteristics of the Sintered and Non-Sintered HA β-TCP Crystals
3.2. Effect of the Bioceramics on the Intracellular Calcium Concentration
3.3. Effect of the Bioceramics on the Regulated JNK Pathway
3.4. Effect of the Bioceramics on the p-Src Pathway
3.5. Effect of the Bioceramics on the EGFR Signaling Pathway
3.6. Effect of the Bioceramics on the Raptor/MTORC1 Signaling Pathway
3.7. Effect of the Bioceramics on the p38 Signaling Pathway
3.8. Effect of the Bioceramics on AKT Phosphorylation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoshikawa, H.; Myoui, A. Bone tissue engineering with porous hydroxyapatite ceramics. J. Artif. Organs. 2005, 8, 131–136. [Google Scholar] [CrossRef]
- Betz, R.R. Limitations of autograft and allograft: New synthetic solutions. Orthopedics 2002, 25, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Khan, Y.; Yaszemski, M.J.; Mikos, A.G.; Laurencin, C.T. Tissue engineering of bone: Material and matrix considerations. J. Bone Jt. Surg. Am. 2008, 90, 36–42. [Google Scholar] [CrossRef]
- Müller, P.; Bulnheim, U.; Diener, A.; Lüthen, F.; Teller, M.; Klinkenberg, E.-D.; Neumann, H.-G.; Nebe, B.; Liebold, A.; Steinhoff, G. Calcium phosphate surfaces promote osteogenic differentiation of mesenchymal stem cells. J. Cell. Mol. Med. 2008, 12, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Song, T.; Chen, X.; Wang, M.; Yang, X.; Xiao, Y.; Zhang, X. Osteoinductivity of porous biphasic calcium phosphate ceramic spheres with nanocrystalline and their efficacy in guiding bone regeneration. ACS Appl. Mater. Interfaces 2019, 11, 3722–3736. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Kim, J.H.; Shim, J.H.; Hwang, N.S.; Heo, C.Y. Bioactive calcium phosphate materials and applications in bone regeneration. Biomater. Res. 2019, 23, 4. [Google Scholar] [CrossRef] [Green Version]
- Kitsugi, T.; Yamamuro, T.; Takeuchi, H.; Ono, M. Bonding behavior of three types of hydroxyapatite with different sintering temperatures implanted in bone. Clin. Orthop. Relat. Res. 1988, 234, 280–290. [Google Scholar] [CrossRef]
- Raynaud, S.; Champion, E.; Bernache-Assollant, D. Calcium phosphate apatites with variable Ca/P atomic ratio II. Calcination and sintering. Biomaterials 2002, 23, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-J.L. Sintering Processes. In Sintering—Densification, Grain Growth, and Microstructure, 1st ed.; Butterworth-Heinemann: Oxford, UK, 2005; pp. 3–8. [Google Scholar]
- Grasso, S.; Biesuz, M.; Zoli, L.; Taveri, G.; Duff, A.I.; Ke, D.; Reece, M.J. A review of cold sintering processes. Adv. Appl. Ceram. 2020, 119, 115–143. [Google Scholar] [CrossRef] [Green Version]
- Sofronia, A.M.; Baies, R.; Anghel, E.M.; Marinescu, C.A.; Tanasescu, S. Thermal and structural characterization of synthetic and natural nanocrystalline hydroxyapatite. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 43, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Indurkar, A.; Choudhary, R.; Rubenis, K.; Locs, J. Advances in sintering techniques for calcium phosphates ceramics. Materials 2021, 14, 6133. [Google Scholar] [CrossRef]
- Saha, N.; Keskinbora, K.; Suvaci, E.; Basu, B. Sintering, microstructure, mechanical, and antimicrobial properties of HAp-ZnO biocomposites. J. Biomed. Mater. Res. B Appl. Biomater. 2010, 95, 430–440. [Google Scholar] [CrossRef]
- Champion, E. Sintering of calcium phosphate bioceramics. Acta Biomater. 2013, 9, 5855–5875. [Google Scholar] [CrossRef] [PubMed]
- Biesuz, M.; Galotta, A.; Motta, A.; Kermani, M.; Grasso, S.; Vontorová, J.; Tyrpekl, V.; Vilémová, M.; Sglavo, V.M. Speedy bioceramics: Rapid densification of tricalcium phosphate by ultrafast high-temperature sintering. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 127, 112246. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Y.; Uskokovic, P.S.; Tsui, C.P.; Veljovic, D.; Petrovic, R.; Janackovic, D. Influence of microstructure and phase composition on the nanoindentation characterization of bioceramic materials based on hydroxyapatite. Ceram. Int. 2009, 35, 2171–2178. [Google Scholar] [CrossRef]
- Ruys, A.J.; Wei, M.; Sorrell, C.C.; Dickson, M.R.; Brandwood, A.; Milthorpe, B.K. Sintering effects on the strength of hydroxyapatite. Biomaterials 1995, 16, 409–415. [Google Scholar] [CrossRef]
- Kitsugi, T.; Yamamuro, T.; Nakamura, T.; Kokubo, T.; Takagi, M.; Shibuya, T.; Takeuchi, H.; Ono, M. Bonding behavior between two bioactive ceramics in vivo. J. Biomed. Mater. Res. 1987, 21, 1109–1123. [Google Scholar] [CrossRef]
- Laquerriere, P.; Kilian, L.; Bouchot, A.; Jallot, E.; Grandjean, A.; Guenounou, M.; Balossier, G.; Frayssinet, P.; Bonhomme, P. Effect of hydroxyapatite sintering temperature on intracellular ionic concentrations of monocytes: A TEM-cryo-x-ray microanalysis study. J. Biomed. Mater. Res. 2001, 58, 238–246. [Google Scholar] [CrossRef]
- Eliaz, N.; Metoki, N. Calcium phosphate bioceramics: A review of their history, structure, properties, coating technologies and biomedical applications. Materials 2017, 10, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurgelane, I.; Buss, A.; Putnina, M.; Loca, D.; Locs, J. Effect of sintering temperature on sorption properties and compressive strength of calcium phosphate ceramic granules. Mater. Lett. 2021, 282, 128858. [Google Scholar] [CrossRef]
- Frasnelli, M.; Sglavo, V.M. Flash sintering of tricalcium phosphate (TCP) bioceramics. J. Eur. Ceram. Soc. 2018, 38, 279–285. [Google Scholar] [CrossRef]
- Ryu, H.-S.; Youn, H.-J.; Hong, K.S.; Chang, B.-S.; Lee, C.-K.; Chung, S.-S. An improvement in sintering property of beta-tricalcium phosphate by addition of calcium pyrophosphate. Biomaterials 2002, 23, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, Y.; Shibata, Y.; Kataoka, Y.; Miyazaki, T.; Nishiyama, N. Osteoblast-like cell proliferation on tape-cast and sintered tricalcium phosphate sheets. Acta Biomater. 2008, 4, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hanagata, N.; Maeda, M.; Minowa, T.; Ikoma, T.; Fan, H.; Zhang, X. Porous hydroxyapatite and biphasic calcium phosphate ceramics promote ectopic osteoblast differentiation from mesenchymal stem cells. Sci. Technol. Adv. Mater. 2009, 10, 025003. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, O.; Shiwaku, Y.; Hamai, R. Octacalcium phosphate bone substitute materials: Comparison between properties of biomaterials and other calcium phosphate materials. Dent. Mater. J. 2020, 39, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Habraken, W.J.E.M.; Tao, J.; Brylka, L.J.; Friedrich, H.; Bertinetti, L.; Schenk, A.S.; Verch, A.; Dmitrovic, V.; Bomans, P.H.H.; Frederik, P.M.; et al. Ion-association complexes unite classical and non-classical theories for the biomimetic nucleation of calcium phosphate. Nat. Commun. 2013, 4, 1507. [Google Scholar] [CrossRef] [Green Version]
- Horváthová, R.; Müller, L.; Helebrant, A.; Greil, P.; Müller, F.A. In vitro transformation of OCP into carbonated HA under physiological conditions. Mater. Sci. Eng. C Mater. Biol. Appl. 2008, 28, 1414–1419. [Google Scholar] [CrossRef]
- Muralithran, G.; Ramesh, S. The effects of sintering temperature on the properties of hydroxyapatite. Ceram. Int. 2000, 26, 221–230. [Google Scholar] [CrossRef]
- Stevie, F.A.; Donley, C.L. Introduction to x-ray photoelectron spectroscopy. J. Vac. Sci. Technol. A 2020, 38, 063204. [Google Scholar] [CrossRef]
- Pagano, S.; Lombardo, G.; Caponi, S.; Costanzi, E.; Michele, A.D.; Bruscoli, S.; Xhimitiku, I.; Coniglio, M.; Valenti, C.; Mattarelli, M.; et al. Bio-mechanical characterization of a CAD/CAM PMMA resin for digital removable prostheses. Dent. Mater. 2021, 37, e118–e130. [Google Scholar] [CrossRef]
- Zhou, T.; Gao, B.; Fan, Y.; Liu, Y.; Feng, S.; Cong, Q.; Zhang, X.; Zhou, Y.; Yadav, P.S.; Lin, J.; et al. Piezo1/2 mediate mechanotransduction essential for bone formation through concerted activation of NFAT-YAP1-ß-catenin. eLife 2020, 9, e52779. [Google Scholar] [CrossRef]
- Zheng, W.; Gu, X.; Hu, D.; Hao, Y. Co-culture with synovial tissue in patients with rheumatoid arthritis suppress cell proliferation by regulating MAPK pathway in osteoblasts. Am. J. Transl. Res. 2019, 11, 3317–3327. [Google Scholar]
- Zhu, W.-Q.; Ming, P.-P.; Zhang, S.-M.; Qiu, J. Role of MAPK/JNK signaling pathway on the regulation of biological behaviors of MC3T3-E1 osteoblasts under titanium ion exposure. Mol. Med. Rep. 2020, 22, 4792–4800. [Google Scholar] [CrossRef]
- Li, X.; Lin, H.; Zhang, X.; Jaspers, R.T.; Yu, Q.; Ji, Y.; Wu, G. Notoginsenoside R1 attenuates oxidative stress-induced osteoblast dysfunction through JNK signaling pathway. J. Cell. Mol. Med. 2021, 25, 11278–11289. [Google Scholar] [CrossRef] [PubMed]
- Yaginuma, T.; Gao, J.; Nagata, K.; Muroya, R.; Fei, H.; Nagano, H.; Chishaki, S.; Matsubara, T.; Kokabu, S.; Matsuo, K.; et al. p130Cas induces bone invasion by oral squamous cell carcinoma by regulating tumor epithelial–mesenchymal transition and cell proliferation. Carcinogenesis 2020, 41, 1038–1048. [Google Scholar] [CrossRef]
- Thouverey, C.; Ferrari, S.; Caverzasio, J. Selective inhibition of Src family kinases by SU6656 increases bone mass by uncoupling bone formation from resorption in mice. Bone 2018, 113, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Parsons, S.J.; Parsons, J.T. Src family kinases, key regulators of signal transduction. Oncogene 2004, 23, 7906–7909. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, T.; Yasuda, K.; Mizuta, K.; Kawaue, H.; Kokabu, S. Tyrosine Kinase Src Is a Regulatory Factor of Bone Homeostasis. Int. J. Mol. Sci. 2022, 23, 5508. [Google Scholar] [CrossRef] [PubMed]
- Alvandi, Z.; Opas, M. c-Src kinase inhibits osteogenic differentiation via enhancing STAT1 stability. PLoS ONE 2020, 15, e0241646. [Google Scholar] [CrossRef]
- Raymond, E.; Faivre, S.; Armand, J.P. Epidermal growth factor receptor tyrosine kinase as a target for anticancer therapy. Drugs 2000, 60, 15–23. [Google Scholar] [CrossRef]
- Fan, J.B.; Liu, W.; Yuan, K.; Zhu, X.H.; Xu, D.W.; Chen, J.J.; Cui, Z.M. EGFR trans-activation mediates pleiotrophin-induced activation of Akt and Erk in cultured osteoblasts. Biochem. Biophys. Res. Commun. 2014, 447, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. p38 MAPK signaling in osteoblast differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasini, M.; Cagidiaco, I.; Fambrini, E.; Miceli, M.; Carli, E. Life quality of children affected by cleft lip palate and alveolus (CLPA). Children 2022, 9, 757. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, Y.; Kim, J.; Kim, S.; Chung, S.h.; Wie, J. Development of Cellular Signaling Pathways by Bioceramic Heat Treatment (Sintering) in Osteoblast Cells. Biomedicines 2023, 11, 785. https://doi.org/10.3390/biomedicines11030785
Jung Y, Kim J, Kim S, Chung Sh, Wie J. Development of Cellular Signaling Pathways by Bioceramic Heat Treatment (Sintering) in Osteoblast Cells. Biomedicines. 2023; 11(3):785. https://doi.org/10.3390/biomedicines11030785
Chicago/Turabian StyleJung, Yoona, Jooseong Kim, Sukyoung Kim, Shin hye Chung, and Jinhong Wie. 2023. "Development of Cellular Signaling Pathways by Bioceramic Heat Treatment (Sintering) in Osteoblast Cells" Biomedicines 11, no. 3: 785. https://doi.org/10.3390/biomedicines11030785